
海洋链霉菌Streptomycessp.SCSIO 1672及其代谢产物水杨酸的分离鉴定
2010-09-25 09:30罗明和汪中文黄洪波朱清华田新朋白志川张长生鞠建华
微生物学杂志 2010年6期
罗明和,汪中文,黄洪波,朱清华,田新朋,白志川,张 偲,张长生,鞠建华*
(1.西南大学园艺园林学院,重庆 400700;2.中国科学院海洋生物资源可持续利用重点实验室,广东广州 510301; 3.广东省海洋药物重点实验室,广东广州 510301;4.中国科学院南海海洋研究所,广东广州 510301)
海洋链霉菌Streptomycessp.SCSIO 1672及其代谢产物水杨酸的分离鉴定
罗明和1,汪中文2,3,4,黄洪波2,3,4,朱清华2,3,4,田新朋2,3,4,白志川1,张 偲2,3,4,张长生2,3,4,鞠建华2,3,4*
(1.西南大学园艺园林学院,重庆 400700;2.中国科学院海洋生物资源可持续利用重点实验室,广东广州 510301; 3.广东省海洋药物重点实验室,广东广州 510301;4.中国科学院南海海洋研究所,广东广州 510301)
从南海海洋沉积物中分离得到1株海洋放线菌,鉴定为链霉菌Streptom ycessp.SCSI O 1672。通过优化发酵条件,采用海虾生物致死活性和高效液相色谱追踪,利用有机溶剂萃取、正相硅胶、反相硅胶等各种色谱层析方法分离出活性化合物,通过波谱数据解析出海洋放线菌SCSI O 1672次级代谢产物中的该活性化合物为水杨酸。
海洋放线菌;链霉菌;水杨酸
微生物中的放线菌是药物的重要来源之一。目前在已发现的23000个微生物活性次级代谢产物中有超过10000个来源于放线菌,占了近45%。这些活性物质具有抗肿瘤、细胞毒活性、抗感染及免疫抑制等广泛的生理活性[1]。经过近几十年的研究开发,人们从陆生放线菌中发现新抗生素的机率大大降低[2],于是把探索的目光转向海洋。海洋环境具有低温、高压、高盐等特点,这为海洋放线菌发展独特的代谢途径和机体防御机制并产生结构新颖、活性独特的代谢产物提供了条件。近10 a来,人们从海洋放线菌中发现了大量结构新颖的活性物质。例如,Fenical研究小组从Salinispora tropicCNB-392分离得到的salinosporamide A具有很好的选择性细胞毒活性[3]; Fernández-Chimeno等[4]从M icrom onosporasp.L-25-ES25-008中分离出的大环内酯类化合物I B-96212对小鼠淋巴性白血病细胞P-388有极强抑制活性(IC50=0.0001 mg/L),对肿瘤细胞系A-549、HT-29及MEL-28有明显的细胞毒活性(I C50=1 mg/L);Riedlinger等[5]从Verrucosisporasp. AB-18-032中分离得到了一类多环聚酮类抗生素abyssomicin C,可强烈抑制革兰阳性菌的生长。本文对从中国南海海洋沉积物中分离纯化的240株海洋放线菌发酵产物进行海虾致死活性筛选,发现编号为SCSI O 1672的海洋放线菌发酵粗提物有海虾致死活性。对该菌株的16S r DNA进行分析,鉴定为链霉菌属(Streptom yces)。采用海虾毒性为指标的活性追踪方法,对SCSI O 1672的活性次级代谢产物进行了研究,从中分离得到1个化合物—水杨酸,并对该菌株产水杨酸的发酵培养基进行了初步筛选优化。产水杨酸的链霉菌尚未见文献报道,本文报道该菌株的16S rDNA序列分析,发酵产物中水杨酸的分离鉴定及产水杨酸的培养基的初步筛选。
1 材料与方法
1.1 材料
1.1.1 菌种 海洋放线菌SCSI O 1672分离于中国南海北部沉积物,菌种保存于中国科学院南海海洋研究所。
1.1.2 培养基 实验采用培养基JNP1(可溶性淀粉5 g/L,鱼粉2 g/L,海藻糖2 g/L,几丁质2 g/ L,粗海盐30 g/L)、JNP2(可溶性淀粉10 g/L,酵母膏4 g/L,蛋白胨2 g/L,粗海盐30 g/L)和经优化的培养基AM2a(可溶性淀粉20 g/L,大豆粉10 g/L,蛋白胨2 g/L,酵母膏5 g/L,粗海盐30 g/ L)、AM2ab(可溶性淀粉20 g/L,大豆粉10 g/L,蛋白胨2 g/L,酵母膏5 g/L,CaCO32 g/L,NaCl 4 g/L,粗海盐30 g/L)。培养基配制时调pH 7.0~7.2,121℃灭菌20 min。
1.2 方法
1.2.1 发酵 发酵培养基的优化:将放线菌SCSI O 1672接种于装有50 mL不同发酵培养基的250 mL三角瓶中,28℃200 r/min培养8 d。放大培养发酵;种子培养:将放线菌SCSI O 1672接种于装有50 mL AM2a培养基的250 mL三角瓶中,28℃200 r/min培养50 h;发酵:按10%的接种量将种子接入装有200 mL AM2ab培养基的1000 mL三角瓶中,28℃200 r/min培养7 d。
