
两种血管紧张素转化酶抑制肽作用于靶标的分子机理
2010-03-23 05:36郭慧青潘道东
食品科学 2010年23期
郭慧青,毛 慧,赵 波,潘道东,2,*
(1.南京师范大学 国家乳品加工技术研发分中心,江苏 南京 210097;2.宁波大学生命科学与生物工程学院,浙江 宁波 315211)
两种血管紧张素转化酶抑制肽作用于靶标的分子机理
郭慧青1,毛 慧1,赵 波1,潘道东1,2,*
(1.南京师范大学 国家乳品加工技术研发分中心,江苏 南京 210097;2.宁波大学生命科学与生物工程学院,浙江 宁波 315211)
采用脱脂乳为原料,利用四步反相高压液相色谱从Lactobacillus helveticus和Lactobacillus casei subsp. casei制作的酸乳中分离到两种血管紧张素转化酶Ⅰ(angiotensin Ⅰ-converting enzyme,ACE)抑制肽VPP和IPP,测得它们抑制ACE活性的IC50分别为8.89μmol/L和5.17μmol/L。根据氨基酸序列分析的结果,构建VPP和IPP的分子结构,通过分子柔性对接方法研究它们与ACE相互作用的分子机理,确定它们的作用位点、作用力类型及相互作用能。结果表明:VPP、IPP和ACE的活性口袋之间均形成3个氢键,且存在疏水,亲水等作用力,IPP与ACE之间的结合较VPP更稳定,与IPP比VPP具有较低的IC50值的实验结果相一致。
VPP;IPP;ACE抑制肽;分子模拟
血管紧张素转化酶Ⅰ(angiotensinⅠ- converting enzyme,ACE,EC 3.4.15.1)是一种哺乳动物组织中普遍存在的Zn2+依赖型梭二肽酶,为膜整合的单链糖蛋白。它能将无升压活性的血管紧张素Ⅰ(Asp-Arg-Va-Tyr-Ile-His-Pro-Phe-His-Leu)末端的二肽(His-Leu)切割下来,使之转变为有较强血管收缩调节活性的血管紧张素Ⅱ(Asp-Arg-Val-Tyr-Ile-His-Pro-Phe);同时,该酶还能使缓激肽失活,转变为没有活力的缓释肽,从而使血管平活肌收缩,导致血压升高[1-6]。ACE抑制肽是经蛋白质分解酶的作用而产生的一类具有抑制ACE活性的多肽物质,
它通过抑制ACE的活性,阻止血管紧张素Ⅰ转化为血管紧张素Ⅱ反应的发生以及缓激肽的裂解,从而起到降低高血压的作用。
乳蛋白是一种全价蛋白质,除了作为人类主要的膳食蛋白外,还是多种生物活性物质的主要来源。近年来,国内外的研究表明,乳源性ACE抑制肽具有安全性高、无副作用等优点[4]。对乳源性ACE抑制肽的研究也因此成为了一个新的热点,但研究多集中在ACE抑制肽的生产方法和分离途径上,对ACE抑制肽的构效关系和抑制机理鲜有报道。
本研究从具有较强蛋白分解酶系统的Lactobacillus helveticus JCM 1004和 Lactobacillus casei subsp. casei ATCC393所制作的酸乳中分离出具有较强抗高血压作用的活性肽VPP和IPP,进行氨基酸序列分析并测试它们的抑制ACE活性的IC50;根据氨基酸序列分析的结果,采用分子模拟的方法,构建VPP和IPP的分子结构,研究其与ACE作用的分子机理,这方面的研究对于了解其关键结构,设计活性更高的抑制剂都具有重要的意义。
1 材料与方法
1.1 材料与试剂
脱脂乳粉 市售;Lactobacillus helveticus JCM 1004、Lactobacillus casei subsp. casei ATCC393实验室筛选保藏菌株;hippuryl-l-histidyl-l-leucine (HHL)、angiotensin-Ⅰ converting enzyme (ACE、) 4-(2-hydroxyerhyl) piperazine-1-erhanesulfonic acid (HEPES) Sigma公司;其他试剂购自于南京生兴生物技术有限公司。
