
GJB2基因条件敲除小鼠耳蜗外毛细胞凋亡时程观察△
2012-01-11 05:41张延平张晓强刘雅莉黄玮孙玉蕊
听力学及言语疾病杂志 2012年4期
张延平 张晓强 刘雅莉 黄玮 孙玉蕊
1 中国人民解放军第309医院耳鼻咽喉科(北京 100091); 2 河南省医学情报研究所; 3 河南省卫生厅
GJB2基因位于染色体13q12,编码缝隙连接蛋白26(Connxin 26, Cx26),是构成细胞间缝隙连接(gap junction)的重要组成部分。Cx26在哺乳动物耳蜗中广泛存在,它所构成的纯合缝隙连接和杂合缝隙连接在维持耳蜗正常生理功能中起重要作用。近年来的研究发现该基因突变是导致人类非综合征型聋(nonsyndromic sensorineural hearing impairment,NSHI)的主要原因之一。目前已经报导的人类GJB2突变已经超过120种,其中绝大多数为功能丧失性突变[1]。为了探讨GJB2突变导致耳聋的机制,研究者先后报道了三种GJB2条件敲除小鼠的动物模型,即Cx26loxP/loxP OtogCre[2], Cx26loxP/ loxP Pax2-Cre和Cx26loxP/loxP foxg1-Cre小鼠[3]。上述三种动物都是耳蜗中特异性缺失Cx26表达,动物出生时内耳发育正常,毛细胞在听觉开始后不久出现死亡,听觉发育成熟后出现耳聋,经检测,凋亡相关蛋白caspase 3在P17时的动物耳蜗细胞中表达阳性,提示凋亡可能是毛细胞死亡的主要原因[2]。上述研究主要关注整个耳蜗的细胞死亡情况,并没有详细探讨外毛细胞的凋亡情况。因此本研究利用异硫氰酸荧光素(fluorescein isothiocyanate, FITC)鳌和的鬼笔环肽(phalloidin )和碘化丙啶(propidium iodide,PI)联合染色技术,研究GJB2基因条件敲除小鼠听觉发育成熟前后不同时期的基底膜毛细胞变化情况,为进一步研究凋亡在敲基因动物耳聋发病中的作用打下基础。
1 材料与方法
1.1 实验动物 GJB2基因条件敲除小鼠cCx26loxp/loxp-Pax2-Cre(cCx26ko)由转基因小鼠Cx26loxp/loxp与 Cx26loxp/--Pax2-Cre小鼠(均由美国Emory大学林曦教授实验室提供)杂交获得,野生型BALB/c小鼠作为正常对照,动物统一编号,统一喂养,双盲法测定,分别选取P8、P14、P18和P21四个发育阶段各6只小鼠进行实验,出生后第3天剪鼠尾提取基因组DNA确定基因型(方法及结果略)。
