
溶氧水平对鳊鱼、中华倒刺鲃幼鱼游泳能力的影响
2013-04-19 08:50赵文文曹振东付世建
水生生物学报 2013年2期
赵文文 曹振东 付世建
(重庆师范大学进化生理与行为学实验室, 重庆市动物生物学重点实验室, 重庆 401331)
溶氧水平对鳊鱼、中华倒刺鲃幼鱼游泳能力的影响
赵文文 曹振东 付世建
(重庆师范大学进化生理与行为学实验室, 重庆市动物生物学重点实验室, 重庆 401331)
在(25 ± 1)℃条件下, 以鳊鱼(Parabramis pekinensis)[体重(4.70 ± 0.11)g,n= 32]、中华倒刺鲃(Spinibarbus sinensis)[体重(3.26 ± 0.06)g,n= 32]幼鱼为研究对象, 采用鱼类游泳代谢测定仪在水体溶氧为8、4、2、1 mgO2/L条件下分别测定其临界游泳速度(Ucrit)和游泳代谢率(MO2), 并计算出静止代谢率(MO2rest)、最大游泳代谢率(MO2max)、代谢范围(MS)及单位位移能耗(COT)等相关参数。结果显示, 随着溶氧水平的下降, 鳊鱼、中华倒刺鲃幼鱼的Ucrit均逐渐下降, 除中华倒刺鲃幼鱼的值在4与8 mgO2/L下没有显著性差异外, 其他各组均差异显著(P<0.05); 在同一溶氧水平下的中华倒刺鲃Ucrit显著大于鳊鱼(P<0.05)。两种鱼的MO2max和MS均随DO的下降而显著下降, 但MO2rest在溶氧水平低于1 mgO2/L才显著下降(P<0.05)。研究还发现鳊鱼、中华倒刺鲃幼鱼的MO2在同一游泳速度下随溶氧水平下降而降低, 而在相同溶氧水平下随游泳速度的上升而显著升高(P<0.05);COT随游泳速度上升而显著降低(P<0.05), 但在高游泳速度下相对稳定, 在同一游泳速度下随着DO的下降有所减小。中华倒刺鲃的COT整体上小于鳊鱼, 且在低游泳速度下差异更大。87.5%(1—8 mgO2/L)溶氧水平的下降导致两种鱼类相似的Ucrit变化(53%vs.50%), 但溶氧水平由8降到4 mgO2/L时, 鳊鱼32%MO2max的下降导致Ucrit下降13%, 但同样的溶氧水平下降虽然导致中华倒刺鲃的MO2max下降20%,但由于MO2rest和COT的下降, 其Ucrit并没有显著的变化。实验结果表明: 不同溶氧水平对不同鱼种游泳能力的影响存在差异, 这种差异与其代谢对策密切相关。
鳊鱼; 中华倒刺鲃; 溶氧水平; 临界游泳速度; 游泳能力; 代谢率; 适应对策
鱼类种类繁多、生活史复杂, 生理活动受到各种环境因子的影响[1]。由于水体环境较陆生环境氧气含量低、空间和时间变化大, 如何应对环境低氧胁迫成为鱼类的首要生理问题之一。因此, 在环境对鱼类生理生态影响的相关研究中, 溶氧因子的作用备受关注[2—5]。游泳运动是鱼类逃逸、捕食、繁殖等行为实现的主要方式, 为其生存、生长及繁衍等提供了基本保证[6,7]。鱼类的游泳行为伴随着大量的能量消耗, 游泳时的能量代谢适应对策对于鱼类生存繁衍同样至关重要[8]。临界游泳速度(Critical swimming speed, Ucrit)是衡量鱼类有氧运动能力的重要指标, 与觅食、洄游等生理活动联系密切[9], 相关研究有着重要的理论意义[9]。
环境溶氧水平的骤然变化对鱼类的游泳能力的影响研究发现不同的鱼类由于生理生化的差异对水体溶氧变化敏感性不同[11—14]。这种长期进化过程产生的敏感性的差异可能决定了不同鱼类在环境溶氧骤变的适应能力及其生存状况。三峡库区的春泄、秋泄导致水位上升及淤泥中有机质分解等都会导致中下层水体缺氧。然而我国有关环境溶氧变化对鱼类游泳能力的影响研究仅最近在瓦氏黄颡鱼(Peltebagrus vachelli)[14]、南方鲇(Silurus meridionalis)[11]、鲫鱼(Auratus auratus)[12]和金鱼(Carassius auratus)[15]上有报道。鳊鱼(Parabramis pekinensis)和中华倒刺鲃(Spinibarbus sinensis)是两种生活习性较为接近的温水性鲤科鱼类, 广泛分布于长江水域的中下层水体。因此, 本研究在4个不同溶氧水平(8、4、2和1 mgO2/L)条件下通过测定鳊鱼和中华倒刺鲃幼鱼的临界游泳速度和游泳代谢率, 考察急性溶氧变化对游泳能力的影响, 并获取静止代谢率(Resting metabolic rate, MO2rest)、最大游泳代谢率(Maximum metabolic rate, MO2max)、代谢范围(Metabolic scope, MS)及单位位移能耗(Cost of transport, COT)等相关的参数, 旨在揭示不同鲤科鱼类应对环境溶氧的改变所采取的能量代谢适应策略,具有重要的理论意义。同时, 研究数据的获取可为三峡库区鱼类生物多样性保护工作提供更加丰富的资料, 具有重要的应用价值。