1.2 仪器与设备
超净工作台 上海上净净化食品有限公司;PHS-3C精密pH 计 上海雷磁仪器厂;722可见分光光度计 上海棱光技术有限公司;CL-22M高速冷冻离心机 赛特湘仪离心机仪器有限公司;立式压力蒸汽灭菌器 上海博迅实业有限公司医疗设备厂;μBondasphere C18(3.9mm×150mm)色谱柱 美国Waters公司;YMC-Pack ODS-AP-303(4.6mm×250mm)色谱柱 、TSK- Octadecyl 4PW(3.9mm×150mm)色谱柱、YMC-Pack ODS-100S (3.9mm×150mm)色谱柱 日本YMC公司;Discovery Studio 2.1 (DS 2.1)软件包 创腾科技有限公司。
1.3 方法
1.3.1 酸乳的制备及预处理
取10g/100mL脱脂乳在90℃条件下杀菌15min,杀菌结束后冷却到37℃,无菌操作条件下,接种体积分数4.0%的Lactobacillus helveticus JCM1004和 Lactobacillus casei subsp. casei ATCC393所制作的发酵剂,在40℃条件下培养发酵10h,培养发酵结束后用50%的乳酸将其pH值调节至3.4~3.6,8000r/min离心10min,收集上清液,再用10g/100mL的氢氧化钠溶液调pH8.3,8000r/min离心10min,收集上清液冷冻干燥后,用于抑制ACE活性及多肽含量的测定。多肽含量的测定采用Church等[7]的方法。
1.3.2 抗高血压肽的分离[8]
将酸乳冻干粉溶于洗脱液A(乙腈-水-三氧乙酸的体积比100:900:1),再用孔径为0.2μm的过滤膜过滤,除去大分子物质,滤液再经四步反相高压液相色谱[8]分离精制得抗高血压活性肽。将1.0mL上述滤液通过以下4个色谱柱,μBondasphere C18采用梯度洗脱,在0~40min洗脱时间内,洗脱液B(乙腈-水-三氧乙酸体积比100:900:1)的体积分数从0%上升至45%,在41~70min洗脱时间内,B的体积分数从45%升至75%,在71~90min洗脱时间内,B的体积分数从75%升至100%。YMCPack ODS-AP-303、YSK-Octadecyl 4PW色谱柱的洗脱液、洗脱梯度同上。将第3个色谱柱所得活性较高样品溶于洗脱液A(乙腈-水-三氟乙酸体积比100:900:0.8),过YMC-Pack ODS-100S色谱柱,洗脱液B(乙腈-水-三氟乙酸体积比100:900:0.8)梯度如下,在0~45min洗脱时间内,从0%升至40%,在46~75min洗脱时间内,从41%升至70%,在76~90min洗脱时间内,从71%升至100%。各阶段洗脱液的流速均为1.0mL/min,检测波长为215nm,收集各个峰的洗脱液,冷冻干燥后进行抑制ACE活性实验。
1.3.3 抑制ACE活性测定
抑制ACE活性的测定采用Cushman等[9]的方法做适当改进[6,10]。用含有0.3mol/L NaCl的0.1mol/L硼酸盐缓冲液(pH8.3)将Hip-His-Leu配成5.0mmol/L的溶液。在10mL试管中分别加入200μL的5mmol/L Hip-His-Leu溶液和80μL的酸乳上清液(或溶于蒸馏水的多肽溶液),于37℃条件下保温3min后,再加入20μL ACE溶液(溶解于蒸馏水中,活力为0.1U/mL),混匀后在37℃下保温30min,再加入250μL的1.0g/100mL盐酸溶液以终止反应,再加入1.7mL醋酸乙酯,经15s振荡混匀后,静置5min,用移液管吸取1.0mL的醋酸乙酯层,真空冷冻干燥后,加入1.0mL蒸馏水,混匀后在228nm波长处测定吸光度。在上述条件下,抑制50%的ACE活性(IC50)为样品的一个活性单位。IC50为抑制50%的ACE活性所需抑制剂的浓度。

