
植物数量性状“主基因+多基因”混合遗传模型及其在烟草上的应用
2013-03-21 01:29张兴伟王志德刘艳华任民杨洋
中国烟草学报 2013年3期
张兴伟,王志德,刘艳华,任民,杨洋
1 农业部烟草生物学与加工重点实验室,中国农业科学院烟草研究所,青岛 266101;
2 丹东市烟草专卖局(公司)宽甸分公司,丹东 118222
烟草育种所注重的若干性状,一般都是数量性状(quantitative character)。数量性状由多个基因控制,每个基因控制性状表现的一部分,难以根据表型的差异检测出具体基因。经典数量遗传学一般只能从一组基因的总体上得到有关遗传效应及其相对重要的信息。而对植物遗传改良来说,需要有关数量性状遗传体系以及其中具体基因的重要信息,以便通过育种手段进行遗传操作。植物数量性状“主基因+多基因”混合遗传模型的出现,能有效地解决上述问题。“主基因+多基因”混合遗传模型是植物数量性状的通用模型,盖钧镒等[1]在此基础上发展了一套适合植物遗传分析的分离分析方法。该方法适于育种工作者利用杂种分离世代的数据对育种性状的遗传组成做出判断,制订相应的育种策略,也可用以校验QTL定位所揭示的性状遗传组成。
1 植物数量性状“主基因+多基因”混合遗传模型的主要环节
1.1 分离世代的理论分布
在二倍体核遗传、不存在母体效应、主基因和多基因无互作和连锁、配子和合子均无选择的假定下,每一主基因型由于多基因和环境的修饰呈正态分布,整个分离世代的表型分布为多个主基因型正态成分分布的混合分布;不分离世代如P1、P2、F1等的表型分布为正态分布。
1.2 遗传模型
目前,植物数量性状“主基因+多基因”混合遗传模型主要包括以下几种模型:一对主基因(A)、两对主基因(B)、多基因(C)、一对主基因+多基因(D)、两对主基因+多基因(E)、三对主基因(F)、三对主基因+多基因(G)。
1.3 试验群体类型
单世代分析,包括P1、P2、DH或RIL群体3个世代的联合分析,P1、P2、F1、F24个世代的联合分析,P1、P2、F1、F2、F2:35 个世代的联合分析,P1、P2、F1、F2、B1、B26 个世代联合分析以及 P1、P2、F1、F2:3、B1:2、B2:26个家系世代试验联合分析。
1.4 ECM算法及对数极大似然函数
按照试验群体类型建立各种模型的混合分布函数,并建立各模型的对数极大似然函数。由各试验群体及世代的观察次数分布通过IECM(iterated expectation and conditional maximization)[2]算法对各模型的对数极大似然函数估计出相应的各世代、各成分分布的参数,以及相应的似然函数值。
1.5 筛选最优模型及估算遗传参数
方法是由模型的似然函数值计算出AIC(Akaike’s information criterion)[3]值,AIC值最小的模型为最佳模型,模型间AIC值差异不大时要进行适合性测验,这组测验包括均匀性U12、U22及检验,nW2检验和Dn检验,同时选得各成分分布参数。采用最小二乘法由最优遗传模型的各成分分布参数计算出主基因和多基因的相关遗传参数。
2 植物数量性状“主基因+多基因”混合遗传模型在烟草中的应用(详见表1)