1 材料与方法
1.1 实验鱼的来源与驯化
实验鳊鱼、中华倒刺鲃幼鱼均购自重庆市合川水产校, 实验前于实验室驯养2周。实验用水为曝气后自来水, 驯化水温(25 ± 1)℃。溶氧水平≥7 mgO2/L, 每天以商业饲料饱足饲喂1次, 日换水量约为驯化水体的10%, 光制为14 L∶10 D。从驯化2周的幼鱼中挑选身体健康、体重接近的鳊鱼(4.70 ± 0.11) g和中华倒刺鲃(3.26 ± 0.06) g幼鱼各32尾作为实验对象。
1.2 实验方案与操作方法
将适应性驯化后的鳊鱼、中华倒刺鲃幼鱼各随机分为4组(1、2、4和8 mgO2/L, n = 8), 把实验鱼从驯化水槽中依次转移至游泳代谢仪中进行实验测定, 整个实验过程中通过电脑控制向水体环境中充氧气和氮气来调节其氧含量始终维持在各个设定溶氧水平的±0.05 mgO2/L以内, 溶氧通过溶氧仪(HQ10, Hach Company, Ioveland, Colorado, USA)监测。
临界游泳速度测定 实验鱼实验前均禁食2d。实验测定设备采用本实验室自行设计的密闭式鱼类游泳代谢测定仪, 其整体结构和工作原理已在相关文献报道[2,11]。实验时分别将单尾鳊鱼、中华倒刺鲃幼鱼转入游泳管中适应4h, 以消除转移过程中的胁迫影响, 期间管内的流速约为3.0 cm/s (约0.5倍体长, 能保证水体充分交换但不会引起鱼的活动)[11]; 然后按Brett的方法进行Ucrit测定[16]。在测定过程中, 实验鱼在初始速度为5.0 cm/s下持续游泳20min后, 以速度增量(Δv) 5.0 cm/s和持续时间(Δt) 20min不断增加水流速度直至实验鱼达到运动力竭状态, 力竭的评判标准为鱼停在游泳管末端筛板20s以上[17]。
游泳代谢率测定 采用上述设备测定实验鱼在游泳过程中的代谢率, 在测定Ucrit的同时每2min测定一次封闭水体的溶氧值, 以溶氧值随时间变化斜率的绝对值和水体的体积计算每尾鱼每分钟的代谢率。氧含量采用溶氧仪检测。实验结束后将实验鱼取出称重并测量体长。随后将仪器在相应的速度下空转20min得到细菌耗氧量。实验期间游泳代谢仪内水体的溶氧水平不低于设定水平的85%。
1.3 参数计算
实验鱼绝对临界游泳速度(Ua, cm/s)采用以下公式测定:
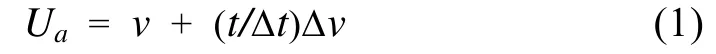
式中, v是实验鱼所具有的次最大速度(完成持续时间的最大速度, 单位为cm/s); 在本研究中Δv 为5.0 cm/s, Δt为20min; t是在最大速度下实际持续时间。为消除体长对游泳速度的影响将绝对游泳速度除以体长获得相对临界游泳速度(Ur), 其计算公式为

式中, BL为实验鱼的体长(单位为cm)。
实验鱼每尾鱼游泳代谢率(MO2perfish, mgO2/h):
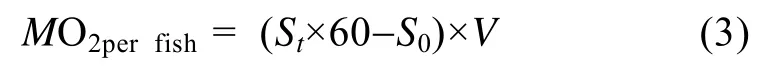
式中MO2perfish(mgO2/h)代表每尾鱼耗氧率, St为每档期间(20min)溶氧值随时间变化的斜率, 60为60min, S0为1h的细菌耗氧, V为运动呼吸仪的系统体积(3.50 L)。为了消除鱼体重不同对耗氧率产生的影响, 以0.75为体重系数把耗氧率标准化为1 kg体重获得标准体重代谢率:
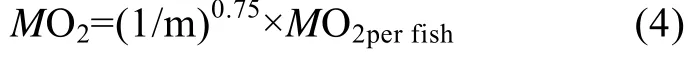
式中MO2[mgO2/(kg·h)]为标准体重代谢率, MO2perfish同(1)式, m为鱼体重(kg)。
由MO2获得单位位移能耗(COT):
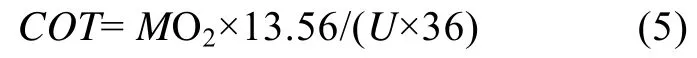
式中COT[J/(kg·m)]表示单位位移能耗, U为游泳速度, 13.56为氧热当量系数(J/mgO2)。
1.4 数据统计
实验数据以Excel2003进行常规计算, 然后采用SPSS17.0进行相关统计分析, 溶氧水平和种类对鱼类Ucrit、MO2rest、MO2max、MS的影响采用双因素方差分析(ANOVA)。溶氧水平、种类和游泳速度对代谢率和COT的影响采用三因素ANOVA, 若有显著差异采用Duncan多重比较。统计值均以平均值土标准误(Mean ± SE)表示, 显著性水平为P<0.05。
