
甘蓝无蜡粉突变体叶表皮蜡质超微结构观察
2013-02-24 09:01牟香丽
中国蔬菜 2013年4期
牟香丽 王 超 王 帅
(东北农业大学园艺学院,黑龙江哈尔滨 150030)
植物能够适应外界环境的变化并能生存下来的一个重要机制是能够分泌蜡质到角质层表面或者角质层内,充当阻挡紫外辐射及病虫侵害的第一道屏障(Kunst & Samuels,2003)。蜡质存在于植物的木栓层、受伤的组织(Hamiton,1995)、花粉、种皮等组织器官中(Preuss et al.,1993)。近年来,与植物表皮蜡质相关的研究越来越多,已经从小麦(Muria & Taira,1999)、拟南芥(Broun et al.,2004)、水稻(周小云,2006)、小盐芥(赵宝,2007)等植物中克隆了一些与蜡质相关的基因,这为阐明蜡质组分的生理生化作用及其分子机制打下了基础。
结球甘蓝(Brassica oleraceaL.var.capitataL.)是世界上许多国家的主要蔬菜作物,其表面都披有不同程度的蜡粉霜,叶色呈粉蓝色或蓝绿色;而无蜡粉的亮叶型结球甘蓝突变体整个株体表面无蜡粉覆盖,颜色油亮,叶脉呈黄绿色或绿色,整株光泽度和绿色度都要高于野生型。经过多年的选培工作,该突变性状已能稳定遗传,为了区别原来的普通甘蓝,称此类型甘蓝为亮叶甘蓝。据初莲香和王余文(1993)报道,亮叶甘蓝质地脆嫩,叶片薄,有光泽,富含糖、VC 及干物质等营养成分。亮叶甘蓝还具有一定的抗虫性,Stoner(1990)研究认为,在无蜡粉甘蓝上,甘蓝蚜的种群数量比正常甘蓝植株低1/10,小菜蛾幼虫的数量是正常植株的1/10;Lin等(1984)发现无蜡粉甘蓝能影响初孵小菜蛾的产卵、取食等行为,而且1 龄幼虫在无蜡粉甘蓝上的成活率不到正常植株的10%。因此,对于甘蓝无蜡粉突变体的研究具有十分重要的意义。
关于植物叶表蜡粉突变体的报道甚多,蜡质基因一直被研究者所关注,但对蜡质突变体超显微水平的研究甚少,实际上对蜡质突变体超微结构进行观察对于了解蜡质突变情况及探索蜡质发育意义重大。本试验以突变型亮叶甘蓝和野生型结球甘蓝为试材,对甘蓝叶片发育过程中表皮最外层蜡质发育及突变情况进行系统地观察,以期在超显微水平上了解甘蓝无蜡粉突变体。
1 材料与方法
1.1 材料
供试材料为蜡质缺失突变型甘蓝材料亮叶98-1030 和蜡质正常的野生型甘蓝材料98-1030,均由东北农业大学园艺学院甘蓝课题组提供和保存。98-1030 为早熟甘蓝品种迎春自交得到,早熟,圆球形,蜡粉正常;2007年,在以98-1030 为试验材料进行组织培养试验时,发现无蜡粉突变体,颜色油亮,无蜡粉,并命名为亮叶98-1030。
