
苏云金芽孢杆菌生物杀虫剂发酵生产的影响因素及其工艺选择
2010-08-29 06:56常明孙启宏周顺桂倪晋仁
生态环境学报 2010年6期
常明,孙启宏,周顺桂,倪晋仁
1. 中国环境科学研究院,北京 100012;2. 北京大学环境工程系//教育部水沙科学重点实验室,北京 100871;3. 广东省农业环境综合治理重点实验室//广东省生态环境与土壤研究所,广州 510650
苏云金芽孢杆菌(Bacillus thuringiensis, Bt)简称苏云金杆菌,是一类能在代谢过程中产生芽孢和伴孢晶体的革兰氏阳性细菌,其制剂是目前世界上产量最大的微生物杀虫剂[1]。自1901年日本学者石渡繁胤(Ishiwata)从病死家蚕中分离出第一株苏云金芽孢杆菌以来,全世界已分离出了 71个血清型86个亚种。其杀虫谱已由无脊椎动物节肢动物门中的鳞翅目扩大到双翅目、鞘翅目、直翅目等9个目的昆虫;同时还发现对螨类、线形动物门中的动植物寄生线虫、原生动物门中的鞭毛虫、变形虫和草履虫以及扁形动物门中的扁虫、吸虫、绦虫等有特异毒性的菌株[2]。目前不少亚种已被成功应用于农业,用来防治粮食作物、蔬菜、棉花、大豆、烟草、果树等多种害虫。
Bt制剂的主要杀虫活性成分是伴孢晶体(亦称杀虫晶体蛋白或 δ-内毒素)。杀虫晶体蛋白(Insecticide Crystal Proteins, ICPs)可占培养物生物量总干重的 20%~30%,分子量在 27~150 kDa之间[3]。Bt的杀虫机制为:伴孢晶体被昆虫吞食后,在中肠的碱性环境和蛋白酶的作用下δ-内毒素被分解与激活,成为具有杀虫活性的毒性肽。毒性肽与幼虫肠上皮细胞膜的专一受体结合,导致膜穿孔,肠道内溶物渗入血腔,同时芽孢趁机侵入增殖,引起足以致死的败血症,导致昆虫全身麻痹或痉挛而死[4]。近几十年来,人们对Bt的发酵生产展开了大量研究,并已形成了较为成熟的生产工艺。本文提炼了国内外众多研究者的实践经验,希望通过对发酵影响因素的分析与发酵工艺发展的探讨为 Bt生物杀虫剂的进一步开发与应用提供参考。
1 苏云金杆菌发酵生产的影响因素
1.1 培养条件
1.1.1 温度
Bt的生长代谢与温度密切相关。温度直接影响细胞内酶的形成与活性,关系到细胞对营养物质的吸收与利用,并影响到菌体数量与伴孢晶体质量。大量研究表明,Bt的最适生长温度为28~32 ℃,发酵时温度过高,会导致Bt毒素基因丢失或蛋白失活,毒效降低;温度过低,则生长缓慢,发酵周期延长[5]。另外,细胞生长最适温度与晶体蛋白中各基因合成的最适温度有时会存在一定差异。Melek Özkan等人考查了温度对Bt subsp. israelensis HD500蛋白基因合成的影响,结果发现Cry4Ba在25 ℃时含量最高,而菌体生长及Cry11Aa合成的最适温度分别是37 ℃和30 ℃[6]。不同苏云金亚种甚至同一亚种的不同菌株,其最佳发酵温度亦有所不同,发酵时需针对菌株特性选择最具经济效率的温度。
1.1.2 pH值
pH不仅影响芽孢的萌发,也影响芽孢的形成。研究表明,初始pH值为7.0左右时,芽孢萌发率最高,当pH值<6.5或>8.0时,萌发率均在45%以下[1]。发酵过程中,Bt先利用碳源增殖,糖代谢产生大量有机酸,pH下降,随后在酶的作用下,有机酸进一步被异化,pH值逐渐回升,芽孢与晶体开始形成。伴孢晶体的蛋白质转化合成过程需要酶的参与,而这种酶的合成与酶活最适pH均为中性,pH值在6.5以下或9.1以上该酶的活性急剧下降。若pH不能回升,芽孢和晶体难于形成[7],若pH值过早升高,又会使营养体增殖提前结束,影响到芽孢和晶体的产量。另外,过高的碱性对伴孢晶体还有一定的溶解作用,会降低发酵液的毒效。发酵过程中可以流加缓冲液,或采用酸碱进行调解,以控制最佳pH值。Khanh Dang Vu等人采用NaOH/H2SO4、NaOH/CH3COOH、NH4OH/CH3COOH和NH4OH/H2SO44种酸碱组合对淀粉废水培养Bt var.kurstaki HD-1的发酵过程进行调节,使pH值保持在7.0±0.1,结果发现以NaOH/H2SO4和NH4OH/H2SO4为pH调节剂对碱性蛋白酶和淀粉酶的产生十分有利,而以NaOH/CH3COOH为pH调节剂可显著提高晶体蛋白产量和毒效[8]。由此可见,采用适宜的pH调节剂,对Bt的发酵过程进行有效调控,是Bt发酵生产的关键因素。
