
紫云英的农艺性状变异
2013-04-25 10:03王建红王松涛
草业科学 2013年8期
张 贤,王建红,曹 凯,王松涛
(1.浙江省农业科学院环境资源与土壤肥料研究所,浙江 杭州 310021; 2.贵州省安顺市西秀区农业局,贵州 安顺 561000)
紫云英(Astragalussinicus)属于豆科黄芪属一年生或二年生草本植物。其被中国、日本等亚洲国家普遍种植,我国长江流域和长江以南各省均有栽培,而以长江下游各省栽培最多[1-2]。我国是紫云英的原产地,也是世界上利用和种植紫云英最早、栽培面积最大的国家,在20世纪六七十年代种植面积达稻区面积的60%~70%[3]。我国紫云英按开花和成熟期可分特早熟种、早熟种、中熟种和晚熟种4类。特早熟种主要分布在气候温暖的广东、广西和福建省(区);早熟种分布的地域较广,在广东、广西、福建、四川、江西、湖南、湖北和河南等省(区)均有分布;中熟种分布在江西、湖南、浙江、安徽、江苏和上海等省(区);晚熟种主要分布在江苏、浙江两省[4]。紫云英茎叶柔嫩多汁、叶量丰富、富含营养物质,可青饲,也可调制干草或青贮料,是一种优质的豆科牧草[1]。同时,紫云英也是我国主要的绿肥作物[5],能有效增加冬闲田地面覆盖,对改良和培肥土壤,提高作物产量和品质,以及促进农区牧业的发展都有着重要作用,在国内外被大力应用于养地培肥[6-8]。
从表型性状来检测遗传变异是最直接的方法,具有快速、简便易行的特点。由于表型和基因型之间存在着基因表达、调控、个体发育等复杂的中间环节,以农艺性状检测遗传变异其实是根据表型的差异来反映基因型的差异。本研究对我国南方不同地区广泛栽培的9个紫云英品种进行农艺性状分析,以揭示其变异程度和变异规律,以期为紫云英的遗传改良及合理开发利用提供科学依据。
1 材料与方法
1.1试验材料 本研究共选取了9个紫云英品种,包括3个早熟种,4个中熟种和2个晚熟种。分别由安徽省农业科学院、江西省农业科学院和福建省农业科学院提供。
宁波大桥:纯度较高的晚熟高产农家品种,原产浙江省宁波市鄞州区,是我国紫云英开花和种子成熟最迟的品种,植株高大、茎秆粗壮。
遂昌:原产浙江省遂昌县,为当地农家品种,属晚熟种。
闽紫1号:福建省农业科学院从四川南充地区农科所的74(3)104-l系统选育而成,为早熟偏迟品种,早发性好、植株较高大、茎秆粗壮。
闽紫6号:福建省农业科学院土壤肥料研究所以江西省南城县的良种植株为母本,浙紫5号的优良株系“浙紫5号-13”为父本进行杂交,经系统选育而成,属中熟偏迟品种。
闽紫7号:福建省农业科学院培育的新品种,苗期生长快、耐阴性较好,为中熟偏迟种。