2 结果
2.1 不同的溶氧水平对鳊鱼和中华倒刺鲃幼鱼Ucrit的影响
溶氧水平和种类对Ucrit均有显著影响, 且存在交互作用(P<0.05)(表1)。随着溶氧水平的下降, 两种鱼的Ucrit均呈逐渐下降的趋势, 但中华倒刺鲃在较高溶氧水平(4—8 mgO2/L)范围内Ucrit对溶氧变化不敏感。在各溶氧水平下中华倒刺鲃Ucrit均大于鳊鱼(P < 0.05), 但在中等溶氧水平二者差距更大。中华倒刺鲃在1、2、4和8 mgO2/L溶氧水平下的Ucrit分别比鳊鱼高18%、33%、27%和12%(图1)。
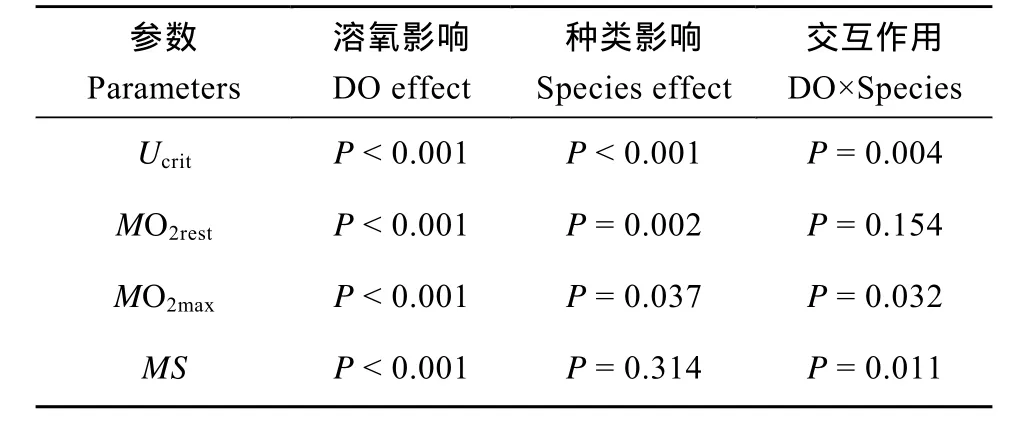
表1 溶氧水平对鳊鱼和中华倒刺鲃影响的双因素方差分析表Tab.1 The results of a two-way ANOVA analysis according to the effect of dissolved oxygen level and species on experimental measured variables
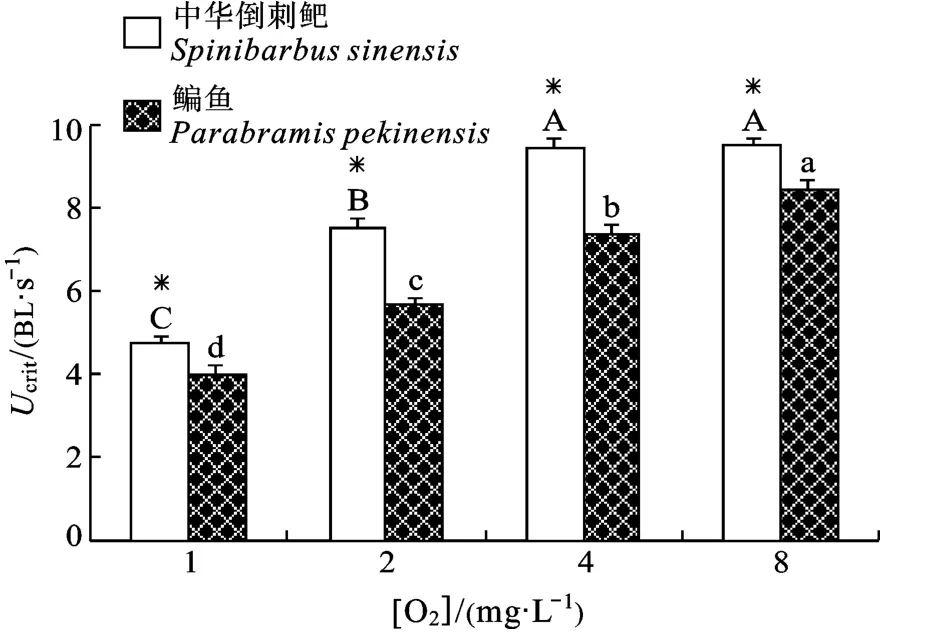
图1 溶氧水平对鳊鱼和中华倒刺鲃幼鱼临界游泳速度的影响Fig.1 Effect of dissolved oxygen level on critical swimming speed in juvenile Parabramis pekinensis and Spinibarbus sinensis a, b, c, d (A, B, C): 不同字母表示不同溶氧水平组鳊鱼(或中华倒刺鲃)差异显著(P<0.05); *: 同一溶氧水平下两种鱼差异显著;下同
2.2 不同的溶氧水平对鳊鱼和中华倒刺鲃幼鱼MO2rest的影响
溶氧水平和种类对MO2rest均有显著影响(P< 0.05), 但无交互作用(表1)。两种鱼1 mgO2/L组的MO2rest均显著小于其他溶氧水平实验组(P<0.05)。中华倒刺鲃的MO2rest显著小于鳊鱼(P<0.05)(图2)。
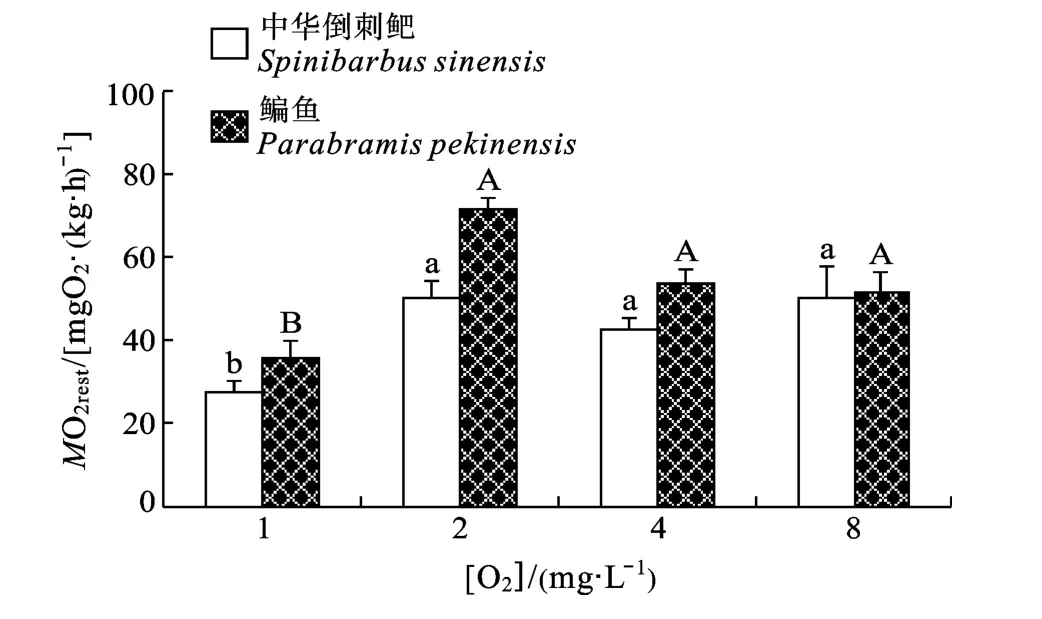
图2 溶氧水平对鳊鱼和中华倒刺鲃幼鱼MO2rest的影响Fig.2 Effect of DO level on resting metabolic rate in juvenile Parabramis pekinensis and Spinibarbus sinensis
2.3 不同的溶氧水平对鳊鱼和中华倒刺鲃幼鱼MO2max的影响
溶氧水平和种类对MO2max均有显著影响, 且存在交互作用(P<0.05)(表1)。随着溶氧水平的下降,两种鱼的MO2max均显著下降(P<0.05)。仅8 mgO2/L溶氧水平下两种鱼的MO2max存在显著差异(P<0.05) (图3)。
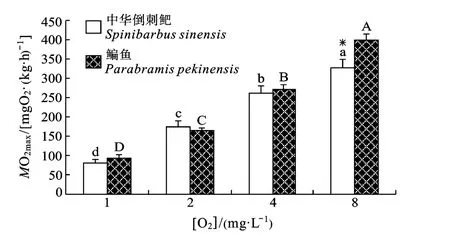
图3 溶氧水平对鳊鱼和中华倒刺鲃幼鱼MO2max的影响Fig.3 Effect of DO level on MO2maxin juvenile Parabramis pekinensis and Spinibarbus sinensis
2.4 不同的溶氧水平对鳊鱼和中华倒刺鲃幼鱼MS的变化
两种鱼的MS均随着溶氧水平的下降而下降,各溶氧水平组间均有显著差异(P<0.05)(表1)。但溶氧水平和种类间存在交互作用, 随着溶氧水平的下降, 鳊鱼的MS下降更为显著, 特别是4—8 mgO2/L (图4)。
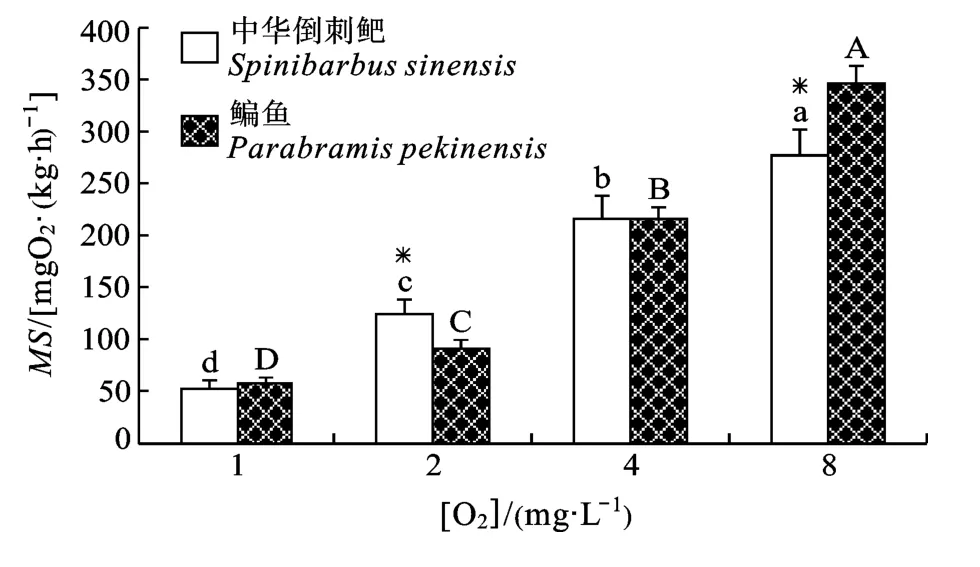
图4 不同溶氧水平对鳊鱼和中华倒刺鲃幼鱼MS的影响Fig.4 Effect of dissolved oxygen level on MS in juvenile Parabramis pekinensis and Spinibarbus sinensis
2.5 不同溶氧水平对两种幼鱼MO2和COT的影响
游速、溶氧水平和种类对特定游速下的代谢率均有显著影响, 且存在显著的交互作用(表2)。在低等游速下, 鳊鱼的MO2和COT显著大于中华倒刺鲃且在高溶氧水平下更为显著(P<0.05)。两种鱼的MO2均随着游泳速度的上升而上升(图5), COT随着游泳速度的上升而下降到20 cm/s以上不再有显著差异。由于不同实验组和鱼类的MO2随游速上升的上升速率不同, 在中等游速下两种鱼的MO2和COT没有显著差异, 而在高速下仅4 mgO2/L组中华倒刺鲃和鳊鱼有显著差异。