1.2 实验试剂 磷酸盐缓冲液(PBS,Merk, 德国)、多聚甲醛(Merk, 德国)、Triton X-100(Sigma,美国)、FITC-conjugated phalloidin (Enzo, 法国),PI(Sigma,美国),Fluoromount-GTM(SouthBiotech, 美国),解剖显微镜(Olympus,日本)、激光共聚焦显微镜(LSM 510,Zeiss, Jena,德国) 等。
1.3 耳蜗基底膜铺片 动物用戊巴比妥腹腔注射麻醉后,断头取耳蜗,在解剖显微镜下,用分离针将蜗尖挑开一小孔,挑破圆窗膜,取出镫骨,用细玻璃吸管将4%多聚甲醛由蜗尖小孔灌注,使固定液经前庭阶、鼓阶从前庭窗和圆窗流出,然后将标本放入该固定液里4 ℃冰箱内固定过夜。10% EDTA充分脱钙,在冰PBS中将骨迷路去除,解剖出耳蜗基底膜,分为底、中、顶三段,进行后续染色。
1.4 FITC- phalloidin 和PI染色 耳蜗基底膜经0.25% Triton X-100漂洗5 min, 浸泡于新鲜配置的 FITC- phalloidin 染液中染色30 min ,标记静纤毛和毛细胞表皮板,经PBS漂洗3次后,再用PI (10 μg/ml) 复染 10 min,标记细胞核,再次用PBS漂洗后,抗荧光衰减封片剂 Fluoromount-GTM封片。荧光显微镜下观察毛细胞的变化,激光共聚焦显微镜拍照,每个时间点重复3次。
2 结果
2.1 野生型BALB/c小鼠耳蜗染色结果 荧光显微镜下可见,野生型BALB/c小鼠耳蜗出生后各阶段被绿色异硫氰酸荧光素标记的鬼笔环肽染色的毛细胞肌动蛋白主要分布在毛细胞的纤毛和表皮板。外毛细胞排列整齐,纤毛呈V型排列,表皮板完好,结构正常。被PI染色的毛细胞核大小一致,染色均匀,外形规则,排列有序,无细胞核缺失(图1b)。
2.2 cCx26小鼠耳蜗基底膜染色结果 荧光显微镜下,P8时cCx26小鼠耳蜗形态与相应时期野生型BALB/c小鼠耳蜗形态无明显差异(图1a1)。P14时cCx26小鼠表皮板开始稀松,外毛细胞(outer hair cells, OHC)核排列开始出现不规则,有些细胞有核染色加深现象,但尚未出现明显的表皮板缺损和核形态异常(图1a2 )。在P18时,耳蜗的表面形态在荧光显微镜下出现明显的变化,OHC 核出现固缩、染色浓集、体积变小、核碎裂成片断化,甚至核缺失(图2)。FITC标记的鬼笔环肽染色的毛细胞可见散在性的纤毛结构模糊,表皮板塌陷,将肌动蛋白染色与毛细胞核染色同部位检查,毛细胞核轻微固缩时其表皮板和纤毛结构及染色大致正常,仅在毛细胞核明显固缩时出现表皮板变形和塌陷,提示此时外毛细胞周围的支持细胞,如Dieter细胞,也开始出现了零星的破坏。上述变化在中回最为明显(图2b),其次为底回(图2c),而顶回(图2a)只是出现OHC的形态不规则,并没有明显的核的破坏或消失。在P21时,耳蜗表面结构出现了更为严重的破坏,整个耳蜗都出现了OHC的缺失,中回的破坏最为严重,甚至出现了基底膜的片断化断裂和缺失,提示OHC和支持细胞都出现了缺失 (图1a3)。与野生型小鼠相比, cCx26ko小鼠各阶段内毛细胞纤毛染色不规则,深浅不一,细胞核未见明显形态异常及缺失。