试验于2012年春在本院园艺站和农场进行。将98-1030 和亮叶98-1030 播种于园艺站温室,并于四叶一心时移栽于农场大田。
1.2 方法
自甘蓝幼苗出土开始,在一叶苗期、三叶苗期、结球前期、结球后期及成熟期分别取样。在苗期选取新生幼叶为材料,在结球期和成熟期选取最内层外叶为材料。
扫描电镜样品制作:分别将新鲜叶片冲洗干净,避开叶脉切成5 mm×10 mm 的长方形小块,写好编号,四氧化锇熏蒸48 h,晾干3 d,E-1010(HITACHI)型离子溅射镀膜仪镀金膜,置于S-3400N(Hitachi)型扫描电子显微镜下进行观察。
2 结果与分析
2.1 一叶苗期甘蓝叶表皮蜡质超微结构
甘蓝种子出苗后,第1片真叶刚刚展开时,从外观上看,甘蓝蜡质突变型与野生型的子叶光滑度和绿色度没有明显区别;第1片真叶在绿色度和光滑度上也差别不显著。
分别将突变型和野生型的甘蓝幼苗子叶和真叶置于扫描电子显微镜下。结果表明(图1):两者的子叶表皮蜡质差异极显著,突变型叶片表皮有类似瘤状突起,处于蜡质发育的初始阶段,未形成独立的蜡质颗粒;而野生型叶片表皮蜡质数量明显多于突变型,蜡质晶体结构为片状或聚集成团。两者的第1片真叶表皮蜡质差异显著,突变型叶片表皮还未见蜡质发育;而野生型已有逐渐发育完全的蜡质晶体,发育进程由颗粒状到片状再到不规则三维结构。
2.2 三叶苗期甘蓝叶表皮蜡质超微结构
三叶苗期的甘蓝叶片已经完全暴露到外界环境中,表观上突变型与野生型甘蓝幼苗叶片绿色度和光滑度已有差异,突变型叶片光滑且亮绿;野生型叶片覆有蜡粉霜。
电镜观察结果表明(图2),突变型叶片表皮的背腹面都发现有蜡质晶体存在,但是发育不健全,以颗粒状为主,即使片状也比野生型的蜡质晶体小;野生型蜡质晶体发育成熟,以片状、针状为主。
2.3 结球前期甘蓝叶表皮蜡质超微结构
结球前期的甘蓝根、茎、叶都处于快速生长时期。表观上突变型与野生型叶片绿色度和光滑度差异极为显著,突变体叶片光滑,颜色亮绿;野生型叶片粉蓝色,覆有蜡粉霜。
电镜观察结果(图3),突变型与野生型叶片背腹面蜡质晶体积累都进一步增加。突变型的蜡质晶体仍然是颗粒状,且其数量腹面比背面多;野生型的蜡质晶体排列更加密集,腹背面出现不同的晶体结构,腹面主要是圆柱状,背面主要是片状。

图1 一叶苗期甘蓝叶表皮蜡质晶体扫描电镜观察结果(×3000,×2000)
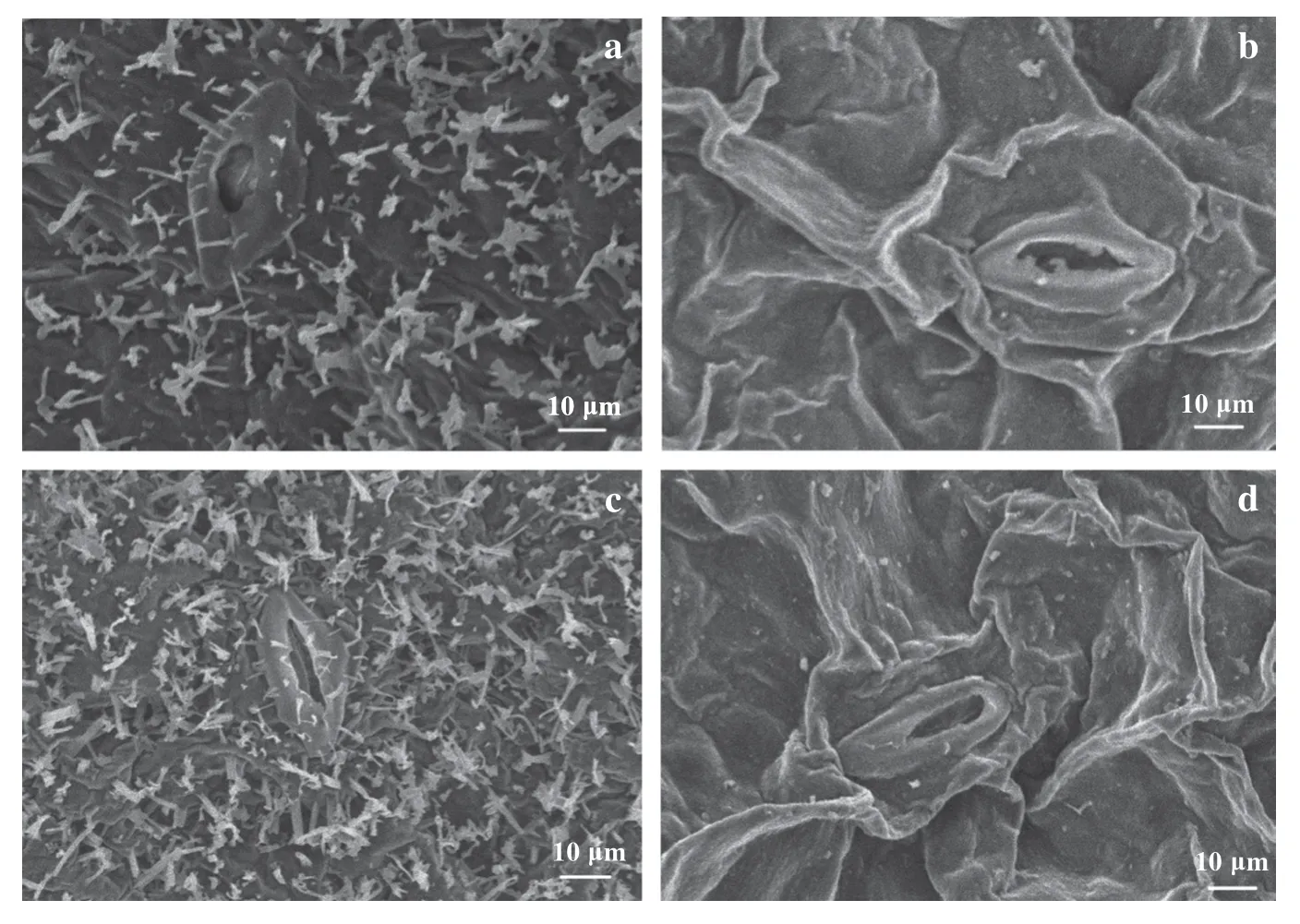
图2 三叶苗期甘蓝叶表皮蜡质晶体扫描电镜观察结果(×3000)
2.4 结球后期甘蓝叶表皮蜡质超微结构
结球后期的甘蓝根、茎、叶处于生长旺盛时期,各器官已经趋于成熟。表观上突变型甘蓝的外叶和球叶呈现亮绿色;而野生型甘蓝的外叶呈粉蓝色,球叶呈粉绿色,都覆有蜡粉霜。电镜观察此期突变型叶片腹面蜡质晶体为颗粒状,背面蜡质晶体不明显;野生型叶片背腹面蜡质晶体密集,以片状为主(图4)。