式中:A为含有酸乳乳清(或溶于蒸馏水的多肽溶液)和ACE溶液的吸光度;B为不含发酵乳乳清(或溶于蒸馏水的多肽溶液)样品,但含有ACE溶液;C为含脱脂乳乳清(或蒸馏水)和ACE溶液。
1.3.4 肽的氨基酸组成及序列测定
将多肽溶解于6g/100mL HCl中,在真空下于110℃条件下水解24h,用高压液相色谱系统测定其氨基酸的组成,肽的氨基酸序列分析采用序列分析仪。
1.3.5 分子模拟方法
分子力学与分子动力学模拟采用Discovery Studio 2.1 (DS 2.1)软件包,在CHARMm力场下完成[11-16]。ACE结构来自PDB结构数据库(PDB code: 1UZE.pdb),将ACE配合物中的抑制剂分子依那普利(enalaprilat)和水分子去掉,保留辅因子Zn+和Cl-得到ACE对接结构。将小肽对接范围设定为8A,用Flexible Docking工具将配体(VPP,IPP)柔性对接到受体ACE中,通过比较对接结果的打分值来获得最终的稳定构象,最后计算配体-受体配合物的相互作用结合能。
2 结果与分析
2.1 酸乳中ACE抑制肽的分离鉴定
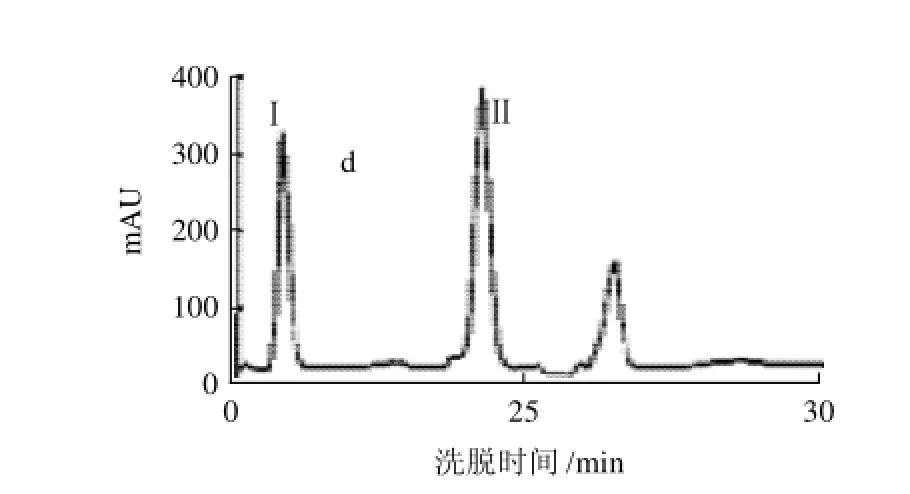
图1 酸奶中ACE抑制肽的反相高压液相色谱图Fig.1 Reversed-phase HPLC elution profiles detected at 215 nm of sour milk
酸乳中抗高血压肽的分离结果如图1所示。上述四步色谱柱分离进行完毕后,色谱图上呈现3个单独吸收峰,经测定,其中峰Ⅰ和峰Ⅱ具有较强的抗ACE活性功能,将这两个峰的收集液分别用其他色谱柱分离均显示单一吸收峰,这表明,经上述四步反相高压相色谱分离得到的样品已比较纯净。且峰Ⅰ和峰Ⅱ的ACE抑制活性分别为419U和496U,而原冻干酸乳粉为1342U,峰Ⅰ和峰Ⅱ抗ACE活性回收率之和占原分离样品(酸乳粉)抗ACE活性的68.18%,由此表明酸乳的抗ACE活性主要是峰Ⅰ和峰Ⅱ所含活性肽所起的作用。

表1 ACE活性抑制物质的氨基酸序列及其IC50值Table 1 Amino acid sequence of angiotensin I-converting enzyme (ACE) inhibitor and the concentration of ACE inhibitor need to inhibit 50% of the ACE activity (IC50)
由表1肽的氨基酸组成及序列分析可知,峰Ⅰ的氨基酸组成为Val 1.0和Pro 1.95,峰Ⅰ为Val-Pro-Pro (VPP);峰Ⅱ的氨基酸组成为Ile 1.0和Pro 2.06,它为Ile-Pro-Pro(IPP)。VPP和IPP的IC50值分别为8.89μmol/L和5.17μmol/L。VPP和IPP的分子结构如图2所示。
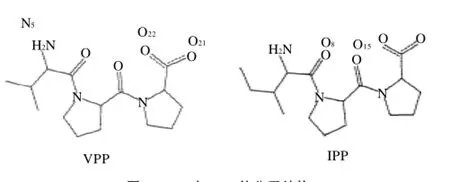
图2 VPP与IPP的分子结构Fig.2 Chemical structural formula of VPP and IPP
2.2 ACE抑制肽的分子模拟