3 讨论
3.1 溶氧水平对鳊鱼和中华倒刺鲃COT的影响
在饱和溶氧水平下, 鳊鱼的COT显著大于中华倒刺鲃,在较低游泳速度下更为明显(图5)。这一方面是因为中华倒刺鲃的MO2rest的较低, 另一方面是因为与鳊鱼相比, 中华倒刺鲃体形更为流线型, 运动阻力小, 因此单位位移的能耗较低[18]。随着溶氧水平的下降, 两种鱼的COT均出现不同程度的下降。这一方面是由于低氧下一些生理功能的关闭或下调[2], 另一方面是因为无氧代谢在运动中所起的作用增加[8]。
3.2 溶氧水平对鳊鱼、中华倒刺鲃幼鱼临界游泳速度的影响
鱼类的Ucrit是衡量其有氧游泳能力的常用指标[19], 溶氧水平对鱼类临界游泳速度的影响已有报道。在一般情况下, Ucrit随环境溶氧的下降而降低。然而不同鱼类随溶氧水平的下降其Ucrit下降的幅度及下降的敏感区间不同, 为不同鱼类在适应不同环境的过程中长期进化的结果。研究发现, 同样在25℃条件下, 75%溶氧水平的下降会导致较不耐低氧的海鲈(Dicentrarchus labrax)和黄颡鱼Ucrit下降29% 和31%[13,20]; 但低氧耐受能力较强的鲫鱼和南方鲇仅下降11%和17%[11,12]。本实验发现当溶氧水平的下降50%时, 鳊鱼和中华倒刺鲃的Ucrit分别下降13%和1%(后者没有显著变化), 说明鳊鱼的Ucrit对溶氧水平的变化更为敏感, 而4—8 mgO2/L的溶氧区间为中华倒刺鲃幼鱼游泳运动的耐氧保守区。然而随着溶氧水平的进一步降低, 当下降到12.5%时, 二者Ucrit下降的幅度分别为53%和50%,差异不再明显。
3.3 不同溶氧水平下两种幼鱼的代谢适应对策
随着溶氧的下降, 鱼类有氧运动能力是否下降,与鱼类的代谢对策密切关联。鱼类运动能力是否维持取决于其代谢空间(MS)的变化, 而代谢空间的变化取决于呼吸能力即MO2max和基础代谢消耗即MO2rest的差值[21]。一方面Ucrit与溶氧水平的关系可能受鱼类生态习性的影响, 如伏击取食的鱼类南方鲇, 运动能力弱, 在常氧下Ucrit的限制来自于肌肉组织而不是呼吸系统的供氧能力, 因此水体溶氧水平50%的下降不会对机体运动情况下的摄氧能力产生任何影响, MO2max和Ucrit都不会变化[11]。本研究发现50%DO的下降造成鳊鱼和中华倒刺鲃MO2max下降32%和20%, 说明溶氧水平的下降影响到其呼吸摄氧能力, 但中华倒刺鲃的Ucrit并没有显著变化,这是因为虽然随溶氧水平的下降会导致中华倒刺鲃呼吸能力和MO2max的下降, 但同时机体其他生理功能的下调降低了维持能量消耗和COT, 在适度低氧条件下仍能维持较高水平的有氧运动能力, 其内在生理生化机制还有待进一步探讨。
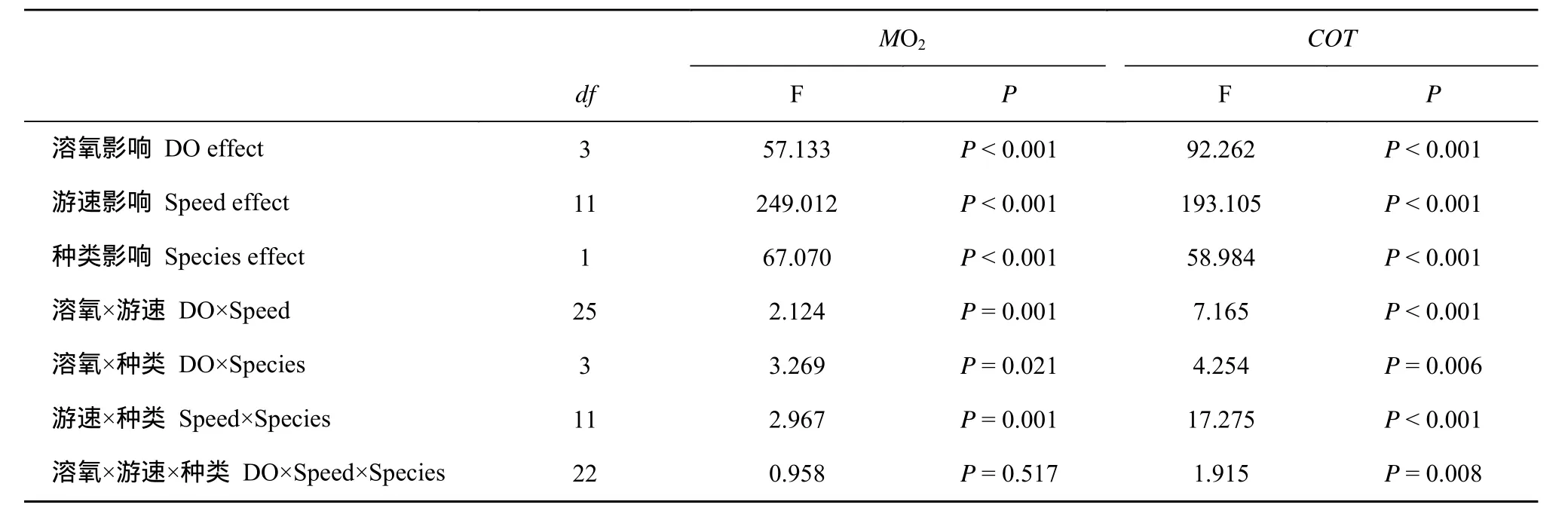
表2 溶氧水平、种类和游泳速度对MO2和COT的影响的方差分析表Tab.2 The results of a three-way ANOVA analysis according to the effect of DO level, species, and swimming speed on MO2and COT
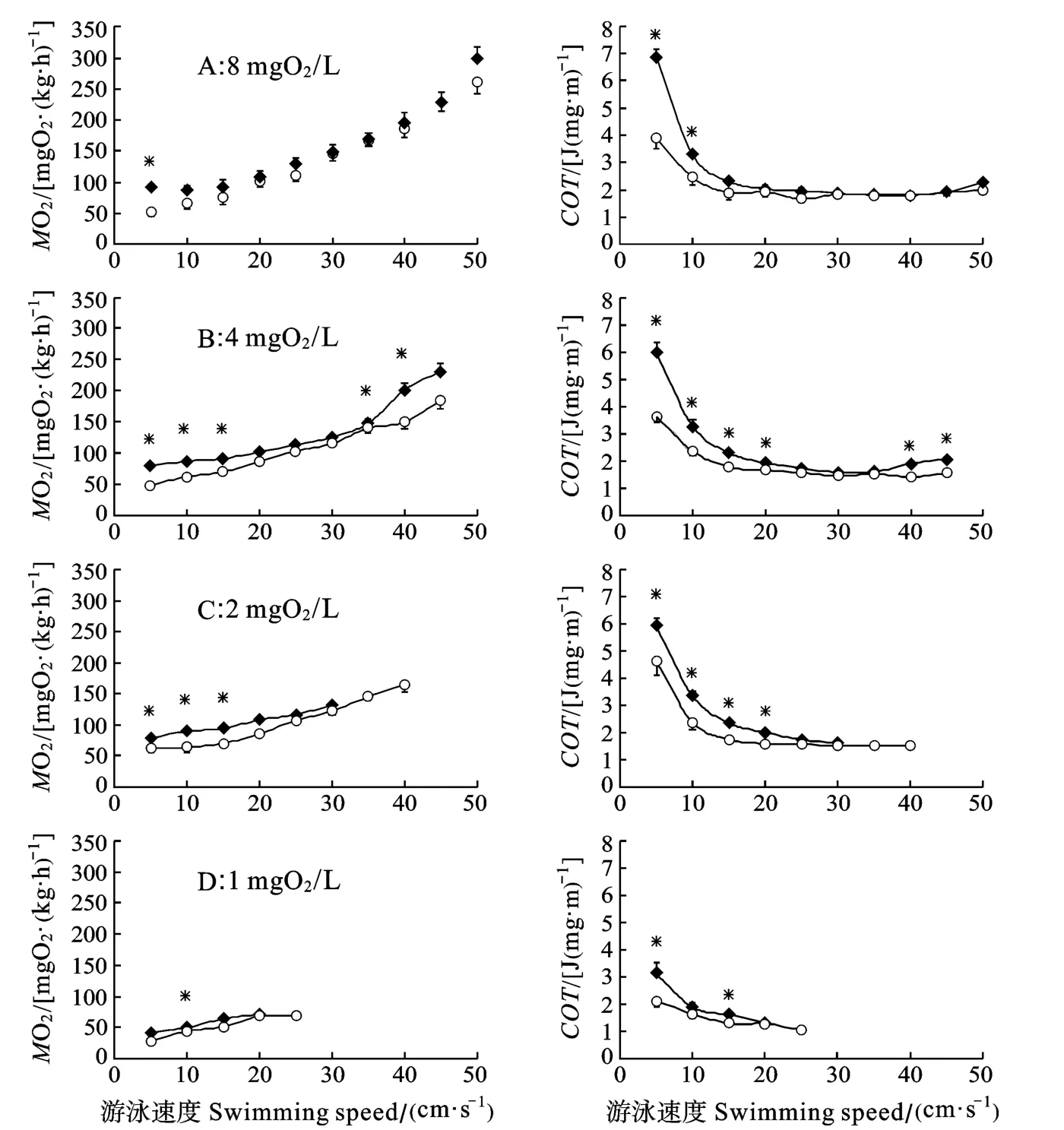
图5 不同溶氧水平下鳊鱼(◆)和中华倒刺鲃(○)幼鱼的游泳代谢率和COTFig.5 Effect of DO level on swimming MO2and COT in juvenile Parabramis pekinensis (◆) and Spinibarbus sinensis (○) swam under different swimming speed
[1] Yuan X, Tu Z Y, Han J C, et al.Effects of flow rate on swimming behavior and energy consumption of Schizothorax chongi [J].Acta Hydrobiologica Sinica, 2012, 36(2): 270—275 [袁喜, 涂志英, 韩京成, 等.流速对细鳞裂腹鱼游泳行为及能量消耗影响的研究.水生生物学报, 2012, 36(2): 270—275]