余江大叶:原产江西省余江县,为江西省优良的地方品种,属中熟偏早品种。
弋江:原产安徽省南陵县大江镇,为安徽省优良的农家品种,栽培历史悠久,属中熟偏早品种。
信阳:原产于河南省信阳市,为河南省优良的农家品种。属早熟种,冬前生长缓慢、开花早、植株矮小、产草量低[3]。
粤肥2号:原名“东莞种”,从广东省原有普通栽培品种选育而得。早生快发、无滞冬期,为早熟品种。
1.2试验地建植 试验地位于浙江省农业科学院杨度试验基地。田间试验采用完全随机区组设计,设有3个重复。在每个重复内各小区随机排列,小区面积为1 m×2 m。试验所用紫云英采取条播方式,于2010年10月播种。播种后覆土1 cm,浇水,随后进行一般牧草田间管理。
1.3试验方法 对12个紫云英农艺性状进行测定,包括株高、叶长、叶宽、茎粗、分枝数、花序数、单株干质量,以及粗蛋白、粗纤维、粗灰分、粗脂肪和无氮浸出物含量。
各小区于盛花期随机选取5个植株测定株高、叶长、叶宽、茎粗、花序数、分枝数和单株干质量,取平均值。具体测定方法如下:叶长为叶基部到叶尖的长度;叶宽为叶中部距离最大处长度;株高为地面至植株顶部绝对高度;茎粗在茎最粗处测量;花序数为每植株花序数;分枝数为每植株分枝数;单株干质量为植株地上部分生物量,采集紫云英地上部分全株样品,65 ℃(48 h)烘干,称重。
于盛花期取各小区紫云英全株样品, 65 ℃(48 h)烘干,粉碎,按四分法取样。分别采用GB/T 6432-1994,GB/6434-2006,GB/T 6438-2007,及GB/T 6433-2006对粗蛋白、粗纤维、粗灰分和粗脂肪含量进行分析。
无氮浸出物以干基计算:
无氮浸出物=(1-粗蛋白-粗脂肪-粗纤维-灰分)×100%。
1.4数据分析 通过变异系数分析紫云英品种内的变异状况,用单因素方差分析及多重比较对农艺性状进行统计分析,并采用系统聚类研究紫云英品种间亲缘关系。统计分析中所用的分析软件为SAS 8.0。
2 结果与分析
2.1紫云英品种内变异特征 变异系数(CV)反映性状均值上的离散程度。变异系数越大,该性状在品种内离散程度越大,品种内个体间差异越大;相反,变异系数越小,品种内离散程度越小,个体间差异越小。12个性状在品种内变异系数差异较大,平均变异系数最小的仅0.040(无氮浸出物含量),而最大的则高达0.310(分枝数)(表1)。12个农艺性状变异系数平均值由大到小依次是分枝数、花序数、单株干质量、茎粗、株高、粗灰分、叶长、粗脂肪、粗蛋白、叶宽、粗纤维和无氮浸出物。