
菹草(Potamogeton crispus)附着物对水体氮、磷负荷的响应*
2013-05-29 10:37陆全平王国祥毛丽娜魏宏农
湖泊科学 2013年3期
董 彬,陆全平,王国祥,毛丽娜,林 海,周 锋,魏宏农
(1:南京师范大学地理科学学院,江苏省环境演变与生态建设重点实验室,南京 210023)
(2:江苏省淡水水产研究所,南京 210017)
(3:临沂大学资源环境学院,临沂 276000)
随着人类活动对湖泊生态系统影响的加剧,世界许多湖泊大型水生植物逐渐减少,甚至消失[1-3],这一现象引起了全世界湖沼及环境学专家的广泛关注.有研究者推测这可能与富营养化水体中附着物的增加有关[1,4].附着物是沉水植物茎、叶表面的一个复杂的系统,包括附着藻类、菌类、动物、有机和无机碎屑等,可使到达植物表面的光发生衰减,限制植物的光合作用[5-6],使水与植物表面间物质的传输距离和物质传递的阻力增大[7],最终影响植物生长.Asaeda 等发现附着藻对穿叶眼子菜(Potamogeton perfoliatus L.)有遮荫的影响[8],秦伯强等和宋玉芝等也证实了附着物对沉水植物生长有抑制作用[9-11].但是,由于附着物结构和成分复杂,目前还没有标准的研究方法来测定附着物的生物量和一些基本的参数.有些研究者采用与研究浮游生物相似的微观方法,但很容易忽略肉眼可见的较单细胞藻类个体大的生物,因此相关研究多集中在附着藻类[12-14]和附着细菌[15]的种类和数量上.在烘干和燃烧的基础上,采用重量法估计附着有机物和无机物,采用叶绿素a(Chl.a)含量来指示附着藻类可综合反映附着物的特征.
目前,附着物对沉水植物生长期影响的研究较为集中,但对富营养化水体中附着物对沉水植物衰亡影响的研究较少,对沉水植物生长周期内无机碎屑的动态变化的研究亦较少见,而对于系统研究多种不同氮、磷负荷的富营养化水体对沉水植物附着物影响的文献则更鲜见.鉴于此,通过添加不同浓度氮、磷营养盐,运用重量法和叶绿素a 含量定量综合研究不同氮、磷负荷水体对沉水植物菹草(Potamogeton crispus)附着物(附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量)的影响,为富营养化浅水湖泊生态系统的养分循环和沉水植物的衰亡研究提供理论依据.
1 材料和方法
1.1 实验设计
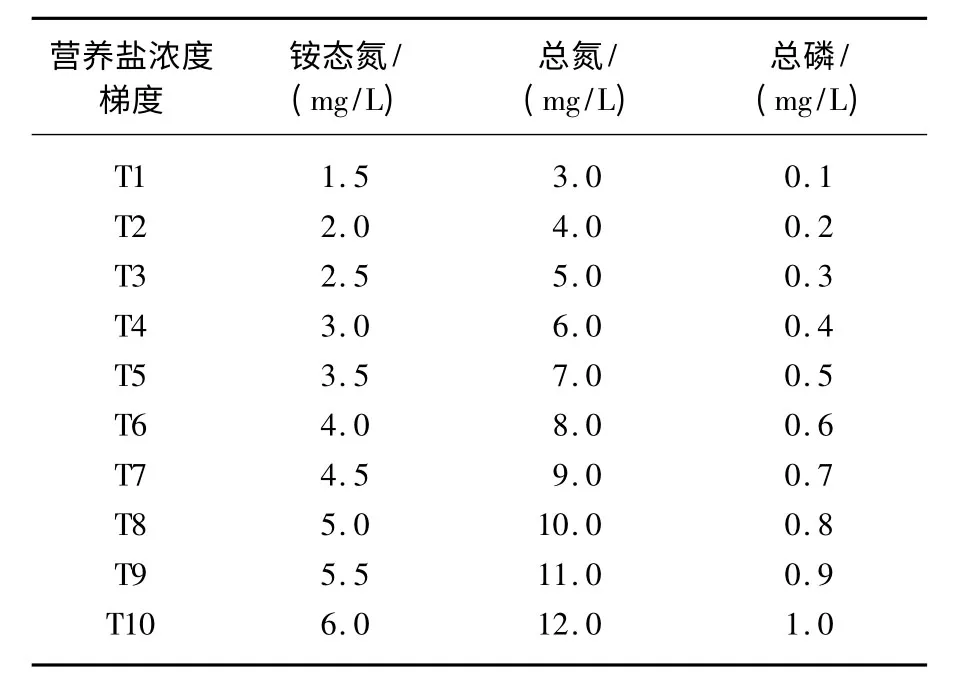
表1 实验水体的氮、磷营养盐浓度梯度Tab.1 Nutrients concentration of experiment waters
从江苏省洪泽湖采集菹草石芽,预处理后,选取大小一致、出芽1 ~2 cm 的石芽供实验.实验容器为直径59 cm、高70 cm 的圆柱形高密度聚乙烯桶,桶底铺10 cm 左右的黄土,经暴晒3 d 后,加自来水至距桶口10 cm处,系统稳定3 周后每桶种植40 株菹草石芽,实验设10 组氮、磷营养盐浓度(表1),每组3 个平行.菹草幼苗长至15 cm 左右时配制 水 体 营 养 盐 (添 加 NH4Cl、NaNO3、KH2PO4)至设定浓度,每周测定水质,及时补充营养盐和因采样、蒸发损失的水分,维持水质稳定在设定浓度范围内.实验在玻璃温室内进行,自2011年11月开始,至2012年5月下旬植株大量死亡分解时结束.