为了在分子水平上了解ACE抑制肽与ACE相互作用的机理,用DS 2.1软件包Flexible Docking模块,采用分子动力学柔性对接的方法模拟相互作用的具体情况。对接结果以LibDockScore、LigScore1、LigScore2、PLP1、PLP2、Jain、PMF、PMFO4等打分函数的打分,同时综合考虑配体与受体形成氢键的数目以及二者相互作用能量的高低,来判断最稳定的构象。经计算总共获得了64种不同构象,以打分最高的前20种构象为研究对象,选取最稳定的构象(VPP和IPP)如图3、4所示。

图3 VPP与ACE的相互作用图Fig.3 Interaction schemes of VPP and ACE
研究表明,VPP与ACE活性口袋周围的氨基酸残基存在氢键、疏水、亲水等作用。在VPP与ACE的对接结果中,按LibDockScore降序排列,再综合比较各个打分函数的打分值,构象10是最优构象,如表2所示。

表2 VPP与ACE之间形成氢键的键长与键角Table 2 Length and angle of hydrogen bond between VPP and ACE
VPP与ACE活性口袋之间形成3个氢键,ACE 356位的丙氨酸与VPP的N5原子通过H32形成氢键,键长2.810A,键角127.0°;522位的精氨酸的NH1与VPP的O22原子通过HH12形成氢键,键长2.859A,键角115.6°;精氨酸的NH1与VPP的O21原子通过HH12也形成氢键,键长2.793A,键角167.8°,可以看出,3个氢键中,通过O21原子形成的氢键键长最短,键角最大,作用力最强,在VPP与ACE的相互作用中起着关键作用。总之,VPP与ACE所形成的氢键较强。VPP中的缬氨酸、脯氨酸都为疏水性氨基酸,与ACE活性口袋周围的疏水性氨基酸形成疏水作用力。VPP分子中的亲水性基团(氨基、羰基、羧基)与ACE活性口袋周围的大量亲水性氨基酸形成很强的亲水作用力。氢键、亲水、疏水等作用力共同稳定了VPP与ACE复合物的构象(图4)。
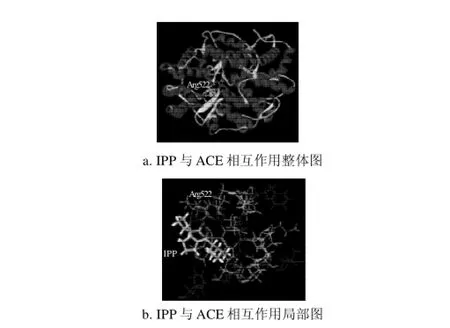
图4 IPP与ACE的相互作用图Fig.4 Interaction schemes of IPP and ACE
在IPP与ACE的对接结果中,按LibDockScore降序排列,再综合比较各个打分函数的打分值,构象11是最优构象。

表3 IPP与ACE之间形成氢键的键长与键角Table 3 Length and angle of hydrogen bond between IPP and ACE
如表3所示,IPP与ACE活性口袋之间也形成3个氢键,ACE 522位精氨酸的NH1与IPP的O8原子通过HH12形成氢键,键长2.729A,键角99.8°;同时,精氨酸的NH1还通过HH12与O15形成氢键,键长2.939A,键角144.7°;精氨酸的NH2通过HH22与O15形成氢键,键长3.039A,键角139.5°。3个氢键中,综合键长键角的情况,NH1与O15原子形成的氢键作用力最强,在IPP与ACE的相互作用中起着关键作用。总之,IPP与ACE形成的氢键作用较强。IPP中的异亮氨酸和脯氨酸均为疏水性氨基酸,与ACE活性口袋周围的疏水性氨基酸形成了疏水作用力。IPP分子中的亲水性基团(氨基、羰基、羧基)也与ACE活性口袋周围的大量亲水性氨基酸形成了很强的亲水作用力。氢键、亲水、疏水等作用力共同稳定了IPP与ACE复合物的构象。
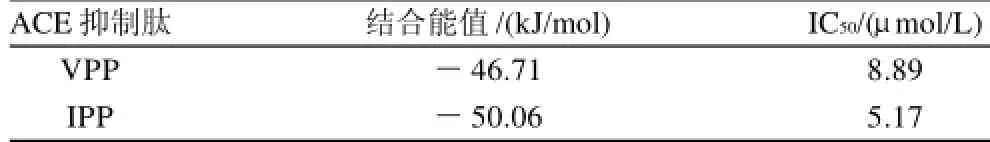
表4 生物活性肽与ACE活性位点相结合,通过分子对接所产生的最稳定构象的结合能值Table 4 Affinity energy values for the best poses obtained by automated molecular docking of bioactive peptides (VPP and IPP) at the ACE-binding site