变异系数最大的性状是分枝数,不同品种间分枝数在品种内离散程度差异显著,从0至0.417不等。品种内变异系数最小的是无氮浸出物含量,其变异系数远小于其它11个性状。在9个紫云英品种中,无氮浸出物变异系数最大仅为0.046,出现在地方品种宁波大桥种中。所测12个农艺性状中分枝数、花序数和干质量等性状品种内变异系数较大,除了变异系数最小的无氮浸出物含量外,叶宽和粗纤维含量平均变异系数也明显低于其它性状,分别为0.092和0.089。
2.2紫云英品种间变异特征 不同紫云英品种,由于各自不同的遗传背景,其农艺性状变异情况也存在着极大差异。对紫云英株高、粗蛋白含量等12个农艺性状进行方差分析及多重比较(表2),以揭示各性状的变异程度和显著性水平。平均值用来表明各观测值相对集中较多的位置,而标准差能反映数据集合的离散程度。
2.2.1紫云英品种间形态特性变异特征 株高、叶长、叶宽是影响紫云英生物产量的重要因素,在紫云英品种间的变异状况表现出极显著的相关性。叶长在品种内及品种间均存在显著差异(P<0.05)。叶长最长的是宁波大桥(14.22 cm),最短的是粤肥2号(9.06 cm),信阳(10.11 cm)较其略高,二者差异不显著,但均显著低于其余紫云英品种。叶宽从3.75~4.53 cm不等,叶宽最小的仍是粤肥2号(3.75 cm)和信阳(3.85 cm),它们与叶宽最宽的闽紫6号(4.53 cm)和闽紫7号(4.44 cm)均存在显著差异(P<0.05)。9个紫云英品种株高在41.78 cm(粤肥2号)到61.22 cm(闽紫6号)之间,其中闽紫6号株高显著高于余江大叶、粤肥2号、弋江和信阳,其余品种间株高差异不显著。
紫云英分枝数最高的品种是宁波大桥、余江大叶和闽紫7号种,平均分枝数均为3.67个·株-1;最低的品种是信阳种,为2.0个·株-1,显著低于宁波大桥和余江大叶(P<0.05)。其余紫云英品种分枝数差异水平相同,无显著差异。茎粗最粗的品种为闽紫6号,平均茎粗4.55 mm,与其余8个紫云英品种均存在显著差异(P<0.05)。
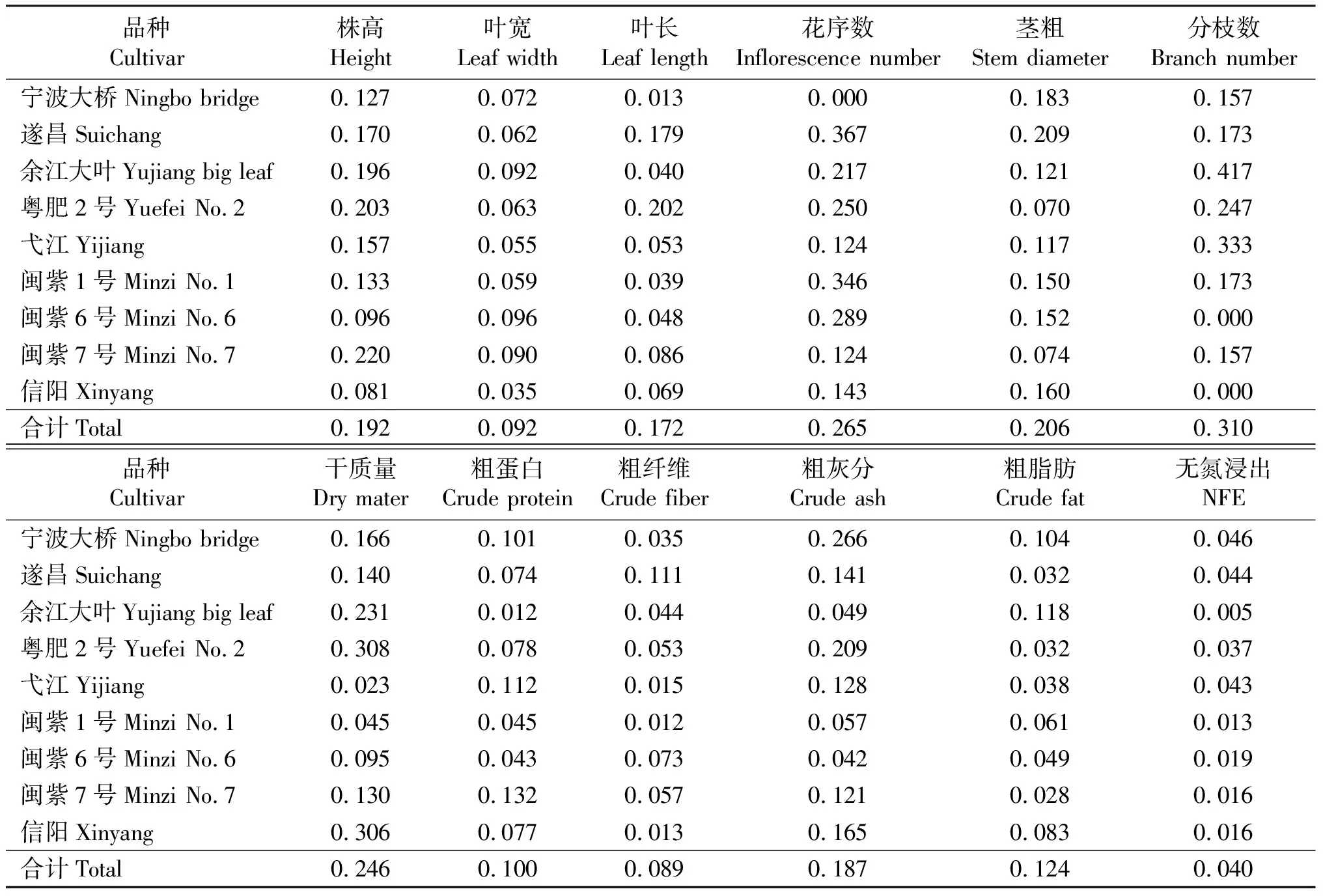
表1 紫云英农艺性状变异系数Table 1 Coefficient of variation of agronomic traits in Chinese milk vetch
紫云英生殖生长相关性状每植株花序数与叶长、叶宽等营养生长性状存在一定的负相关。营养生长较弱的粤肥2号每植株花序数最多,平均值为8个,其与花序数较少的宁波大桥、余江大叶、弋江、闽紫1号及闽紫7号差异显著(P<0.05)。除粤肥2号外,花序数较多的还有信阳及闽紫6号,分别为7个和6个。