2.1 在植物学性状上的应用
王日新等[4]对株高研究结果表明,其遗传符合2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型(E1),主基因遗传率为71.60%-88.55%。张兴伟等[5]对株高研究结果表明,其遗传符合2对加性-显性-上位性主基因+加行-显性-上位性多基因混合遗传模型(E0),主基因遗传率为57.53%。张兴伟等[6]对叶数、叶面积的研究结果表明,其遗传也符合E0模型,主基因遗传率分别为42.63%和30.32%。张兴伟等[5]同时对其它几个植物学性状的研究表明,鲜叶重遗传也符合E0模型,主基因遗传率为44.26%;移栽至中心花开放天数遗传符合E1模型,主基因遗传率为64.79%;茎围和比叶重均受1对完全显性主基因+加性-显性多基因(D3)控制,遗传以多基因为主,其多基因加性效应和显性效应大小相当,比叶重遗传主基因、多基因的加性效应和显性效应大致相当,主基因遗传率分别为2.48%和38.71%。叶形指数受1对加性-显性主基因+加性-显性-上位性多基因(D0)控制,主基因加性效应与显性效应基本相当,主基因遗传率为49.64%。叶长、叶宽、节距和蒴果重受加性-显性-上位性多基因(C0)控制,多基因遗传率分别为60.75%、62.14%、75.08%和82.34%。
2.2 在抗病上的应用
蔡长春等[7]对黑胫病的研究结果表明,黑胫病发病率是由2对具有互补作用的主基因+多基因(E-1-7)控制,主基因遗传率为54.14%。高加明等[8]对青枯病的研究结果表明,其遗传符合E1模型,基因效应以上位性效应为主,主基因遗传率为49.63%。牟建英等[9]对白粉病的研究结果表明,其遗传符合E0模型,以显性效应及上位性效应为主,主基因遗传率为84.43%。
2.3 在化学成分上的应用
蔡长春等[10]对斩株前和晾制后白肋烟的烟碱含量研究结果表明,其遗传受2对主基因控制,同时存在多基因的修饰作用,前者为累加作用(E-1-6),后者为互补作用(E-1-7),主基因遗传率分别为54.88%和45.63%。尹天水等[11]对钾含量的研究结果表明,其遗传符合D0模型,主基因遗传率为52.78%。
2.4 在生理性状上的应用
张兴伟等[12]对叶绿素含量研究结果表明,其遗传符合1对加性-显性主基因+加性-显性-上位性多基因混合遗传模型(D0),主基因遗传率为48.94%。张兴伟等[13]对光合性状研究结果表明,光合速率和蒸腾速率的遗传符合C0模型,多基因遗传率分别为20.69%和13.56%,气孔导度遗传符合E0模型,以上位性效应为主,主基因遗传率为40.59%。
2.5 在烘烤性状上的应用
倪超等[14]、王英俊等[15]对易烤性研究结果表明,其遗传符合2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型(E1),主基因遗传率分别为64.09%、65.61%。郝贤伟等[16]对耐烤性研究结果表明,其遗传符合E0模型,以加性效应为主,主基因遗传率为56.81%-79.61%。
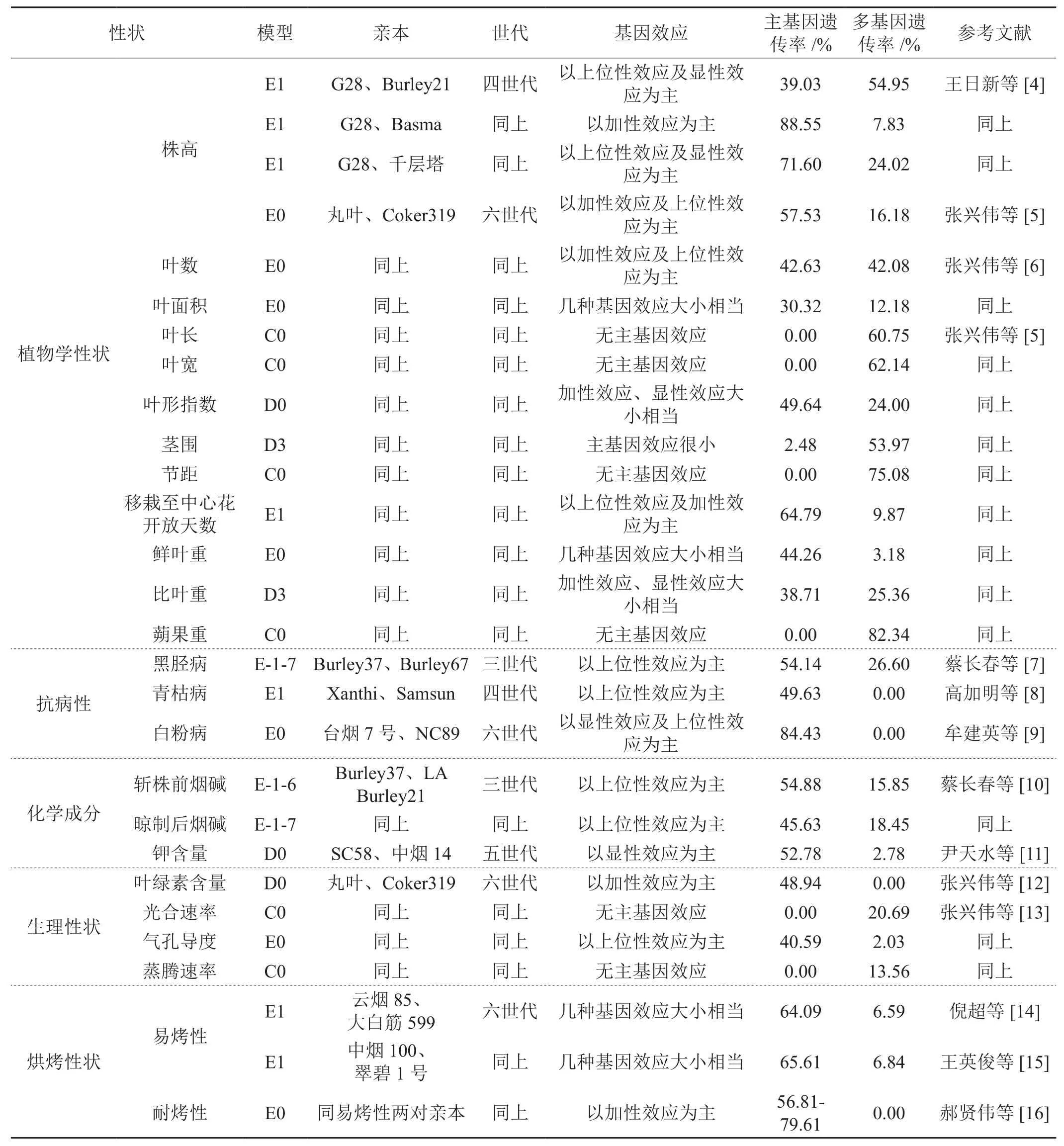
表1 近7年来对栽培烟草重要性状的遗传分析