1.1.3 搅拌速度与通气量
Bt是一种好气性芽孢杆菌,在分解和利用发酵基质以及合成自身物质时要消耗大量氧。如:1摩尔葡萄糖完全氧化为CO2和H2O需要6摩尔氧。发酵培养基中糖的含量可达1%,但氧的含量却很低,饱和浓度也只有0.007%。因此随着细菌的生长,必须不断补充大量的氧才能满足菌体的正常生长。同时氧的存在也可提供一个较高的氧化还原电位,维持细胞内氧化酶系的活性[1]。毒素合成机制也受供氧影响[9],Foda等人通过改变装液量考查了供氧对Bt subsp. entomocidus HD-635的影响。发现供氧受限时(空气:培养基=3∶2)菌体不易存活且难于产生芽孢[10]。工业发酵通常以每分钟通入的空气体积来衡量通气量。在液体深层发酵过程中,搅拌速度和通气量显得尤为重要,搅拌速度通常为400~600 r·min-1,通气量一般控制在 1∶0.6~1.2(发酵培养基体积与每分钟通入空气的体积之比)。通气量过大易产生大量泡沫,从排气口冲出,导致污染,同时也影响氧传递,因此在发酵液中添加消泡剂是十分必要的。在发酵的不同阶段,通气量可以适当调节。发酵前期,菌数低,耗氧少,通气量为1∶0.6~1.0即可;进入对数期,耗氧增大,通气量需提高为1∶1.0~1.2[1]。同时配合搅拌以强化供氧,为防止细胞受损,不宜采用剪切式搅拌。当芽孢形成以后,虽然菌体代谢活性降低,但氧对芽孢的成熟和伴孢晶体的形成有重要影响,因此后期仍应保持较大的通气量,可略低于对数期通气量[9]。
1.1.4 接种
工业生产过程中通常采用3级发酵培养,即摇瓶→种子罐→发酵罐。从种子罐到发酵罐通常有 2种接种方式:芽孢接种和营养体接种。芽孢接种可杀灭杂菌及噬菌体,获得较高的同步率,但芽孢萌发慢,延迟期较长。严格控制灭菌条件,避免污染,采用活性营养体接种芽孢产率可能更高。Pearson和Ward[11]在研究Bt subsp. israelensis的接种条件时,选择有抑制芽孢形成能力的培养基作为种子培养基,这种营养体接种方式能有效提高同步率与细菌总数,芽孢产率在90%以上。Bt对培养基的适应能力不同,在发酵培养基中驯化后再作为种子接种往往能缩短延迟期[12]。另外 Wu和 Chen[13]在研究中还发现接种时机能明显影响毒素产量,在菌体活性最强时(延迟期末,对数期初)接种,Bt subsp.darmstadiensis HD-199的外毒素产量比指数末期接种提高了3倍。
1.2 培养基组分
1.2.1 碳源
Bt是一类化能有机营养型细菌,只能利用有机含碳化合物作为碳源和能源。自然界的含碳化合物种类繁多,并非都能被 Bt利用,常用作碳源培养Bt的主要是糖类[1]。单糖如葡萄糖可直接被 Bt吸收,多糖需经菌体产生的胞外淀粉酶水解成葡萄糖等单糖再被吸收。培养基中含有适量葡萄糖,可以促进菌体生长,缩短发酵周期。但在工业生产中培养基灭菌时,受Fe2+、PO43-的影响,葡萄糖易焦糖化,需单独灭菌,使操作过程变得复杂[14];而且由于葡萄糖代谢产酸,大量葡萄糖会使pH值迅速下降,产生碳源抑制,使代谢进程受阻[15]。因此淀粉、糊精、糖蜜和甘油等多糖都被用作 Bt发酵的常用碳源,它们可以克服葡萄糖代谢过快的弊病,保持发酵后期有一定的糖源。Yasemin Icgen等人考查了各种碳源对 Bt的影响,研究发现蔗糖、乳糖和菊粉作碳源时毒效较高,同时也支持芽孢形成,乳清及糖蜜对毒素合成有促进作用,而其他碳源包括葡萄糖、甘油、麦芽糖、淀粉和糊精都得到较低的毒素量[16];Melek Özkan等人则发现菊粉、糊精、麦芽糖、乳糖、蔗糖、乳清、甘油等对 Bt subsp. israelensis HD500的毒效有促进作用,而葡萄糖、淀粉和糖蜜用作碳源时毒素产量低[6]。可见不同菌株对碳源种类及营养需求存在差异。
1.2.2 氮源