[2] Fu S J, Brauner C J, Cao Z D, et al.The effect of acclimation to hypoxia and sustained exercise on subsequent hypoxia tolerance and swimming performance in goldfish (Carassius auratus) [J].The Journal of Experimental Biology, 2011, 214(12): 2080—2088
[3] Richards J G.Physiological, behavioural and biochemical adaptations of intertidal fishes to hypoxia [J].The Journal of Experimental Biology, 2011, 214(2): 191—199
[4] Mitrovic D, Dymowska A, Nilsson GE, et al.Physiological consequences of gill remodeling in goldfish (Carassius au-ratus) during exposure to long-term hypoxia [J].American Journal of Physiology R, 2009, 297(1): 224—234
[5] Matey V, Richards J G, Wang Y, et al.The effect of hypoxia on gill morphology and ionoregulatory status in the Lake Qinghai scaleless carp, Gymnocypris przewalskii [J].The Journal of Experimental Biology, 2008, 211(7): 1063—1074
[6] Fu S J, Zeng L Q, Li X M, et al.The behavioral, digestive and metabolic characteristics of fishes with different foraging strategies [J].The Journal of Experimental Biology, 2009, 212(14): 2296—2302
[7] Zeng L Q, Zhang Y G, Fu S J, et al.Effects of acute temperature change on resting oxygen consumption rate and critical swimming speed in juvenile southern catfish (Silurus meridionalis Chen) [J].Acta Hydrobiologica Sinica, 2011, 35(2): 276—282 [曾令清, 张耀光, 付世建, 等.双向急性变温对南方鲇幼鱼静止耗氧率和临界游泳速度的影响.水生生物学报, 2011, 35(2): 276—282]
[8] Zhu Y P, Cao Z D, Fu S J.Muscle lactate and blood glucose levels of juvenile darkbarbel catfish (Pelteobagrus vachelli Richardson) in response to exhaustive exercise [J].Acta Hydrobiologica Sinica, 2011, 34(5): 905—912 [朱晏苹, 曹振东, 付世建.不同游泳速度条件下瓦氏黄颡幼鱼的有氧和无氧代谢反应.水生生物学报, 2011, 34(5): 905—912]
[9] Zeng L Q, Cao Z D, Fu S J, et al.Effect of temperature on swimming performance in juvenile southern catfish (Silurus meridionalis Chen) [J].Comparative Biochemistry and Physiology A, 2009, 153(2): 125—130