植物数量性状“主基因+多基因”混合遗传模型在烟草上应用以来,在烟草植物学性状、抗病性状、化学成分、生理性状及烘烤性状等方面,对于揭示相关性状的遗传组成及遗传效应发挥了积极的效用,该方法适于育种工作者利用杂种分离世代的数据对育种性状的遗传组成做出初步判断,制订相应的育种策略,也可用以校验QTL定位所揭示的性状遗传组成。同时该模型也需要不断拓展,以满足复杂数量性状分析的客观要求。
3 关于“主基因+多基因”混合遗传模型的几点思考
“主基因+多基因”混合遗传模型是植物数量性状的通用模型,盖钧镒等[1]在此基础上发展了一套适合植物遗传分析的1对主基因+多基因、2对主基因+多基因和3对主基因+多基因混合模型的分离分析方法。此方法自提出以来已广泛应用于水稻[17]、棉花[17]、油菜[19]、小麦[20]、大豆[21]等多种粮食和经济作物,而在烟草上的应用研究[4-16],这几年也逐渐多了起来。在实际应用中,有些研究者拿来数据直接套用模型,存在不少误区,下面是笔者对于“主基因+多基因”混合遗传模型的几点思考。
3.1 极端亲本材料的选择
亲本一般要选择关于某性状的极端材料(如高抗和高感),这样有助于消除中间材料的干扰和误差,有利于研究该性状真实的遗传规律。
3.2 试验设计
一般要进行正反交和随机区组设计,三次重复。正反交结果可确定是细胞核效应还是质核互作效应;随机区组设计重复之间和处理之间差异是否显著,可说明2个亲本某性状在遗传方面是否存在着真实稳定的差异,进而决定是否可以作为研究该性状遗传规律的试材进行分析。
3.3 数据分析
获取数据后,首先要看不分离群体(P1、P2和F1)的数据是否符合正态分布,SAS可做正态分布分析,最简单的方法画个柱形图看看;若不符合,剔除明显变异的数据,再进行下一步分析;根据AIC准则选择模型时,一般选择3-5个作为备选模型,具体选择多少,要视具体情况而定,若已选模型的AIC值与其它相比,差异较大,则少选几个,反之,则多选几个。
3.4 多年多点多时期数据的精准测定
对某一性状最好进行多年多点多时期的精准测定,这样可研究某一性状数量基因的时空表达规律。
3.5 “主基因+多基因”混合遗传模型基因对数的拓展
育种实践和QTL定位的结果表明,很多数量性状的遗传受到4对甚至更多的主基因控制。章元明等[22]构建的遗传模型最多只包含3对主基因,已不能满足育种研究的需要,因此需要拓展遗传模型,目前王金社等[23]已经把重组自交系群体初步拓展到4对主基因,但更多遗传群体的基因对数仍需拓展。
3.6 “主基因+多基因”混合遗传模型在烟草上的优先应用策略
近几年,随着烟草基因组计划研究的深入,烟草诸多性状的QTL定位成了研究热点。笔者建议,在进行QTL分析前,可采用“主基因+多基因”混合遗传模型对相应性状进行优先分析。这样既可以在早期对育种性状的遗传组成做出初步判断,制订相应的育种策略,也可用以校验QTL定位所揭示的性状遗传组成。
[1] 盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[2] 章元明,盖钧镒. 数量性状分离分析中分布参数估计的IECM算法[J]. 作物学报,2000,26(6):699-706.