9个紫云英品种中单株干质量最高的为中熟种闽紫7号,达4.09 g·株-1。不同品种间干质量差异较大,干质量最低的是浙江当地种遂昌(2.77 g·株-1)及弋江(2.84 g·株-1)。闽紫7号干质量显著高于遂昌、宁波大桥、余江大叶、弋江和闽紫1号(P<0.05)。
2.2.2紫云英品种间营养成分变异特征 所测紫云英品种粗蛋白含量平均值为14.99%,范围13.66%~16.71%(表2)。晚熟种遂昌(16.71%)与中熟种余江大叶(16.25%)粗蛋白含量最高,显著高于早熟种粤肥2号、信阳,及中熟种闽紫6号、闽紫7号(P<0.05);中熟种弋江与早熟种闽紫1号的粗蛋白含量非常接近,在9个品种中属中等水平,与其余品种粗蛋白含量均没有显著差异;粤肥2号、信阳与闽紫6号粗蛋白含量较低,三者差异水平一致;在9个品种中,闽紫7号粗蛋白含量最低,为13.66%。
紫云英品种间粗纤维含量存在显著差异(P<0.05),粗纤维含量平均为17.46%,其中,最高的是信阳,达19.33%,与余江大叶、弋江、闽紫1号、闽紫7号及宁波大桥差异显著(P<0.05);粤肥2号、闽紫6号、遂昌粗纤维含量亦较高,分别为17.97%、18.27%和18.07%,显著高于闽紫7号(P<0.05);粗纤维含量较低的是余江大叶、弋江和宁波大桥。
紫云英品种间粗脂肪含量差异极显著(P<0.05),平均值为3.48%,最高的是弋江(4.03%),粗脂肪含量显著高于除余江大叶和宁波大桥外的其余紫云英品种;粗脂肪含量较高的品种还有余江大叶(3.90%)、宁波大桥(3.63%)和闽紫7号(3.60%);粤肥2号和遂昌粗脂肪含量最低,均为3.10%,粗脂肪含量低的品种还有闽紫6号(3.13%)。
各紫云英品种粗灰分含量平均值为7.29%,范围6.30%~8.57%;宁波大桥粗灰分含量最高;粤肥2号、余江大叶、弋江、闽紫1号、闽紫7号等品种粗灰分含量中等,与所有品种均无显著差异;含量最低的是闽紫6号(6.30%)及信阳(6.37%),显著低于遂昌及宁波大桥(P<0.05)。总的来说,紫云英品种间粗灰分含量的差异低于粗蛋白、粗纤维、粗脂肪等营养成分。
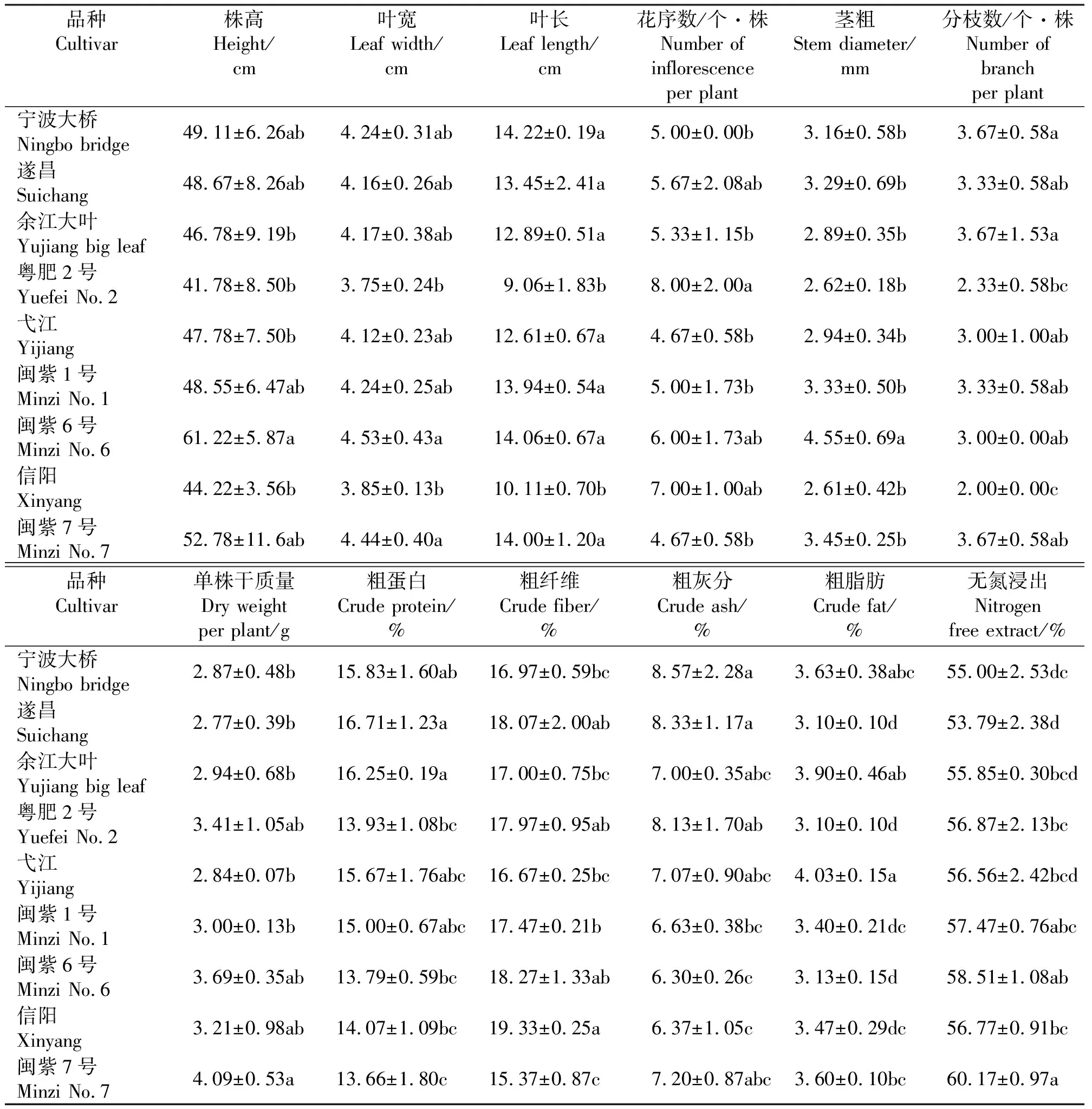
表2 紫云英品种农艺性状Table 2 Agronomic traits in Chinese milk vetch