VPP与IPP都为三肽,无论在体内还是体外,都具有较强的ACE抑制活性。它们只有第一个氨基酸残基是不同的,结构极其相似(图2),这就决定了它们与ACE作用的相似性,它们都与ACE 522位的精氨酸形成很强的氢键,可见ACE 522位的精氨酸为其活性口袋内的关键氨基酸,在配体与受体的结合中起着重要的作用。VPP与IPP结构中,IPP第一个氨基酸中比VPP多一个甲基,这可能是其性质略有差异的主要原因。另外,结合能是衡量一组配体和目标受体结合强弱的最重要的参数,如表4所示,VPP最稳定构象(构象10)的结合能为-46.71kJ/mol,IPP最稳定构象(构象11)的结合能为-50.06kJ/mol,表明配体VPP、IPP与受体ACE之间均有很强的相互作用。但IPP的结合能值较VPP更低,表明IPP与ACE的结合比VPP与ACE的结合更稳定。这也与实验中所得出的IPP比VPP具有较低的IC50的实验结果相一致。
3 结 论
以脱脂乳为原料,通过四步反相高压液相色谱从Lactobacillus helveticus和Lactobacillus casei subsp. casei制作的酸乳中分离到两种抗高血压肽VPP和IPP。经测定,它们抑制ACE活性的IC50分别为8.89mol/L和5.17mol/L;根据氨基酸序列分析的结果,构建了VPP和IPP的分子结构,通过分子柔性对接方法研究了它们与ACE相互作用的分子机理,确定了它们的作用位点、作用力类型及相互作用能。结果表明,VPP、IPP和ACE的活性口袋之间均形成三个氢键,且存在较强的疏水、亲水等作用力,相互作用的结合能分别为-46.71、-50.06kJ/mol,IPP与ACE之间的结合较VPP更稳定,与IPP比VPP具有较低的IC50值的实验结果相一致。
[1]吴炜亮, 吴国杰, 梁道双, 等. ACE抑制肽的生理功能和研究进展[J].现代食品科技, 2006, 22(3): 251-254.
[2]张艳, 胡志和. 乳蛋白源ACE抑制肽的研究进展[J]. 食品科学, 2008, 29(10): 634-640.
[3]FITZGERALD R J, MEISEL H. Milk protein-derived peptide inhibitors of angiotensin-I-converting enzyme[J]. British Journal of Nutrition, 2000, 84(Suppl1): 33-37.
[4]赵树平. 乳杆菌发酵乳中ACE抑制活性和γ-氨基丁酸的研究[D]. 呼和浩特: 内蒙古农业大学, 2008.
[5]HAQUE E, CHAND R. Antihypertensive and antimicrobial bioactive peptides from milk proteins[J]. Eur Food Res Technol, 2008, 227(1): 7-15.
[6]郭宇星. 微生物发酵法水解乳清粉制备生物活性肽的研究[D]. 天津:天津商业大学, 2006.
[7]CHURCH F C, SWAISGOOD H E, PORTER D H, et al. Spectrophotometric assay usingo-phthalaldyhyde for determination of proteolysisin milk an disolated milk protein[J]. J Dairy Sci, 1983, 66(6): 1219-1227.