[3] Akaike H. On entropy maximum principle [M]// Krishnaiah P R. Applications of Statistics. Amsterdam: NorthHolland Publishing Res, 1977:27-41.
[4] 王日新,任民,张兴伟,等. 普通烟草栽培种内株高性状主基因加多基因遗传分析[J]. 中国烟草科学,2009,30(2):15-20.
[5] 张兴伟,王志德,任民,等. 烤烟几个植物学性状的遗传分析[J]. 中国烟草科学,2012,33(5):1-8.
[6] 张兴伟,王志德,孙玉合,等. 烤烟叶数、叶面积的遗传分析[J]. 植物遗传资源学报,2012,13(3):467-472.
[7] 蔡长春,张俊杰,黄文昌,等. 利用DH群体分析白肋烟黑胫病抗性的遗传规律[J]. 烟草科技,2009(1):54-59,63.
[8] 高加明,王志德,张兴伟,等. 香料烟青枯病抗性基因的遗传分析[J]. 中国烟草科学,2010,31(1):1-4.
[9] 牟建英,钱玉梅,张兴伟,等. 烟草白粉病抗性基因的遗传分析[J]. 植物遗传资源学报,2013,14(4).
[10] 蔡长春,张俊杰,黄文昌,等. 利用DH群体分析白肋烟烟碱含量的遗传规律[J]. 中国烟草学报,2009,15(4):55-60.
[11] 尹天水,王树会,石磊. 烤烟烟叶钾含量的遗传分析[J].烟草科技,2005(5):34-38.
[12] 张兴伟,王志德,牟建民,等. 烤烟叶绿素含量遗传分析[J].中国烟草学报,2011,17(3):48-52.
[13] 张兴伟. 烟草微核心种质构建及相关性状数量遗传分析[D]. 中国农业科学院研究生院博士学位论文,2013,10-11.
[14] 倪超,徐秀红,张兴伟,等. 烤烟品种易烤性相关性状的主基因+多基因遗传分析[J]. 中国烟草科学,2011,32(1):1-4, 11.
[15] 王英俊,倪超,翟明泉,等. 烤烟易烤性遗传分析[J]. 作物研究,2011,25(1):42-46.
[16] 郝贤伟,徐秀红,许家来,等. 烤烟品种耐烤性遗传分析[J].中国农业科学,2012,45(23):4939-4946.
[17] 朱世杨,郭媛,洪德林.水稻种子抗老化遗传分析[J].遗传,2008,30(2):217-224.
[18] 袁有禄,张天真,郭旺珍,等. 棉花高品质纤维性状的主基因与多基因遗传分析[J]. 遗传学报,2002,29(9):827-834.
[19] 张书芬,傅廷栋,朱家成,等. 甘蓝型油菜芥酸含量的基因分析[J]. 中国农业科学,2008,41(10):3343-3349.
[20] 张立平,赵昌平,单福华,等. 小麦光温敏雄性不育系BS210育性的主基因+多基因混合遗传分析[J].作物学报,2007,33(9):1553-1557.
[21] 王春娥,盖钧镒,傅三雄,等. 大豆豆腐和豆乳得率的遗传分析与QTL定位[J]. 中国农业科学,2008,41(5):1274-1282.
[22] 章元明,盖钧镒,王永军. 利用P_1、P_2和DH或RIL群体联合分离分析的拓展[J]. 遗传,2001,23(5):467-470.
[23] 王金社,李海旺,赵团结,等. 重组自交家系群体4对主基因加多基因混合遗传模型分离分析方法的建立[J].作物学报,2010,36(2):191-201.
猜你喜欢
临床肝胆病杂志(2020年1期)2020-12-20
聊城大学学报(自然科学版)(2020年1期)2020-11-27
科学导报(2020年51期)2020-09-09
消费导刊(2017年24期)2018-01-31
中国科技纵横(2016年15期)2016-12-29
甘肃教育(2016年22期)2016-12-20
中国烟草科学(2012年5期)2012-07-31
现代电子技术(2009年9期)2009-06-25
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