无氮浸出物是饲料作物干物质中粗蛋白、粗脂肪、粗纤维、灰分以外的营养成分,它的主要成分是淀粉、糖以及它们的近似物,是饲料中很重要营养成分之一。紫云英无氮浸出物含量平均值为56.78%,其中闽紫7号最高(60.17%),遂昌最低(53.79%);闽紫7号和闽紫1号无氮浸出物含量均较高,与除闽紫1号外的其余品种差异显著。
2.3紫云英聚类分析 以12个农艺学性状的平均值计算品种间欧氏距离,并根据欧氏距离进行聚类分析,以揭示紫云英品种间相互关系(图1)。在欧氏距离10处可以将紫云英品种分为3组:中熟偏迟品种闽紫6号首先与其余品种分开,其形态特性表现明显优于其余品种,株高、叶宽、茎粗等性状均为各品种中最高,且叶长、花序数、分枝数、干质量等亦较高,但营养成分含量较低;早熟种信阳及粤肥2号遗传距离较近,聚为一类,这两个品种形态特性和营养成分含量表现均较差;中熟品种闽紫7号、余江大叶、弋江、闽紫1号,以及晚熟种宁波大桥和遂昌各农艺性状表现中等,聚为一类,其中,中熟种余江大叶、弋江和闽紫1号聚为一个小组,同为浙江本地品种的晚熟种宁波大桥和遂昌也聚为一组,与其余中熟种区分开。
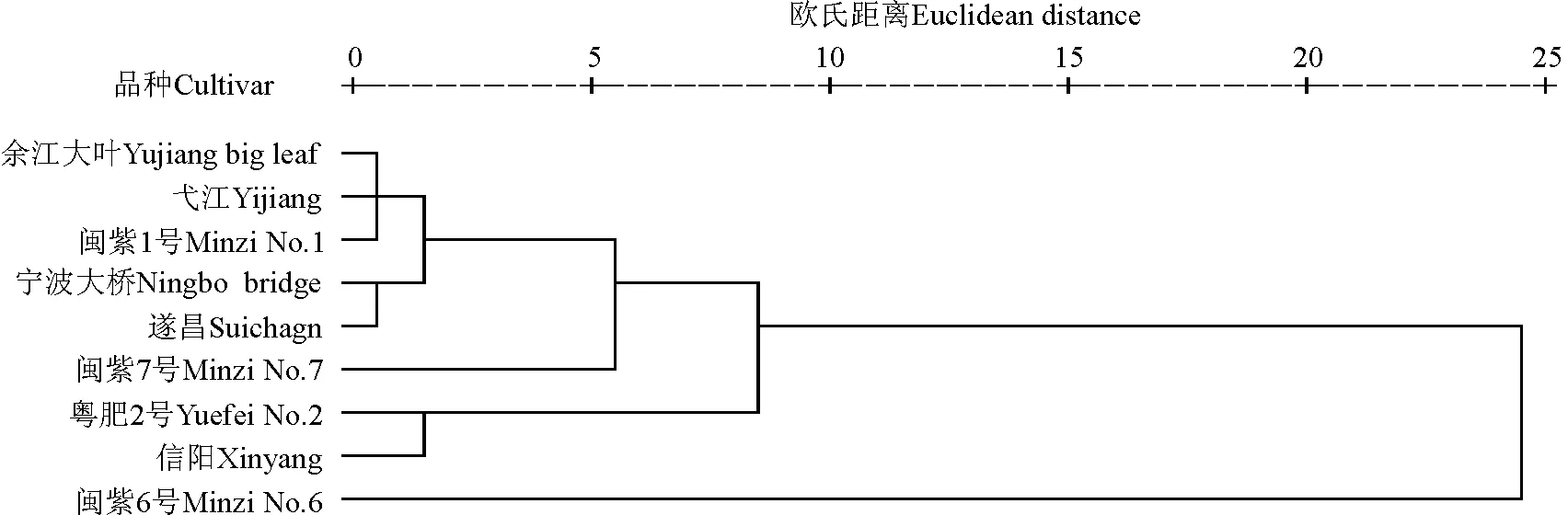
图1 紫云英品种系统聚类Fig.1 Cluster analysis of Chinese milk vetch cultivars
3 讨论与结论
3.1紫云英农艺性状变异特征 表型的变异是基因型、生态型变异丰富度的反应,是基因组遗传变异与环境相互作用的结果。紫云英品种之间的表型性状差异显著,不同品种物候期、鲜草产量、株高、茎粗等性状有明显差异[9]。本研究中,紫云英品种间农艺学性状变异丰富。在不同品种间茎粗、株高、叶长、叶宽差异显著,分枝数在品种间差异较小;营养成分粗蛋白、粗纤维、粗脂肪含量在紫云英品种间差异显著,粗灰分含量的差异低于粗蛋白、粗纤维、粗脂肪。
紫云英表型性状在品种间和品种内均有显著差异。变异系数反映性状均值上的离散程度,是品种内个体间差异大小的体现。所测性状在紫云英品种内变异情况差异显著(P<0.05)。在12个性状中,品种内变异最小的是无氮浸出物含量,其变异系数远远低于其它性状。分枝数、花序数及干质量变异系数最高,在紫云英品种内个体间性状中表现出显著差异。
3.2紫云英品种间遗传距离 群体间遗传距离是用来衡量群体间相似性关系的指标,利用遗传距离构建系统发育树是真实系统发育树的统计学推断,是研究群体间遗传关系的重要方法。孙清信[10]分别采用ISSR和SSR分子标记分析紫云英种质资源的遗传关系,两种方法所获得的聚类结果不同,但品种间遗传距离均未表现出与熟期有相关性。