工业上常用的氮源有黄豆粉、棉籽饼粉、花生饼粉、玉米浆等,其中玉米浆还有较强的缓冲能力,对Bt的生长有利。Salama等人采用棉籽饼粉、鱼粉、牛血等农副产品以及一些当地可得的廉价高蛋白废渣,如饲料酵母、鸡舍垫料、各类豆渣等,采用 Bt subsp. kurstaki HD-1、Bt subsp. kurstaki HD-73和Bt subsp. entomocidus等菌株进行发酵[17],结果发现饲料酵母为 Bt生长的最适氮源,三株菌的芽孢数都可达到1×109CFU·mL-1以上,其次为牛血;而棉籽饼粉更适合培养Bt subsp. kurstaki HD-73,鸡舍垫料对 Bt subsp. kurstaki HD-1和 Bt subsp.entomocidus生长较为有利。当制剂浓度为500µg/ml时,采用饲料酵母、牛血、鸡舍垫料作为氮源的Bt subsp. kurstaki HD-73与Bt subsp. kurstaki HD-1制剂对棉铃虫(H. armigera)的致死率均可达80%~100%,以各种豆渣作为氮源培养的Bt subsp.entomocidus对棉贪夜蛾(S. littoralis)的致死率也可达90%左右。可见不同菌株对氮源种类具有选择性,而不同氮源又影响同一菌株对各类昆虫的毒效。因此在每一个新菌株用于生产之前,都需要进行最适培养基的选择。无机氮源通常难以被利用,但新近研究表明,无机氮能增加蛋白水解活性,蛋白水解活性可进一步影响晶体蛋白的稳定性。Yasemin Icgen等人的研究还表明无机氮源能影响Bt的蛋白组成(该菌株可合成135 kDa Cry I及65 kDa Cry II两种有效蛋白)。当(NH4)2SO4质量浓度从0~2 g·L-1时,65 kDa蛋白随之浓度增加而提高,用NH4Cl替换(NH4)2SO4则导致135 kDa蛋白含量降低,而改用KNO3可导致65 kDa蛋白消失[18];Melek Özkan[6]等人也发现(NH4)2HPO4用作无机氮源时能使Cry11Aa和Cry4Ba蛋白产量提高。目前对无机氮源的研究还在不断深入。
1.2.3 碳氮比
作为微生物生长最重要的营养源,碳、氮源充足是微生物正常生长代谢的必要条件,但同步增加碳、氮源并不能有效提高生物量与毒效,浓度过高还会产生底物抑制,研究发现碳氮比在其中起关键作用。首先,碳氮比会明显影响发酵体系的 pH,高碳氮比培养基的pH在整个代谢过程中不会上升太高,最终停留在7.5左右;而低碳氮比时发酵终点的pH可上升至8.5以上[19]。通常低碳氮比培养基毒素产量略高,但毒素产量还不能完全理解为毒效。Mummigatti和Raghunathan 研究时就遇到了这种情况,尽管低碳氮比时芽孢数及毒素产量略高,但毒效较低;而 Lachhab[21]等人在研究污泥发酵培养Bt HD-1时也发现较高的碳氮比对提高毒效有利,当碳氮比在 8~10左右时,杀虫毒效较好。因此在选择碳、氮源时,不可一味追求生物量与毒素产量,需结合生物毒效确定合理的碳氮比。
1.2.4 矿质元素
Bt需要的矿质养料可分为常量元素和微量元素两大类。常量元素包括磷、镁、钠、钾和钙,主要参与细胞组成、能量转移、物质代谢以及调节细胞原生质状态或细胞通透性等[1]。磷是组成核酸和磷脂的成分,也是苏云金素的重要组成部分,许多酶的活性基中都有磷的参与。常用的磷盐主要是K2HPO4和KH2PO4,在提供磷的同时也补充了钾。Melek Özkan等人报道较高浓度的无机磷对合成Cry4Ba很有效,当培养基中含50~100 m M K2HPO4时,Cry4Ba和Cry11Aa以及芽孢产率均显著提高。磷酸盐对细胞和环境中的酸碱度还起到缓冲作用[6],常用作生测时细胞及晶体的保护剂。镁是核糖体和膜结构的稳定剂,能激活酶反应,对重金属引起的细胞毒害还有一定的拮抗作用。Yasemin Icgen等发现镁影响135 kDa和65 kDa毒性蛋白组分的生物合成,当Mg2+浓度在8×10-5~4×10-3M时,生长、芽孢形成及晶体蛋白合成量都很高[18]。钾的主要作用是维持渗透压和离子平衡。Wakisaka等证实在Bt subsp. kurstaki和aizawai培养基中加入钾盐比钠盐有助于δ-内毒素的形成,当K+浓度达0.20 M时,产量及内毒素活性较高[22]。钙不参与细胞结构组成,而是以离子状态控制细胞的生理状态,如降低细胞质膜的通透性,调节酸碱度,并对一些阳离子的毒性有拮抗作用。与磷类似,钙盐也能起到缓冲作用,培养基中加入适量的CaCl2或CaCO3可缓和pH 变化[23]。