1.2.2 HPLC分析 50 mL发酵产物加80 mL丁酮,搅拌萃取2 h,静置,取上层丁酮减压蒸馏至干,加DMSO配成20 mg/mL供生物活性测试(HPLC分析时,50 mL发酵物提取物溶解于1 mL甲醇,12000 r/min离心2次)。色谱条件:柱温:室温;洗脱剂:A相为乙腈∶水∶冰醋酸(15∶85∶0.1),B相为乙腈∶水∶冰醋酸(80∶20∶0.1),利用程序设定梯度,0~20 min内A相由100%变为0,20~24 min为100%B相冲洗,流速:1.0 mL/min;检测波长:210 nm,254 nm。取甲醇溶解的发酵产物各20μL进行HPLC(Varian ProStar高效液相色谱仪,配PDA检测器)分析,根据HPLC的吸收峰,收集不同吸收峰的化合物,旋转蒸干后用5μL DMSO溶解。检测不同吸收峰化合物的海虾致死活性,以确定活性峰的tR值及活性峰的峰面积。
1.2.3 活性测试 海虾卵的孵化:取海虾卵100 mg置于500 mL烧杯中,加入含3%海盐的人工海水,28℃光照、通气培养24 h;海虾致死活性测试:取96孔细胞培养板,每孔加200μL含15个海虾幼虫的人工海水液。空白对照组加5μL DMSO,样品组加5μL样品液。28℃光照培养24 h,在双目解剖镜下计数海虾死亡个体数目,并计算死亡率。
1.2.4 化合物的分离分析 将发酵产物用3500 r/min离心10 min,取上清液用2倍体积的丁酮萃取3次,萃取液经减压浓缩,所得浸膏拌样上100~200目正相硅胶柱,用氯仿-甲醇梯度洗脱(10∶0~0∶10),所接流分经HPLC分析,取氯仿∶甲醇为9∶1时的流分,上反相硅胶柱,用水-甲醇梯度洗脱(10∶0~0∶10),流分经HPLC检验含活性峰的流分再经Sephadex-LH20凝胶柱层析,用甲醇洗脱得化合物1。化合物1经Bruker DRX2500核磁共振仪(500/125 MHz,T MS为内标)和LCQ DECA XP液质联用仪分析,获得化合物1的氢谱、碳谱和分子量。
1.2.5 放线菌菌株SCSI O 1672的16S rDNA系统发育分析 用细菌16S r DNA序列的通用引物,27F primer(5′-AGAGTTTGATC(AC)TGGCTCAG-3′)和1492R primer(5′-ACGG(CT)TACCTTGTTACGACTT-3′),进行PCR扩增。PCR反应体系(20μL):DNA模板0.1μL,10×PCR Buffer 2.0μL,dNTPs(各2.5 mmol/L)1.6μL,27F primer(20μmol/L)0.4μL,1492R pri mer(20 μmol/L)0.4μL,Mg2+(25 mmol/L)1.2μL,Taq酶(5 U/μL)0.2μL,DMSO 1.0μL,ddH2O 13.1 μL。PCR反应条件:94℃预变性4 min,94℃变性1 min,57℃退火1 min,72℃延伸1.5 min;30个循环;72℃延伸10 min;4℃保温。用凝胶回收试剂盒(Axygen)纯化PCR产物,再连接到pGEMT载体(Promega)中,16S r DNA序列由上海英俊生物技术有限公司测定。根据测序结果,利用
BLAST应用软件搜索GenBank、EMBL等数据库
中相关菌株的16S rDNA序列(http://blast.ncbi. nlm.nih.gov/)。利用CLUSTAL X[6]进行多系列比较分析,并使用MEGA4[7]软件构建系统发育树。
2 结 果
2.1 不同培养基发酵产物的产量及活性
20μL不同培养基的发酵产物经HPLC分离后,检测收集的不同吸收峰的海虾毒性,结果表明AM2a、AM2ab培养基的发酵产物tR=10 min的峰活性最好,24 h海虾致死率80%。AM2a、AM2ab 2种培养基发酵产物tR=10 min的峰面积较大,说明AM2a、AM2ab 2种培养基的发酵产物中tR= 10 min的化合物产率比JNP1、JNP2培养基的发酵产物的产率高,见图1。
2.2 结构鉴定
化合物1:UVλmax205,298 nm;ESI-MS:m/z 137(M-H)-,分子量为138.0,结合氢谱、碳谱可以推测其分子式为C7H6O3。1H NMR(500 MHz, DMSO-d6):δH7.77(1H,d,J=7.5 Hz),δH7.45 (1H,m),δH6.89(1H,m),δH6.86(1H,d,J= 7.5 Hz);13C NMR(125 MHz,DMSO-d6):δC171.8(C-1),δC161.3(C-3),δC134.9(C-5),δC130.7(C-7),δC118.6(C-6),δC116.7(C-4),δC114.1(C-2)。化合物的氢谱显示芳环1,2-取代的4个芳香质子信号峰;碳谱显示1个共轭羧基信号峰(δ171.8)、1个连氧芳环碳信号峰(δ 161.3)和另外5个芳环碳信号,结合质谱给出的分子量,分析其结构为水杨酸,见图2。