而本研究采用形态学分析紫云英品种间遗传关系,系统聚类结果部分反映了紫云英品种的熟期,熟期相同的品种之间表现出了较近的遗传距离。例如,早熟种信阳与粤肥2号,中熟种弋江与余江大叶,晚熟种遂昌与宁波大桥都分别聚在3个组内。但聚类也不能完全体现紫云英品种的熟期,中熟种闽紫6号就与其他的中熟品种分在不同组内。紫云英表型性状与其开花早晚相关,开花期与株高、分枝数间均呈极显著的正相关,植株高分枝数多的紫云英开花期较迟,鲜草产量相应较高[4]。这可能是形态学检测遗传关系更能反映紫云英品种熟期的原因。
3.3紫云英遗传改良前景 紫云英是异花授粉植物,种群内变异大[11],遗传基础十分广泛,这为紫云英的良种选育提供了很好的基础。与张辉等[12]的研究结果相似,本研究中中熟品种闽紫6号以及闽紫7号在叶片性状、株高、茎粗及单株干质量等形态特性上的表现都明显优于其他品种,营养生长旺盛,可作为高生物产量个体选优的重点。紫云英是优质的豆科牧草,在日粮中补充紫云英可提高反刍动物生长率[13]。粗蛋白对于动植物来说是不可缺少的营养物质,是衡量牧草品质的最主要的指标[14]。一般来说,牧草的粗蛋白、粗脂肪含量越高,粗纤维含量越低,则牧草的营养价值就越高。本研究所测9个紫云英品种中,余江大叶粗蛋白含量显著高于其余品种,粗纤维及粗灰分含量与大部分品种无显著差异,同时其粗脂肪含量高。因此,余江大叶可以考虑用作优质高蛋白紫云英品种,以及作为紫云英优良性状选育的材料利用。
[1] 陈默君,贾慎修.中国饲用植物[M].北京:中国农业出版社,2002:413-414.
[2] 陈宝书.牧草饲料作物栽培学[M].北京:中国农业出版社,2001:236.
[3] 陈坚,张辉,朱炳耀,等.紫云英SSR分子标记的开发及在品种鉴别中的应用[J].作物学报,2011,37(9):1592-1596.
[4] 林多胡,顾荣申.中国紫云英[M].福州:福建科学技术出版社,2008:69-83.
[5] 李勇,陈玉林.紫云英混作栽培在农牧业中的作用[J].中国畜禽种业,2010(11):58.
[6] Dae J K,Dae S C,Sungchul C B,etal.Effects of soil selenium supplementation level on selenium contents of green tea leaves and milk vetch [J].Journal of Food Science and Nutrition,2007,12(1):35-39.