微量元素有锰、锌、硅、铜和铁,它们多为辅酶或辅基。Bt对微量元素的需要量很少,通常在水和其他养料成分中作为杂质存在的含量足以满足其需要。铁是细胞色素、细胞色素氧化酶和过氧化氢酶的活性组成成分;锌是乙醇脱氢酶的辅基,许多酶的活性也靠锌激活[1];锰也是多种酶的激活剂,浓度达到10-6M时对Cry4Ba和Cry11Aa毒素产量即有明显影响[6],Yousten’s培养基中含3×10-4M锰对生长及毒素合成最有利。铜也有一定激活作用,加入10-7M时晶体蛋白合成明显受到激发,增加至10-6和 10-5之间时,可导致晶体蛋白产量降低,适宜控制铜浓度可使芽孢数比不加铜时提高3倍。此外,Abrosimova等人发现增加矿物盐的培养基有更好的耐热性能,这可能是由于盐组分(尤其是钾盐)能明显提高细胞的护温屏障[24]。
总之,在 Bt发酵生产中,这些矿质元素的加入量一般为:KH2PO4或K2HPO4为0.075%~0.2%;MgSO4·7H2O 为 0.075%~0.3%;CaCO3为0.075%~0.15% ; MnSO4·H2O 、 FeSO4·7H2O 均 为0.002%。
1.2.5 其它
发酵过程中添加消泡剂等表面活性剂对发酵也有促进作用,Zouari和 Jaoua[25]在培养基中加入了0.5% NaCl和0.1%的Tween-80,发现含固率<59 g·L-1时,Tween-80有助于提高δ-内毒素产量,高于75 g·L-1时,Tween-80在产量提高方面作用不大,但仍能显著降低蛋白水解活性。总之,Bt的微生物发酵是一个复杂的物质及能量转化过程,影响因素较多,且各因素间往往相互影响,需综合调配。表1列出了优化培养基组分及培养条件所得部分代表性成果。
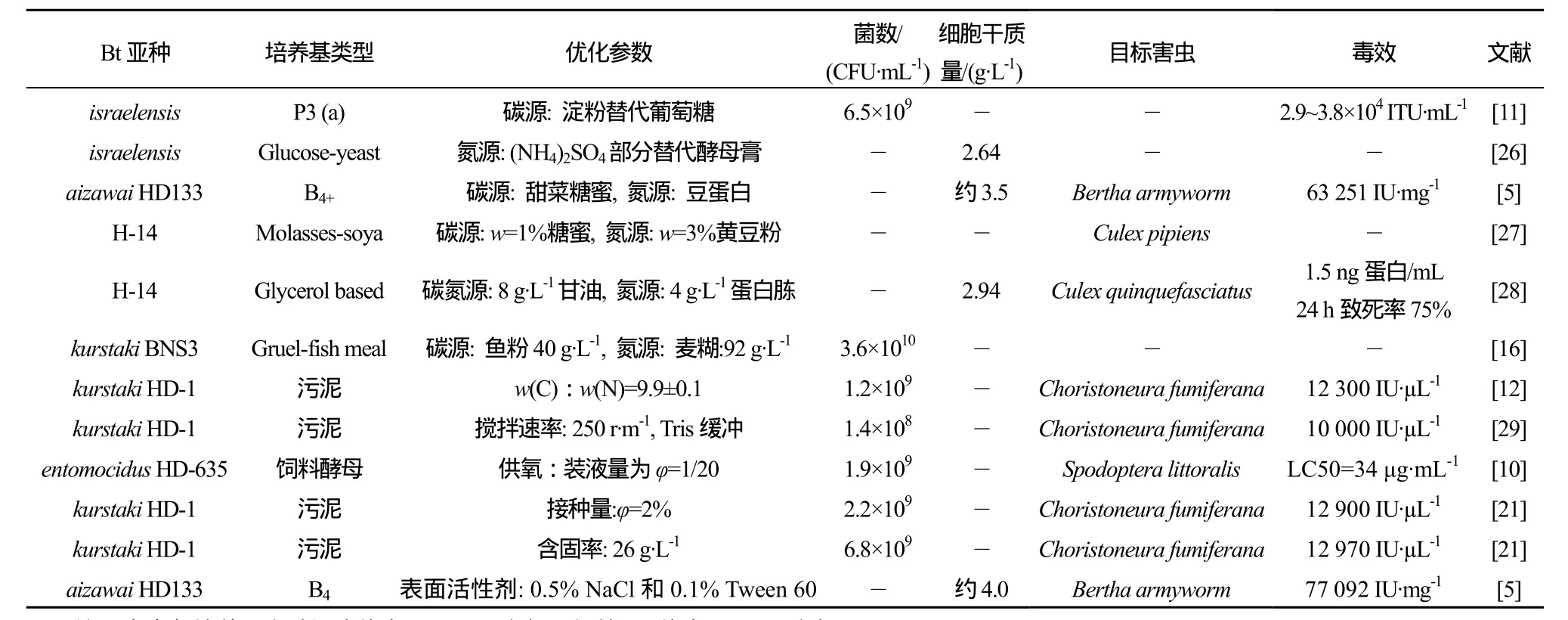
表1 Bt培养条件与培养基组分的优化Table 1 Typical achievements of optimized culture media and culture conditions on Bt fermentation

图1 液体深层发酵基本工艺流程Fig.1 Schematic diagram showing submerged fermentation process of Bt
2 苏云金杆菌的发酵工艺选择
2.1 液体深层发酵:
1956年前苏联发表了用液体培养基摇瓶培养Bt,并用于防治菜青虫的报道[1],从而揭开了Bt液体培养的序幕,Bt制剂之所以能广泛应用,关键在于能通过液体深层发酵大规模生产。其工艺流程如图1所示。
液体发酵主要有分批发酵、补料分批发酵和连续发酵三种方式[30]。分批发酵一次性投料,工艺简单,但若要达到较高的发酵水平,需要较高的基质浓度,这种情况下很容易产生基质和代谢产物抑制[31],同时培养基的粘度增加后,由于影响混合和流动而不利于氧的传递,最终可能使毒效大打折扣。为此人们从反应器和工艺角度进行了改进。采用外环流气升式反应器,通过气体喷射推动液体循环流动以取代传统的机械搅拌方式[32],由于能耗低、结构简单、传质效果好、换热面积大、剪切力低等优点,对Bt毒效的提高有很大帮助,但目前还缺乏大型生产的经验,尚处于研究阶段;提高搅拌速度或增大通风量以改善供氧环境,有助于毒效的提高[33],但势必以增加能耗为代价。于是有人提出了流加工艺,逐渐提高基质浓度以削弱抑制,也因此实现了Bt的连续发酵,但长时间的连续发酵培养基很容易染菌,菌种也易发生退化或产生无孢突变株[34]。综合两者的优点,补料分批发酵被认为有较好的发展前景,即逐渐补料,一次出料。补料方式又可分为连续式和间歇式,Kang等研究发现间歇式补料比连续式细菌增殖快,芽孢密度大;连续式补料在补料过程中即使细胞增殖缓慢也不会像分批发酵那样很快转入芽孢期,补料浓度过高则不能形成芽孢[35]。Zhou等通过控制pH值来调节补料,补料过程中pH保持在7.0左右,避免了营养过剩问题,苏云金素产量比分批发酵提高了89.51%[35]。