图3 结球前期甘蓝叶表皮蜡质晶体扫描电镜观察结果(×3000)

图4 结球后期甘蓝叶表皮蜡质晶体扫描电镜观察结果(×3000)
2.5 成熟期甘蓝叶表皮蜡质超微结构
甘蓝成熟期,叶球的外叶数、质量、体积和开展度基本上停止变化,外叶的功能进一步退化。纵切突变型与野生型甘蓝叶球,由外到内颜色均由绿色到黄绿再到浅黄过度,其心叶颜色无明显区别。但外叶颜色差别显著,突变型的外叶颜色亮绿,野生型外叶粉蓝色且覆有蜡粉霜。电镜观察突变型叶表蜡质晶体仍然是以颗粒状为主;野生型蜡质晶体由棒状转化成针状或片状,片状晶体的末端分散,有蜡质晶体进一步崩溃的迹象(图5)。

图5 成熟期甘蓝叶表皮蜡质晶体扫描电镜观察结果(×3000)
3 结论与讨论
植物叶片表皮最外层蜡质晶体结构一直是研究的热点,Koch 和Ensikat(2008)认为植物叶片表皮蜡质晶体为不规则的三维结构,垂直分布在叶片表面;Barthlott 等(1998)将植物表皮蜡质的微观形态结构分为柱状、棒条状、垂直片状、线状、烟囱状等26 类,其中片状和管状是蜡质晶体结构中最主要的类型。本试验中,野生型甘蓝叶片表皮蜡质晶体垂直分布在叶片表面,且不同生长时期表现出不同的蜡质晶体结构,苗期以颗粒状、片状和针状为主,结球期以圆柱状和片状为主,成熟期以片状和针状为主,片状晶体结构是甘蓝整个生长期的最主要类型。电镜观察蜡质发育过程发现,当蜡质晶体积累逐渐增多时,晶体镶嵌排列更加紧密,电镜下观察不易发现气孔结构。可以推测,蜡质晶体密布成网覆盖在叶片表面,作为叶表天然的保护层,使甘蓝在生长中期和后期有更强的抗逆性。本试验观察到甘蓝叶表皮腹面蜡质数量要多于背面,这可能与叶片腹—背轴非对称发育有关,此观察结果也与周小云(2006)、曾爱松等(2009)的研究结果相似。
另外,本试验通过电镜观察发现突变型甘蓝材料蜡质缺失极显著,与李景涛等(2012)的研究结果一致。但本试验发现突变型甘蓝苗期子叶和真叶有颗粒状和发育不完全的片状蜡质晶体,而且整个生长期以颗粒状晶体结构为主。可以推测,即使表观无蜡粉的甘蓝突变体实际上也有蜡质晶体发育萌芽,可能是某种原因阻碍了蜡质晶体的进一步发育,究其是阻断了蜡质合成途径还是阻碍了蜡质运输途径,或是其他原因,还需要进一步探索。
关于甘蓝叶片表皮蜡质突变体已多有报道,但研究还不够深入。今后,如果能从分子水平去研究突变基因的功能,成功克隆一些相关基因,再通过基因工程手段改良角质层性状,从而提高甘蓝的抗旱及抗病能力,这在育种上意义会更大。
初莲香,王余文.1993.无蜡粉亮叶结球甘蓝的发现及其利用.辽宁农业科学,(3):49-50.