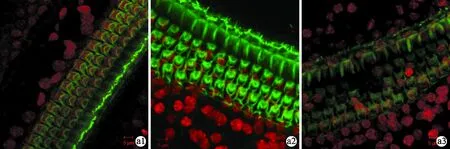
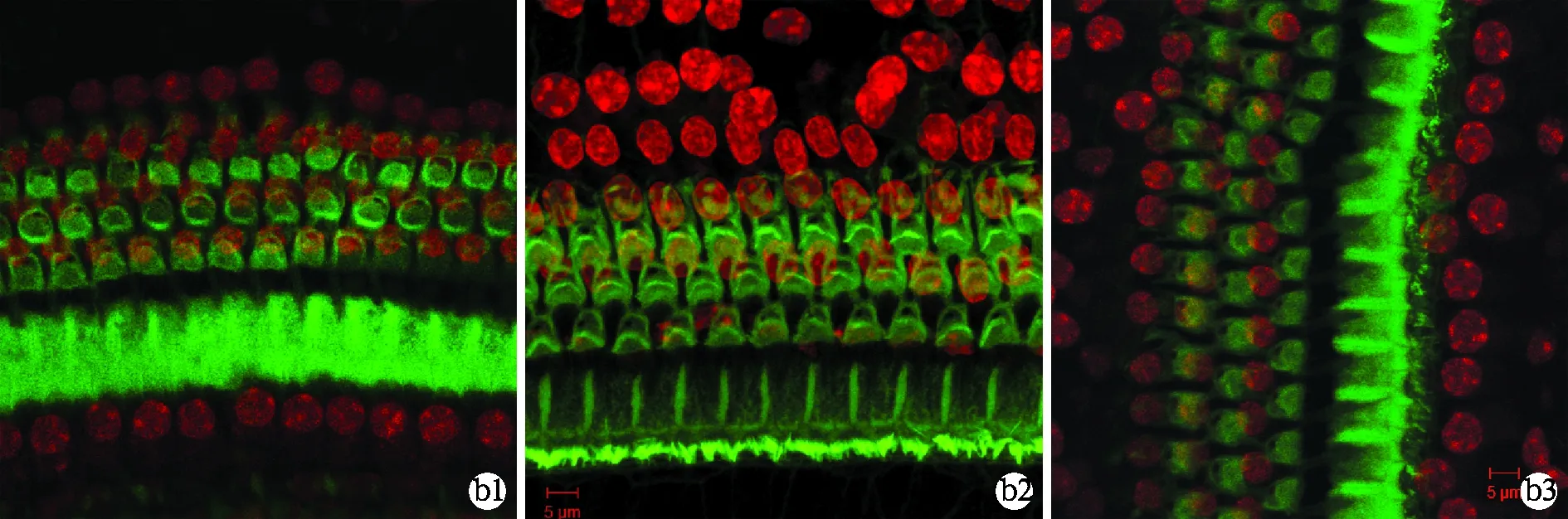
图1 小鼠耳蜗基底膜染色结果(×400)
a1~3为cCx26ko小鼠不同阶段基底膜染色结果,b1~3为野生型小鼠基底膜染色结果。P8时cCx26ko小鼠(a1,中回)与野生型小鼠(b1)相比未见明显差异,P14时cCx26ko表皮板出现疏松、OHCs细胞核深染(a2,中回),但尚无显著细胞缺失。P21时cCx26ko基底膜出现了大范围的缺失,细胞排列紊乱(a3),而野生型小鼠耳蜗基底膜细胞排列连续、整齐(b3)
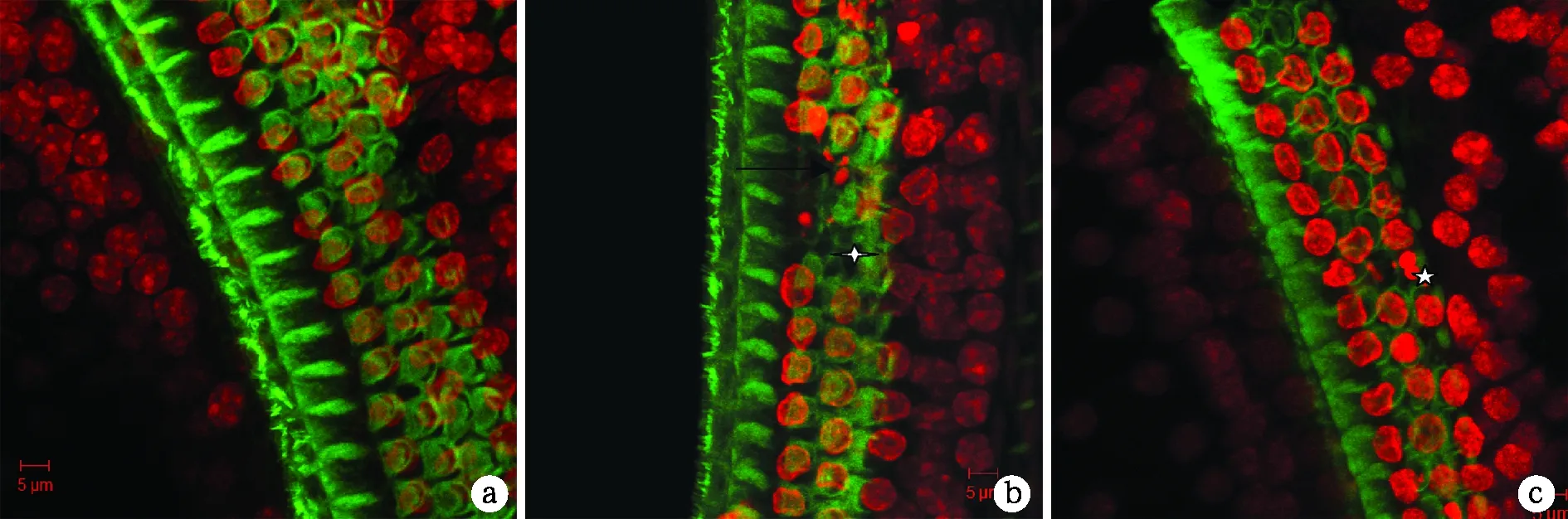
图2 P18cCx26ko小鼠基底膜染色结果(×400)
耳蜗的表面形态在荧光显微镜下出现明显的变化。OHCs 核出现固缩(五角星所示)、染色浓集、体积变小、核碎裂成片断化(箭头所示),甚至核缺失(星号所示)。其中中回(b)较底回(c)和顶回(a)改变明显
3 讨论
全国耳聋分子流行病学调查显示, 我国大多数重度以上感音神经性聋患者仅由为数不多的几个基因突变所致[4,5],其中, 最常见的致聋基因是GJB2 基因, 高达21.01% 的耳聋患者携带该基因突变,GJB2 基因突变在双耳极重度聋组检出率最高,达到 24. 67%[6]。目前GJB2突变导致耳聋的确切机制还不完全清楚。本文通过观察GJB2基因条件敲除小鼠出生后不同阶段的耳蜗PI和鬼笔环肽染色结果,发现在P18时外毛细胞出现了典型的细胞凋亡改变,为进一步研究凋亡在GJB2基因突变导致耳聋的作用机理提供了依据。
基因敲除小鼠是研究基因突变致病机制的良好工具。目前已经有三种GJB2条件敲除小鼠模型报道[2,3],尽管制作动物模型所使用的技术方法不同,但对小鼠耳蜗形态和功能的影响却很一致。三种敲基因动物出生时耳蜗形态与野生型大致相同,提示GJB2基因对于小鼠耳蜗的出生前发育不起重要作用。光学显微镜下观察耳蜗的组织结构显示[1],cCx26ko成年小鼠耳蜗没有明显的膜迷路积水,血管纹细胞也没有明显的缺失,在听觉开始后可见支持细胞和外毛细胞死亡,但内毛细胞形态改变不明显。由于Cre基因的启动子不同,使上述三种小鼠GJB2基因敲除的时间不一致,导致制作的模型小鼠耳蜗在出生后发育中出现一些差异。