[10] Zhang Y, Cao Z D, Fu S J.Effect delayed first feeding on the energy content, body length and swimming performance of southern catfish (Silurus meridionalis Chen) larvae [J].Acta Ecological Sinica, 2007, 27(3): 1161—1167 [张怡, 曹振东,付世建.延迟首次投喂对南方鲇 (Silurus meridionalis Chen) 仔鱼身体含能量、体长及游泳能力的影响.生态学报, 2007, 27(3): 1161—1167]
[11] Zhang W, Cao Z D, Peng J L, et al.The effects of dissolved oxygen level on the metabolic interaction between digestion and locomotion in juvenile southern catfish (Silurus meridionalis Chen) [J].Comparative Biochemistry and Physiology A, 2010, 157(3): 212—219
[12] Zhang W, Cao Z D, Fu S J.Effect of dissolved oxygen level on metabolic mode in juvenile crucian carp [J].Acta Ecological Sinica, 2012, 32(18): 5806—5812 [张伟, 曹振东,付世建.溶氧水平对鲫鱼代谢模式的影响.生态学报, 2012, 32(18): 5806—5812]
[13] Pang X, Cao Z D, Fu S J.The effects of dissolved oxygen on resting oxygen consumption and swimming performance in juvenile darkbarbel catfish (Peltebagrus vachelli) [J].Acta Hydrobiologica Sinica, 2012, 36(2): 255—261
[14] Pang X, Yuan X Z, Cao Z D, et al.The effects of dissolved oxygen levels on resting oxygen consumption and swimming performance in juvenile darkbarbel catfish Peltebagrus vachelli [J].Acta Hydrobiologica Sinica, 2012, 36(2): 255—261 [庞旭, 袁兴中, 曹振东, 等.溶氧水平对瓦氏黄颡鱼幼鱼静止耗氧率和临界游泳运动能力的影响.水生生物学报, 2012, 36(2): 255—261]
[15] Fu S J, Li X M, Zhao W W, et al.The locomotive and metabolic strategies of goldfish under different dissolved oxygen level [J].Journal of Chongqing Normal University (Natural Science), 2010, 27(3): 14—18 [付世建, 李秀明, 赵文文,等.不同溶氧水平下锦鲫的运动和代谢适应对策.重庆师范大学学报(自然科学版), 2010, 27(3): 14—18]
[16] Brett J R.The respiratory metabolism and swimming performance of young sockeye salmon [J].Journal of the Fisheries Research Board of Canada, 1964, 21(5): 1183—1226
[17] Lee C G, Farrell A P, Lotto A, et al.The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O.kisutch) salmon stocks [J].The Journal of Experimental Biology, 2003, 206(18): 3239—3251