2.2 固态发酵:
固态发酵起源于我国传统的“制曲”技术,是利用颗粒载体表面所吸附的营养物质或颗粒本身提供的营养来培养微生物[1]。在相对小的空间内,这些颗粒载体可提供相当大的气液界面,从而满足好气微生物增殖所需要的水份、氧气和营养。20世纪50年代,国外开始将这项技术用于Bt的发酵生产。70年代,我国许多地区与单位都进行了Bt的固态发酵研究,直到上世纪80年代,其生产工艺才逐渐完善。传统的固态发酵按设计规模可分为网盘薄层法、皿箱式、大池通风法以及地坪式等发酵方式[1],如图2所示。
可用于苏云金固态发酵的原料很广泛,但选择时既要考虑物料的营养性,也要考虑载体的通气性。通常使用的载体可分为有机载体和无机载体两类,有机载体如麦麸、米糠、黄豆饼粉、花生饼粉等,既可作为营养源,又兼有载体作用;无机载体如多孔珍珠岩、细沙等,这些物质通气性能较好,但需要另外添加营养成分[31]。杨淑兰等[36]利用麸皮、棉籽饼粉、米糠、草木灰等为发酵原料,进行了百公斤级的Bt subsp. HD-1固态发酵试验,研究发现种龄、发酵温度、初始pH值、基质含水量与草木灰含量等是影响芽孢形成和毒效的重要因素,选择最佳培养条件可使发酵芽孢数稳定在2.0×10 CFU·g-1,在2000倍稀释度条件下对菜青虫的致死率为100%。基质的通气性主要用含水量来控制。适宜的初始含水量,使得培养基有合适的疏松度,颗粒间存在一定空隙,有助于菌体从培养基获得营养成分和氧的传递,从而促进生长繁殖,而过高的含水量会导致培养基粘结成团,多孔性降低,影响氧的传递;含水量过低,则使培养基膨胀程度降低,水的活度低,抑制菌体生长。Capalbo等人将潮湿的稻谷装入聚丙烯袋接种Bt subsp. tolworthi进行了固态发酵实验室研究,湿度控制在50%~60%,产物田间毒效良好,48 h死亡率可达100%[37]。固态发酵具有低投资、低成本、低排污等优点[38],但因输送、搅拌、温度、湿度、pH值和供氧等诸多问题缺少工程解决手段而使其发展受到了限制。陈洪章等人首先提出了压力脉动固态发酵技术,利用压力脉动周期刺激强化生物反应和细胞膜传质速率。压力脉动避免了机械搅拌的缺陷,提高了传质传热效率,降低温度、O2和CO2浓度梯度,促进了毒效的提高。目前,压力脉动固态发酵反应器已成功放大到70 m3的工业级生产规模[39]。
在发酵过程中,液态发酵流动性好,传质、传热性能优于固态发酵,也便于控制;但在后处理过程中,液态发酵通常需要碳酸钙助滤或离心浓缩,操作复杂且有效成分易流失,而固态发酵可以直接进行干燥、粉碎,能源消耗小,但可湿润性较差。可见,工艺都存在各自的优缺点,选用何种发酵工艺还应依据培养基组分与发酵水平,以进行综合调控。
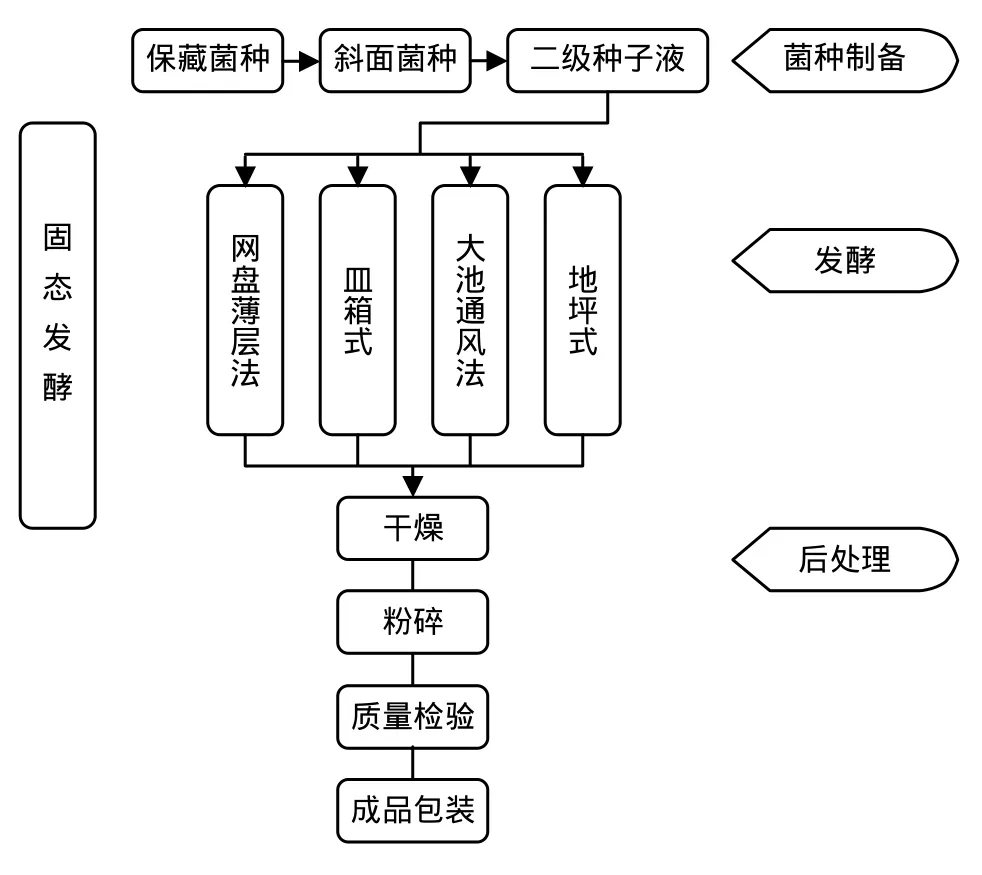
图2 固态发酵基本工艺流程Fig.2 Schematic diagram solid-state fermentation process of Bt
3 展望
化学杀虫剂的弊端引发了人们对生物杀虫剂的关注,而 Bt的研究与应用推动了生物杀虫剂的发展。据专家预测,今后10年内,由于有机农业、森林防治与生物杀蚊剂市场的需求驱动,生物杀虫剂产量将以10%~15%的速率递增[40],Bt生物杀虫剂的研究还有待进一步深入。
(1)原料价格是 Bt生物杀虫剂规模化的制约因素,传统发酵原料价格约占总生产成本的35%~59%[41],因此应致力于开发低成本、当地可得的工农业废弃物为原料,已有学者曾尝试以污泥[42,43]、味精废水[44]、废啤酒酵母浸出液[45]等为发酵原料,这不仅为生物杀虫剂的推广创造了有利条件,还充分利用了废弃资源。目前,低成本培养原料选择与优化还处于起步阶段,如何放大投产是亟待解决的问题。
(2)传统生产工艺已不能满足日益增长的应用需求,高效生产工艺的开发十分迫切。已有研究表明,以提高毒蛋白为目的的高密度补料发酵工艺和压力脉动固态发酵工艺将有广阔的应用前景。