[8]潘道东. 酸乳中抗高血压肽的分离及其特性研究[J]. 食品科学, 2005, 26(1): 205-210.
[9]CUSHMAN D W, CHEUNG H S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung[J]. Biochem Pharmacol, 1971, 20(7): 1637-1648.
[10]田俊英, 潘道东, 郭宇星. 瑞士乳杆菌蛋白酶水解酪蛋白制备血管紧张素转化酶抑制肽的条件优化[J]. 食品科学, 2009, 30(11): 170-174.
[11]毛慧, 蔡炳锋, 赵波, 等. 5-硝基-1H-吲哚-2-羧酸与牛血红蛋白相互作用的分子模拟与光谱研究[J]. 化学研究与应用, 2009, 21(5): 675-679.
[12]毛慧, 蔡炳锋, 赵波, 等. 苏丹红Ⅱ与肌红蛋白相互作用的分子模拟与实验[J]. 应用化学, 2009, 26(11): 1332-1335.
[13]PINA A S, ROQUE A C. Studies on the molecular recognition between bioactive peptides and angiotensinconverting enzyme[J]. Mol Recognit, 2009, 22(2): 162-168.
[14]赵钰岚, 许传莲. 血管紧张素转换酶的结构功能及相关抑制剂[J]. 生物工程学报, 2008, 24(2): 171-176.
[15]王伟, 沈生荣, 冯凤琴, 等. 基于药效团的血管紧张素转换酶抑制肽的结构优化[J]. 中国科学, 2008, 38(4): 357-364.
[16]郑清川, 楚慧郢, 牛瑞娟, 等. 人类乙酰胆碱酯酶(AChE)与抑制剂小分子的对接研究[J]. 中国科学B辑: 化学, 2009, 39(11): 1454-1460.
Molecular Mechanism Study of Targeting of Two Angiotensin-converting Enzyme Inhibitory Peptides
GUO Hui-qing1,MAO Hui1,ZHAO Bo1,PAN Dao-dong1,2,*
(1. Branch Center of National Dairy Processing Technology Developing, Nanjing Normal University, Nanjing 210097, China;2. Faculty of Life Science and Biotechnology, Ningbo University, Ningbo 315211, China)
Two angiotensin-converting enzyme inhibitory peptides VPP and IPP were purified from the sour milk fermented with Lactobacillus helveticus JCM1004 and Lactobacillus casei subsp. casei ATCC393 by using four-step reverse-phase HPLC. VPP and IPP had 50% inhibition of ACE (IC50) of 8.89μmol/L and 5.17μmol/L, respectively. According to their amino acid sequence, the molecular structure of VPP and IPP were simulated, and the molecular mechanism for the function position, type and energy of the interaction between ACE inhibitory peptides and ACE were investigated by the flexible molecule docking technology. It was shown from the theoretical calculation that the three hydrogen bonds existed between the bioactive peptides poses of VPP and IPP. Hydrophobic and hydrophilic interactions also existed between residues at the ACE active site and the peptide poses. The binding between IPP and ACE was more stable than VPP with ACE. This is in good agreement with the experimental result that the IC50value of IPP is lower than that of VPP.
VPP;IPP;ACE inhibitory peptides;molecular modeling
Q518.3
A
1002-6630(2010)23-0001-05
2010-03-18
国家“863”计划项目(2007AA10Z320);江苏省自然科学基金项目(BK2009403);宁波市自然科学基金项目(2009A610180)
郭慧青(1983—),女,硕士研究生,主要从事乳品科学研究。E-mail:qqnjnu@163.com
*通信作者:潘道东(1964—),男,教授,博士,主要从事乳品科学研究。E-mail:daodongpan@163.com
猜你喜欢
高中数理化(2022年16期)2022-09-14
青岛大学学报(工程技术版)(2019年2期)2019-09-10
中学生数理化(高中版.高考理化)(2019年6期)2019-06-22
温州大学学报(自然科学版)(2016年1期)2016-10-27
中国酿造(2016年12期)2016-03-01
应用化工(2014年7期)2014-08-09
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年15期)2014-03-11
无机化学学报(2014年5期)2014-02-28