李景涛,杨丽梅,方智远,刘玉梅,庄木,张扬勇,孙培田.2012.结球甘蓝10Q-961 无蜡粉亮绿性状遗传规律初探.中国蔬菜,(12):37-41.
曾爱松,刘玉梅,方智远.2009.甘蓝耐裂球性与叶表面微形态及细胞组织结构的关系研究.华北农学报,(2):41-45.
赵宝.2007.三种生态型盐芥(Thelungiella halophila)表皮蜡质及其合成相关的KCS 家族各基因表达分析研究〔硕士论文〕.济南:山东师范大学.
周小云.2006.水稻叶表皮蜡质发育及蜡质相关转录因子基因OsWTF1和OsWTF2的克隆与鉴定〔博士论文〕.长沙:湖南农业大学.
Barthlott W,Neinhuis C,Cutler D,Ditseh F,Meusel I,Wilhelmi H.1998.Classification and terminology of plant epieutieular waxes.Botanical Journal of the Linnean Society,126:237-260.
Broun P,Poindexter P,Osborne E,Jiang C Z,Riechmann J L.2004.WIN1,a transcriptional activator of epidermal wax accumulation in Arabidopsis.Proceedings of the National Academy of Sciences of the USA,101(13):4706-4711.
Hamiton R J.1995.Waxes:chemistry,molecular biology and functions.Physiologia Plantarum,115:89-93.
Koch K,Ensikat H J.2008.The hydrophobic coatings of plant surfaces:epicuticular wax crystals and their morphologies,crystallinity and molecular self-assembly.Micron,39(7):759-772.
Kunst L,Samuels A L.2003.Biosynthesis and secretion of plant cuticular wax.Progress in Lipid Research,42(1):51-80.
Lin J,Dickson M H,Eckenrode C J.1984.Resistance ofBrassicalines to the diamondback moth(Lepidoptera:Yponomeutidae)in the field and inheritance resistance.Journal of Economic Entomology,77:1293-1296.
Muria J,Taira T.1999.Isolation and characterization of the threeWaxygenes encoding the granule-bound starch synthase in hexaploid wheat.Gene,234:71-79.
Preuss D,Lemieux B,Yen G,Davis R W.1993.A conditional sterile mutation eliminates surface components fromArabidopsispollen and disrupts cell signaling during fertilization.Gene,7:974-985.
Stoner K A.1990.Glossy leaf wax and plant resistance to insects inBrassica oleraceaunder natural infestation.Environmental Entomology,19(3):730-739.
猜你喜欢
食品科学(2022年23期)2022-12-30
辽宁农业科学(2021年6期)2021-12-02
上海农业科技(2020年5期)2020-10-24
上海蔬菜(2020年5期)2020-10-20
中国病理生理杂志(2018年9期)2018-09-27
麦类作物学报(2018年8期)2018-08-28
浙江农业科学(2017年4期)2017-05-11
癌症进展(2015年5期)2015-07-26
中国现代医生(2015年5期)2015-03-31
中国实验诊断学(2014年4期)2014-08-25