(3)为了进一步提高 Bt的杀虫效果,如延长持效期、扩大杀虫谱等,可以通过对杀虫基因的修饰、改造、转移等基因工程手段构建新型的工程菌[46]。在国外,已有Conder、MVP等10余种Bt工程菌制剂投人了商业应用[47]。在我国,Bt工程菌剂的研发也有扩大趋势,如高效 Bt和荧光假单胞菌的组合基因工程菌剂WG001已于2000年通过安全性审批,更多新型Bt工程菌剂正有待开发与应用。
[1] 喻子牛. 苏云金芽孢杆菌制剂的生产与应用[M]. 北京:农业出版社, 1993.YU Ziniu. Production and application of Bacillus thuringiensis preparations[M]. Agricultural Publishing House, Beijing, 1993.
[2] 鲁松清, 孙明, 喻子牛, 等. 苏云金芽孢杆菌杀重晶体蛋白基因的分类[J]. 生物工程进展, 1998, 18(5): 57-62.LU Songqing, SUN Ming, YU Ziniu, et al. Nomenclature of Bacillus thuringiensis Insecticide Crystal Protein Genes[J]. Chinese Progress of Biotechnology, 1998, 18(5): 57-62.
[3] CHATTOPADHYAY A, BHATNAGAR N B, BHATNAGAR R. Bacterial Insecticidal Toxins[J]. Critical Reviews in Microbiology, 2004,30(1): 33-54.
[4] 格拉泽 AN, 二介堂弘. 微生物生物技术: 应用微生物学基础原理[M]. 陈守文, 喻子牛, 译. 北京: 科学出版社, 2002.GLAZER A N, NIKAIDO H. Microbial biotechnology: fundamentals of applied microbiology[M]. Chen Shouwen, Yu Ziniu, translators.Beijing: Science Press, 2002.
[5] MORRIS O N, CONVERSE V, KANAGARATNAM P, et al. Effect of cultural conditions on spore-Crystal yield and toxicity of Bacillus thutingiensis subap. azawai (HD133)[J]. Journal of Invertebrate Pathology, 1996, 67: 129-136.
[6] MELEK Ö, FILIZ B D, YETIS Ü, et al. Nutritional and cultural parameters influencing antidipteran delta-endotoxin production[J]. Research in Microbiology, 2003, 154: 49-53.
[7] 和致中. 碳、氮、磷三因素对苏云金杆菌的影响[J]. 微生物学通报,1980(7): 7-10.HE Zhizhong. Effects of carbon, nitrogen and phosphorus on Bacillus thuringiensis[J]. Microbiology, 1980(7): 7-10.
[8] VU K D, TYAGI R D, VALÉRO J R, et al. Impact of different pH control agents on biopesticidal activity of Bacillus thuringiensis during the fermentation of starch industry wastewater[J]. Bioprocess and Biosystem Engineering, 2009, 32: 511-519.