[18] Yan G J, He X K, Cao Z D, et al.The trade-off between steady and unsteady swimming performance in six cyprinids at two temperatures [J].Journal of Thermal Biology, 2012, 37(6): 424—431
[19] Shi X T, Chen Q W, Liu D F, et al.Critical swimming speed of young Chinese sucker (Myxocryprinus asiaticus) [J].Acta Hydrobiologica Sinica, 2012, 36(1): 133—136 [石小涛, 陈求稳, 刘德福, 等.胭脂鱼幼鱼的临界游泳速度.水生生物学报, 2012, 36(1): 133—136]
[20] Jourdan-Pineau H, Dupont-Prinet A, Claireaux G, McKenzie D J.An investigation of metabolic prioritization in the European sea bass, Dicentrarchus labrax [J].Physiological and Biochemical Zoology, 2010, 83(1): 68—77
[21] Eliason E J, Clark T D, Hague M J, et al.Differences in thermal tolerance among sockeye salmon populations [J].Science, 2011, 322(6025): 109—112
THE EFFECTS OF DISSOLVED OXYGEN LEVEL ON THE SWIMMING PERFORMANCES OF JUVENILE PARABRAMIS PEKINENSIS AND SPINIBARBUS SINENSIS
ZHAO Wen-Wen, CAO Zhen-Dong and FU Shi-Jian
(Laboratory of Evolutionary Physiology and Behaviour, Chongqing Key Laboratory of Animal Biology, Chongqing Normal University, Chongqing 401331, China)
To investigate the effect of dissolved oxygen (DO) level on swimming performance of juvenile Chinese bream (Parabramis pekinensis) [n=32, body mass: 4.70±0.11 g] and Qingbo (Spinibarbus sinensis) [n=32, body mass: 3.26±0.06 g], we measured the critical swimming speed (Ucrit) and swimming metabolic rate (MO2) and calculated the resting metabolic rate (MO2rest), maximum metabolic rate (MO2max), metabolic scope (MS), and cost of transport (COT) during swimming at 25℃ and DO 8 (100% saturation), 4, 2, and 1 mgO2/L.Ucritvalues of Chinese bream and Qingbo were 8.47 ± 0.19vs9.49 ± 0.14, 7.40 ± 0.17vs9.40 ± 0.28, 5.64 ± 0.16vs7.51 ± 0.21 and 4.02 ±0.23vs4.75 ± 0.13 BL/s at above different DO levels, respectively.Ucritdecreased significantly with DO in both fish species (P<0.05) except that of Qingbo between 4 and 8 