1.2 菹草附着物的采集
分别在菹草快速生长期(3-4月中旬)、成熟期(4月中旬-5月上旬)和衰亡期(5月中旬-6月上旬),从每个实验桶的泥-水界面处随机剪下2 ~3 株菹草装入500 ml 聚乙烯瓶中(采样时力求避免搅动水体以减少植株上附着物的损失),带回实验室24 h 内取出,用软毛刷和无菌水轻轻刷洗植株表面,用显微镜观察确保附着物完全刷下且茎叶表面未受损,刷洗液连同软毛刷冲洗液一并收集,将收集的样品定容至500 ml[8,16].
1.3 附着有机物量、附着无机物量和附着物总量的测定
取200 ml 附着物悬浊液通过Whatman GF/C 滤膜(孔径0.45 μm,抽滤前60℃下烘干至恒重,预先称重,记为M0),用真空泵抽滤后,将带有附着物的滤膜对折,放入坩埚中,在105℃下烘24 h,称重(Dry Weight,DW),记为 M1,然后放入马弗炉,在 550℃燃烧 4 h 后称重(Ash Weight,AW),记为 M2.燃烧损失的质量(M1与M2之差)即为附着物的无灰干重(Ash-Free Dry Weight,AFDW),也表示附着有机物含量[17].附着无机物量为燃烧后的重量(M2)减去滤膜的重量(M0),附着物总量则为烘干重(M1)减去滤膜重(M0).由于菹草的叶面有皱褶,测定其叶面积误差较大,本实验采用单位菹草干重(DW)来表示附着有机物量、附着无机物量和附着物总量,单位为mg/g(DW).
1.4 附着物叶绿素a含量的测定
附着物叶绿素a(Chl.a)含量用90%的丙酮提取后分光光度法进行测定[18].
1.5 水质指标的测定
用连续流动水质分析仪(SKALAR-SAN++,荷兰)分析水体总磷(TP)、总氮(TN)、铵态氮(NH4+-N)、硝态氮(NO3--N)、TP 采用钼锑抗分光光度法测定,TN 采用过硫酸钾氧化紫外分光光度法测定,NH4+-N采用水杨酸分光光度法测定,NO3--N 的测定采用镉柱还原法[18].
1.6 数据处理
采用SPSS 17.0 进行数据处理.统计分析前,对所有的数据先进行正态分布和方差齐性的假设检验.用单因素方差分析(ANOVA)分析营养盐浓度对附着物的影响、生长阶段对附着物的影响,P <0.05为差异性显著,P <0.01 为差异性极显著;用Pearson 相关分析法分析营养盐与附着物、生长阶段与附着物的相关性.