[9] AVIGNONE-ROSSA C A, ARCAS C, MIGNONE C. Bacillus thuringiensis growth, sporulation and δ-endotoxin production in oxygen limited and non-limited cultures[J]. World Journal of Microbiology and Biotechnology, 1992, 8: 301-304.
[10] FODA M S, SALAMA H S, SELIM M. Factors affecting growth physiology of Bacillus thuringiensis[J]. Applied Microbiology and Biotechnology, 1985, 22(1): 50-52.
[11] PEARSON D, WARD O P. Effect of culture conditions on growth and sporulation of Bacillus thuringiensis subsp israelensis and development of media for production of the protein Crystal endotoxin[J]. Biotechnology Letters, 1988, 10(7): 451-456.
[12] VIDYARTHI A S, TYAGI R D, VALÉRO J R, et al. Studies on the production of B. thuringiensis based biopesticides using wastewater sludge as a raw material[J]. Water research, 2002, 36: 4850-4860.
[13] WU W T, CHEN S. An index for determining the culture time of inoculum in cultivation of Bacillus thuringiensis[J]. Biotechnology Letters, 2001, 23: 1201-1204.
[14] 余健秀, 汤慕谨, 徐建敏, 等. 苏云金杆菌小试生产发酵影响因子的研究[J]. 中国生物防治, 2002, 18 (1): 17-20.YU Jianxiu, TANG Mujin, XU Jianmin, et al. The Factors Affecting the Fermentation of Bacillus thuringiensis[J]. Chinese Journal of Biology Control, 2002, 18 (1): 17-20.
[15] SAKHAROVA Z V, RABOTNOVA I L, KHOVRYCHEV M P. Growth and spore formation in Bacillus thuringiensis var. israelensis and development of media for production of protein Crystal endo-toxin[J].Biotechnology Letters, 1988, 10(7): 451-456.
[16] YASEMIN I, BÜLENT I, GÜLAY Ö. Regulation of Crystal protein biosynthesis by Bacillus thuringiensis: Ⅱ . Effects of carbon and n itrogen sources[J]. Research in Microbiology, 2002, 153: 605-609.
[17] SALAMA H S, FODA M S, Dulmage H T. Novel fermentation media for production of δ-endotoxins from Bacillus thuringiensis[J]. J Invert Pathol, 1983, 41: 8-19.
[18] YASEMIN I, BÜLENT I, GÜLAY Ö. Regulation of Crystal protein biosynthesis by Bacillus thuringiensis: Ⅰ . Effect of mineral elements and pH[J]. Research in Microbiology, 2002, 153: 599-604.
[19] SACHDEVA V, TYAGI R D, VALÉRO J R. Factors affecting the production of Bacillus thuringiensis biopesticides[J]. Recent Research and Development of Microbiology, 1999 (3): 363-375.
[20] MUMMIGATTI S G, RAGHUNATHAN N. Influence of media composition on the production of δ-endotoxin by Bacillus thuringiensis var.thuringiensis[J]. Journal of Invertebrate Pathology, 1990, 55: 147-151.
[21] LACHHAB K, TYAGI R D, VALÉRO J R. Production of Bacillus thuringiensis biopesticides using wastewater sludge as a raw material:effect of inoculum and sludge solids concentration[J]. Process Biochemistry, 2001, 37: 197-208.
[22] WAKISAKA W, MASAKI E, NISHIMOTO Y. Formation of Crystalline δ-endotoxin or poly-β-hydroxybutyric acid granules by asporogenous mutants of Bacillus thuringiensis. Applied and Environmental Microbiology, 1982: 43.
[23] SIKDAR D P, MAJUMDAR M K, MAJUMDAR S K. Effect of minerals on the production of the delta endotoxin by Bacillus thuringiensis subsp. israelensis[J]. Biotechnology Letters, 1991, 13(7): 511-514.
[24] ABROSIMOVA L I, AVA P V, BREVA G M, et al. Influence of mineral salts on the level of exotoxin production and productivity of a culture of Bacillus thuringiensis[J]. Microbiology, 1986, 55(3): 337-341.
[25] ZOUARI N, JAOUA S. The effect of complex carbon and nitrogen, salt,Tween-80 and acetate on delta-endotoxin production by a Bacillus thuringiensis var. kurstaki[J]. Journal of Industrial Microbiology and Biotechnology, 1999, 23: 497-502.
[26] AVIGNONE-ROSSA C A, YANTORNO O M, ARCAS J A, et al.Organic and inorganic nitrogen source ratio effects on Bacillus thuringiensis var. israelensis delta-endotoxin production[J]. World Journal of Microbiology and Biotechnology, 1990, 6: 27-31.
[27] ABDEL-HAMEED A, CARLBERG G, ELTAYEB O M. Studies on Bacillus thuringiensis H-14 strains isolated in Egypt-Ⅳ . Characteriz ation of fermentation conditions for delta-endotoxin production. World Journal of Microbiology and Biotechnology[J]. 1991, 7(2): 231-236.
[28] MIRTA M F, OSVALDO M Y, HORACIO A, et al. Effect of the media composition on the growth parameters and biological properties of Bacillus thuringiensis var. israelensis delta-endotoxin[J]. World Journal of Microbiology and Biotechnology, 1990, 6: 32-38.
[29] TIRADO M M L, TYAGI R D, VALÉRO J R, et al. Production of biopesticides using wastewater sludge as a raw material-effect of process parameters[J]. Water Science and Technology, 2003, 48(8):239-246.