mgO2/L.Ucritof Qingbo was significantly higher than those of Chinese bream under all DO levels (P<0.05).Both MO2maxand MS decreased significantly with DO from 8 to 1 (P<0.05) while MO2restshowed a significantly decrease till DO was below 1 mgO2/L.In comparison,MO2increased significantly withUcrit(P<0.05).COTof both fish species decreased withUcritwithin lower swimming speed range but remained unchanged at highUcritrange.COTof Qingbo was lower than that of Chinese bream (P<0.05).In addition,COTdecreased with DO, suggesting that down-regulation of the physiological processes under hypoxic conditions.When DO decreased from 100% to 50%, Chinese bream showed a 13% decrease in Ucritdue to the decreased respiratory capacities (as suggested by a 32% decrease in MO2max) while Qingbo showed no significant decrease in Ucritdespite a 20% decrease in MO2maxdue to the decrease in maintaining metabolism andCOT.However, when DO decreased to 12.5%, both Qingbo and Chinese bream showed similar decreases inUcritcompared to those under 100% DO (53%vs50%).This observation suggested that the swimming performance varied differently with the DO decrease between Chinese bream and Qingbo and was closely related to the metabolic strategy of different fish species.
Parabramis pekinensis; Spinibarbus sinensis; Dissolved oxygen level; Critical swimming speed; Swimming capacity; Metabolic rate; Adaptive strategy
Q591.4
A
1000-3207(2013)02-0314-07
10.7541/2013.20
2012-03-02;
2012-12-28
国家自然科学基金项目(No.31172096); 重庆市自然科学基金(2010BB1089); 重庆市科技创新能力建设项目(CSTC, 2010 CA1010)资助
赵文文(1986—), 女, 山东枣庄人; 硕士研究生; 研究方向为鱼类生理生态。E-mail: zhaowenwen86@126.com
付世建, E-mail: shijianfu9@hotmail.com
猜你喜欢
世界建筑(2022年11期)2022-12-05
知识窗(2022年6期)2022-07-08
工会博览(2022年9期)2022-06-30
中老年保健(2021年10期)2021-11-30
阅读与作文(小学高年级版)(2019年8期)2019-10-16
当代水产(2019年7期)2019-09-03
福建质量管理(2019年2期)2019-01-22
发酵科技通讯(2018年3期)2018-10-16
农家之友(2018年8期)2018-08-24
红领巾·萌芽(2018年3期)2018-04-13