图1 不同培养基发酵产物的水杨酸含量的HPLC分析Fig.1 HPLC analyses of the fermentation products under four culture conditions
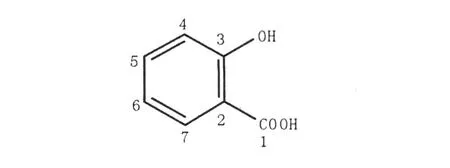
图2 化合物1的结构Fig.2 The chemical structure of compound 1
2.3 放线菌菌株SCSI O 1672的16S rDNA系统发育分析
16S rDNA序列分析表明海洋放线菌SCSI O 1672菌株与链霉菌S.daliensisYI M 31724T和S. rangoonensisLMG 20295T的16S rDNA序列相似性较高,分别为98.3%和97.5%,基于16S rDNA序列的系统发育分析表明海洋放线菌SCSI O 1672菌株与链霉菌S.rangoonensisLMG 20295T为同一分支,支持率为77%(图3),故鉴定为链霉菌Streptom ycessp.SCSI O 1672。

图3 基于16S rDNA序列NJ系统进化树Fig.3 Neighbour-joining tree based on nearly complete 16S rDNA sequences
3 讨 论
水杨酸是生产医药、香料、染料等的重要化工原料。自1943年Strawinski和Stone[9]首先从微生物氧化萘的培养液中分离得到了水杨酸以来,陆续有微生物产水杨酸的报道。比如法幼华等[10]得到了2株产水杨酸较稳定的菌株,鉴定为Pseudom onasovalisAS1.593和Pseudom onas aeruginosaAS1.860;王树青[11]用荧光极毛杆菌(Pseudom onasfluofrscens)和肾炎棒状杆菌(Corynebacterium renals)微生物发酵制取的水杨酸浓度可达20 g/L;Ratledge和W inder[12-13]发现分枝杆菌(M ycobacterium sm egm atis)在缺铁的培养基中能产水杨酸,并对各种离子对水杨酸产量的影响进行了研究;另外Shanmugam和Narayanasamy[14]报道在藓样芽胞杆菌中也产水杨酸; Indiragandhi等[15]发现Acinetobactersp.、Pseudom onassp.和Serratiasp.产水杨酸;Press等[16]报道粘质沙雷菌也产水杨酸;Ratledge和Chaudhry[17]发现Ther m oactinom yces vulgaris菌的发酵液体能分离得到水杨酸。尽管如此,目前还没有发现从链霉菌中分离得到水杨酸的报道。本研究从海洋链霉菌Streptom ycessp.SCSI O 1672的发酵产物中分离得到了水杨酸,丰富了水杨酸的生产菌株库,对该菌株在不同的发酵培养基中水杨酸的产量进行了初步比较研究,发现用AM2a、AM2ab 2种培养基发酵,水杨酸的产量相对较高。
[1]Berdy J.Bioactive microbial metabolites[J].J Antibiot,2005, 58:1-26.
[2]Bull AT,Ward AC,Goodfellow M.Search and discovery strategies for biotechnology:the paradigm shift[J].Microbiol Mol Biol Rev,2000,64(3):573-606.
[3]Feling RH,Buchanan GO,Mincer TJ,et al.Salinosporamide A: a highly cytotoxic proteasome inhibitor from a novel microbial source,a marine bacterium of the new genus Salinospora[J]. Angew Chem Int Ed,2003,42(3):355-357.
[4]Fernández-Chimeno R I,Canedo L,Espliego F,et al.I B-96212, a novel cytotoxic macrolide produced by a marine Micromonospora.I.taxonomy,fermentation,isolation and biological activities[J].J Antibiot,2000,53(5):474-479.
[5]Riedlinger J,Reicke A,Zāhner H,et al.Abyssomicins,inhibitors of the para-aminobenzoic acid pathwayproduced by the marine Verrucosispora Strain AB-18-032[J].J Antibiot,2004,57 (4):271-279.