[30] KANG B C, LEE S Y, CHANG H N. Enhanced Spore Production Of Bacillus thuringiensis By Fed-Batch Culture[J]. Biotechnology Letters,1992, 14 (8): 721-726.
[31] 朱玮, 赵兵, 王晓东, 等. 生物农药苏云金芽孢杆菌的研究进展[J].过程工程学报, 2004, 4(3): 282-288.ZHU Wei, ZHAO Bing, WANG Xiaodong, et al. Progress of the Study on Bacillus thuringiensis as Biopesticides[J]. The Chinese Journal of Process Engineering, 2004, 4(3): 282-288.
[32] HUANG T K, WANG P M, WU W T. Cultivation of Bacillus thuringiensis in an Airlift Reactor with Wire Mesh Draft Tubes[J]. Biochemical Engineering Journal, 2001, 7: 35-39.
[33] DHOUHA G, NABIL Z, HASSEN T. Improvement of Bacillus thuringiensis delta-endotoxin production by overcome of carbon catabolite repression through adequate control of aeration[J]. Enzyme and Microbial Technology, 2007, 40(4): 614-622.
[34] ACRAS J, YANTORNO O, ERTOLA R. Effect of High Concentration of Nutrients on Bacillus thuringiensis Culture[J]. Biotechnology Letters, 1987, 9(2): 105-110.
[35] ZHOU Jingwen, CHANG Yafei, XU Zhenghong. Production of thuringiensin by fed-batch culture of Bacillus thuringiensis subsp. darmstadiensis 032 with an improved pH-control glucose feeding strategy[J]. Process Biochemistry, 2007, 42 (1): 52-56.
[36] 杨淑兰,张怡,梁淑兰,等. 生物杀虫剂苏云金杆菌的固态发酵[J].化工冶金, 1993, 14(2): 162-167.YANG Shulan, ZHANG Yi, LIANG Shulan, et al. Solid fermentation of Bacillus thuringiensis based biopesticide[J]. Journal of Chemical Metallurgy, 1993, 14(2): 162-167.
[37] CAPALBO D M F, MORAES I O, PELIZER L H. Solid-State fermentation of Bacillus thuringiensis tolworthi to control fall armyworm in maize. Electronic Journal of Biotechnology, 2001, 4(2): 112-115.
[38] VIMALA D P S, RAVINDER T, JAIDEV C. Cost-effective production of Bacillus thuringiensis by solid-state fermentation[J] Journal of Invertebrate Pathology, 2005,88(2): 163-168.
[39] 陈洪章, 李佐虎. 固态发酵新技术及其反应器的研制[J]. 化工进展,2002, 21(1): 37-40.CHEN Hongzhang, LI Zuohu. Study on Solid-state Fermentation and Fermenter[J]. Chemical Industry and Engineering Progress, 2002,21(1): 37-40.
[40] TYAGI R D, SIKATI F V, BARNABE S, et al. Simultaneous production of biopesticide and alkaline proteases by Bacillus thuringiensis using sewage sludge as a raw material[J]. Water Science and Technology, 2002, 46(10): 247-254.
[41] The Bacillus thuringiensis Production Handbook[M]. CPL Scientifics Ltd., Newbury, UK: CPL Press, 1993.
[42] 常明, 周顺桂, 卢娜, 等. 微生物转化污泥制备苏云金杆菌生物杀虫剂[J]. 环境科学, 2006, 27(7): 1450-1454.CHANG Ming, ZHOU Shungui, LU Na, et al. Bioconversion of sewage sludge to biopesticide by Bacillus thuingiensis[J]. Environ Sci,2006, 27(7): 1450-1454.
[43] BRAR S K, VERMA M, TYAGI R D, et al. Bacillus Thuringiensis Fermentation of Primary and Mixed Sludge: Rheology and Process Performance[J]. Journal of Environmental Engineering, 2008, 134(8):659-670.
[44] 杨建州, 张松鹏. 利用味精废水发酵生产苏云金芽孢杆菌的发酵条件研究[J]. 食品与发酵工业, 2002, 28 (4): 28-32.YANG Jianzhou, ZHANG Songpeng. Study on the Conditions of Bacillus thuringiensis Fermentation in Monosodium Glutamate Wastewater[J]. Food and Fermentation Industry, 2002, 28 (4): 28-32.
[45] SAKSINCHAI S, SUPHANTHARIKA M, VERDUYN C. Application of a simple yeast extract from spent brewer's yeast for growth and sporulation of Bacillus thuringiensis: a physiological study[J]. World Journal of Microbiology and Biotechnology, 2001, 17: 307-316.
[46] BAIG D N, MEHNAZ S. Determination and distribution of cry-type genes in halophilc Bacillus thuringiensis isolates of Arabian Sea sedimentary rocks[J]. Microbiological Research, 2010, 165(5): 376-383.
[47] KRIEGER R. Hayes' Handbook of Pesticide Toxicology (Third Edition)[M]. UK: Academic Press, 2010: 119-229.
猜你喜欢
上海理工大学学报(2021年3期)2021-07-20
陶瓷学报(2021年1期)2021-04-13
陶瓷学报(2021年1期)2021-04-13
上海农业科技(2019年3期)2019-06-25
江西农业(2018年9期)2018-08-20
河北林业科技(2016年5期)2016-11-08
中国酿造(2016年12期)2016-03-01
导航定位学报(2015年2期)2015-06-05
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年9期)2014-03-11