研究显示[2]Cx26loxP/loxP OtogCre小鼠从出生到P14耳蜗的形态与野生型均无明显差异,从P14开始,Corti器细胞缺失从内毛细胞的支持细胞开始,逐渐蔓延到外毛细胞的支持细胞,然后是外毛细胞以及其他细胞,但内毛细胞和螺旋神经节细胞形态完整,短声ABR测试小鼠反应阈提高约30 dB;此外在P17小鼠耳蜗中检测到caspase 3表达阳性,提示凋亡可能是cCx26ko小鼠耳蜗细胞死亡的主要原因之一。但是在Cx26loxP/loxP;Pax2Cre小鼠,GJB2基因在耳蜗出生后的发育中似乎起到了决定性的作用,光镜下观察到从 P9开始,耳蜗的Corti隧道和Neul氏间隙都未形成,细胞的死亡起始于 Claudius 细胞,然后是外毛细胞和螺旋神经节细胞,小鼠的听力损失也较为严重,短声ABR测试显示几乎所有的频率反应阈均提高到70~90 dB SPL[3]。可见不同胚胎时间敲除GJB2基因,均可导致小鼠耳聋和耳蜗细胞的缺失,但产生的形态学改变和细胞缺失的顺序存在细微的差别。本研究对Cx26loxP/loxP;Pax2Cre小鼠耳蜗扫描电镜观察结果提示外毛细胞缺失发生在P14和P21之间,但cCx26ko小鼠耳蜗毛细胞发生缺失的确切时间还不完全清楚[7]。因此本研究着重观察了外毛细胞缺失与凋亡的关系,同时探讨了外毛细胞缺失的时间,希望为今后进一步研究细胞凋亡在GJB2基因突变致聋机制中的作用提供实验依据。
PI是一种标记死亡细胞DNA的荧光染料,应用PI染色方法,可以对已经死亡但还存在有细胞结构的凋亡和坏死毛细胞进行定量分析。在PI染色的动物耳蜗基底膜中,可以观察到四种类型的毛细胞核,即正常、凋亡、坏死和缺失,毛细胞核固缩和深染色通常被认为是细胞凋亡的典型形态学特征。但因耳蜗基底膜其它种类的细胞核也可以被PI染色,故在荧光显微镜下观察毛细胞核形态学变化时应注意与其它细胞核相区别。鬼笔环肽特异性标记耳蜗毛细胞静纤毛和表皮板的纤丝状肌动蛋白,缺失意味着毛细胞的死亡和溶解。异硫氰酸荧光素标记的鬼笔环肽与PI联合染色可以发现早期凋亡的毛细胞及观察凋亡不同时期细胞纤毛和表皮板与毛细胞核形态学变化的关系[8]。
本文利用FITC标记的鬼笔环肽和PI对野生型和敲基因小鼠耳蜗基底膜进行染色,系统研究了小鼠出生后不同发育时期耳蜗Corti器外毛细胞的缺失方式和缺失时程,结果提示cCx26ko小鼠耳蜗外毛细胞的缺失方式为凋亡,在P18时外毛细胞发生了典型的凋亡改变,耳蜗Corti器的细胞损害在P14时可能已经开始。这一结果为进一步研究cCx26ko小鼠耳蜗细胞凋亡机制提供了基础。