[6]Thompson JD,Gibson TJ,Plewniak F,et al.The Clustal X windows interface:flexiblestrategies for multiplesequence alignment aided by quality analysis tools[J].Nucleic Acids Res,1997,25(24):4876-4882.
[7]Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular evolutionary genetics analysis(MEGA)software version 4.0[J]. Mol Biol Evol,2007,24(8):1596-1599.
[8]Saitou N,Nei M.The neighbor-joining method:A new method for reconstructing phylogenetic trees[J].Mol Biol Evol,1987,4 (4):406-425.
[9]Strawinski RJ,Stone RW.Conditions governing the oxidation of naphthalene and the chemical analysis of its products[J].J Bacteriol,1943,45:16.
[10]法幼华,徐诗伟,梁家騵,等.微生物氧化萘制取水杨酸[J].微生物学报,1974,14(1):113-111.
[11]王树青.微生物发酵法制取水杨酸[J].科技通报,1988,4 (2):34-36.
[12]Ratledge C,W inder FG.The accumulation of salicylic acid by mycobacteria suring growth on an iron-deficient medium[J]. Biochem J,1962,84:501-506.
[13]Ratledge C,Hall MJ.Influence of metal ions on the formation of mycobactin and salicylic acid in mycobacterium smegmatis grown in static culture[J].J Bacteriol,1971,108(1):314-319.
[14]Shanmugam P,Narayanasamy M.Optimization and production of salicylic acid by rhizobacterial strain Bacilluslichenifor m is MML2501[J].Int J Microbiol,2009,6(1).
[15]Indiragandhi P,Anandham R,Madhaiyan M.Characterization of plant growth-promoting traits of bacteria isolated from larval guts of diamondback moth Plutella xylostella(Lepidoptera:Plutellidae)[J].Curr Microbiol,2008,56:327-333.
[16]Press CM,Wilson M,Tuzun S,et al.Salicylic acid produced by Serratia marcescens90-166 is not the primary determinant of induced systemic resistance in cucumber or tobacco[J].Mol Plant Microbe Interact,1997,10(6):761-768.
[17]Ratledge C,Chaudhry MA.Accumulation of iron-binding phenolic acids by actinomycetales and other organisms related to the mycobacteria[J].J Gen Microbiol,1971,66:71-78.
Isolation and Identification of a Marine Stra in
Streptomyces sp.SCSIO 1672 and Its Metabolite Salicylic Acid
LUO Ming-he1,WANG Zhong-wen2,3,4,HUANG Hong-bo2,3,4,ZHU Qing-hua2,3,4, TIAN Xin-peng2,3,4,BA I Zhi-chuan1,ZHANG Si2,3,4,ZHANG Chang-sheng2,3,4,JU Jian-hua2,3,4
(1.Coll.of Horticult.&Landscape Archit.,SW Uni.,Chongqing400700; 2.Key Lab.of Marine Bio-Res.Sustainable Util.,Guangzhou510301;3.Guangdong Key Lab.of Marine Materia Medica,Guangzhou510301; 4.Ctr.for Marine Microorg.,South China Sea Inst.of Oceanology,Chinese Acad.of Sci.,Guangzhou510301)
A marine actinomycetes strain was isolated and identified as Streptomyces sp.1672 by 16S rDNA sequence analysis.Its fer mentation condition was optimized and its bioactive compound was tested by means of HPLC analysis and bioassay-guided fractionation using brine shrimp toxicity test,and was isolated by solvent extraction,normal and reverse-phase silica gel and other chromatographies.The structure of the bioactive compound was identified as salicylic acid by spectral data analyses.
marine actinomycetes;Streptomyces sp.;salicylic acid
Q939
A
1005-7021(2010)06-0022-05
中国科学院知识创新工程重要方向项目(KSCX2-Y W-G-065,KZCX2-Y W-JC202,KSCX2-Y W-G-073, KZCX2-EZCX2-EWC-G-12);中国科学院南海海洋所领域前沿项目(LYQY200805)
罗明和 男,硕士研究生。主要从事海洋微生物及其活性次级代谢产物研究。
*通讯作者。Tel:020-89023028,E-mail:jju@scsio.ac.cn
2010-08-05;
2010-11-05
猜你喜欢
当代水产(2022年1期)2022-04-26
皮肤病与性病(2021年3期)2021-07-30
国际呼吸杂志(2019年22期)2019-12-09
中国药物应用与监测(2015年5期)2015-12-11
少儿科学周刊·少年版(2015年3期)2015-07-07
少儿科学周刊·少年版(2015年3期)2015-07-07
应用海洋学学报(2014年4期)2014-11-22
应用化工(2014年11期)2014-08-16
天然产物研究与开发(2014年6期)2014-04-27
食品工业科技(2014年15期)2014-03-11