[7] Naomi A,Hideto U.Nitrogen dynamics in paddy soil applied with various 15 N-labelled green manures [J].Plant and Soil,2009,322(1-2):3-4.
[8] Nakayama H.Characteristics of rice (Oryzasativa) growth, yield and soil nitrogen by cultivating Chinese milk vetch (AstragalussinicusL.) as green manure [J].Tohoku Agricultural Research,2005,58:35-36.
[9] 张辉,曹卫东,吴一群,等.不同紫云英品种物候期及主要经济性状研究[J].草业科学,2010,27(2):109-112.
[10] 孙清信.紫云英SSR引物的开发及种质资源遗传多样性分析[D].福州:福建农林大学,2012.
[11] 林新坚,曹卫东,吴一群,等.紫云英研究进展[J].草业科学,2011,28(1):135-140.
[12] 张辉,袁廷茂,杨秉业,等.紫云英新品种(系)比较试验[J].草业科学,2011,28(10):1831-1834.
[13] Jian X L,Jun A Y,Hong W Y.The effects of supplementary Chinese milk vetch silage on the growth rate of cattle and their intake of ammoniated rice straw [J].Animal Feed Science and Technology,1997,65:79-86.
[14] 于辉,姚江华,刘荣,等.四个紫花苜蓿品种草产量、营养品质及越冬率的综合评价[J].中国草地学报,2010,32(3):108-111.
猜你喜欢
农业知识(2022年9期)2022-10-13
上海农业科技(2020年2期)2020-04-02
华东师范大学学报(自然科学版)(2020年1期)2020-03-16
自然资源情报(2018年12期)2018-12-28
中学时代(2018年6期)2018-11-20
海峡姐妹(2018年7期)2018-07-27
作文与考试·小学高年级版(2016年14期)2016-09-10
环球中医药(2015年4期)2015-02-27
党建文汇·上(2014年6期)2014-07-04
新疆农垦科技(2014年9期)2014-02-28