1 Hoang Dinh E, Ahmad S, Chang Q, et al. Diverse deafness mechanisms of connexin mutations revealed by studies using in vitro approaches and mouse models[J]. Brain Res,2009,1 277:52.
2 Cohen-Salmon M, Ott T, Michel V, et al. Targeted ablation of connexin26 in the inner ear epithelial gap junction network causes hearing impairment and cell death[J]. Curr Biol,2002,12:1 106.
3 Wang Y, Chang Q, Tang W, et al. Targeted connexin26 ablation arrests postnatal development of the organ of Corti[J]. Biochem Biophys Res Commun,2009,385:33.
4 于飞, 韩东一, 戴朴, 等. 1190 例非综合征性耳聋患者GJB2 基因突变序列分析[J]. 中华医学杂志, 2007,87:2 814.
5 Dai P, You Y, Cui J, et al. GJB2 mutat ion spectrum in deaf population in a typical southeastern area o f China[J]. J Otol, 2006,1:94.
6 王国建, 袁永一, 李荣,等. 不同听力学表型人群中常见耳聋基因突变检出率的分析[J].临床耳鼻咽喉头颈外科杂志,2011,25:445.
7 张延平,佟明望, 李丽娜,等.GJB2基因条件敲除小鼠耳蜗扫描电镜观察[J]. 山东大学学报(医学版),2010,48:9.
8 杨卫平,胡博华.耳蜗毛细胞凋亡早期的检测方法[J].军医进修学院学报,2005,26:122.
猜你喜欢
听力学及言语疾病杂志(2022年5期)2022-09-20
基础医学与临床(2020年6期)2020-02-12
中华耳科学杂志(2020年6期)2020-01-08
特种经济动植物(2019年9期)2019-01-08
数码世界(2018年1期)2018-12-23
振动与冲击(2018年4期)2018-03-05
中华耳科学杂志(2018年6期)2018-01-16
振动与冲击(2017年14期)2017-07-19
天然产物研究与开发(2016年6期)2016-06-05
中华耳科学杂志(2015年3期)2015-12-01