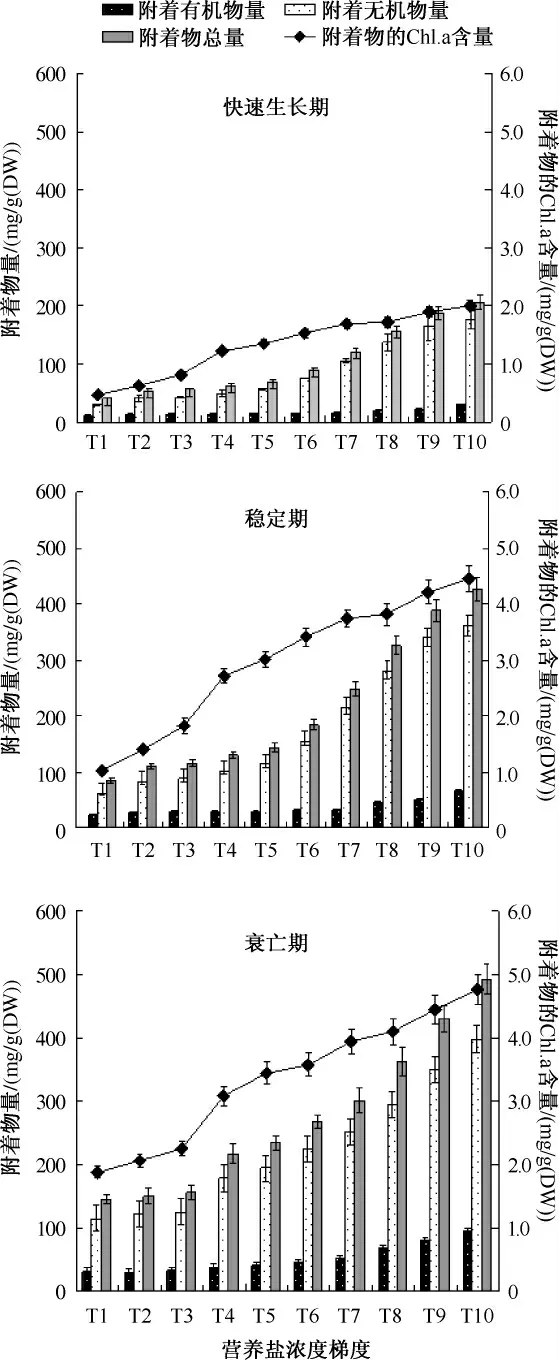
图1 不同营养盐浓度下菹草附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量特征Fig.1 Chlorophyll-a,organic matter,inorganic matter and total periphyton attached on Potamogeton crispus in different concentrations of nutrient
2 结果与分析
2.1 水体氮、磷负荷对菹草附着物的影响
水体氮、磷负荷对沉水植物菹草茎叶附着物的Chl.a 含量、附着有机物量、附着无机物量、附着物总量有显著的促进作用(图1).随着氮、磷负荷的增大,附着物的Chl.a 含量、附着有机物量、附着无机物量、附着物总量呈上升趋势,氮、磷负荷最大的 T10 组(TN 12.0 mg/L,TP 1.0 mg/L)达到最大,且在菹草快速生长期、稳定期和衰亡期均存在这种趋势.T10 组附着物的Chl.a 含量、附着有机物量、附着无机物量、附着物总量分别为2.005 ~4.765、29.027 ~ 94.886、176.881 ~ 397.750 和 205.909 ~ 492.636 mg/g(DW),分别是 T1 组的2.537 ~4.335、2.827 ~3.105、3.489 ~5.883 和 3.408 ~5.105 倍.这可能是因为较高的氮、磷负荷适宜菹草附着藻类和细菌的生长,能促进附着无机物的积累.
2.2 菹草不同生长阶段附着物特征
2.2.1 菹草衰亡期附着物的Chl.a 含量明显高于生长期 菹草附着物的Chl.a 含量在快速生长期、稳定期和衰亡期存在显著差异(P <0.05)(图2a),衰亡期最高,快速生长期最低,且在各营养盐浓度下均存在这一趋势.衰亡期附着物的Chl.a 含量为稳定期的1.046 ~1.826 倍,为快速生长期的2.324 ~4.059 倍.
2.2.2 菹草衰亡期附着有机物量明显高于生长期 同附着物的Chl.a 含量相似,菹草附着有机物量在快速生长期、稳定期和衰亡期存在显著差异(P <0.05)(图2b),衰亡期最高,快速生长期最低,即菹草快速生长季节和生物量较大的季节(成熟期),附着有机物量较少,而菹草衰亡期较多.衰亡期附着有机物量为稳定期的1.046 ~1.638 倍,为快速生长期的2.323 ~3.640 倍.在不同营养盐浓度下均存在这一趋势,且氮、磷负荷越高,差异越显著.
2.2.3 菹草衰亡期附着无机物量明显高于生长期 同附着物的Chl.a 含量和附着有机物量相似,菹草附着无机物量在快速生长期、稳定期和衰亡期也存在显著差异(P <0.05)(图2c),表现为衰亡期>稳定期>快速生长期,在不同营养盐浓度下均存在这一趋势.衰亡期附着无机物为稳定期的1.029 ~1.858 倍,为快速生长期的 2.101 ~3.792 倍.
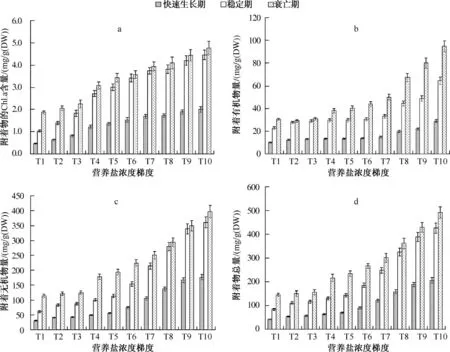
图2 不同营养盐浓度下不同生长阶段菹草附着物的Chl.a 含量(a)、附着有机物量(b)、附着无机物量(c)和附着物总量(d)特征Fig.2 Contents of chlorophyll-a(a),organic matter(b),inorganic matter(c)and total contents(d)of periphyton attached on Potamogeton crispus during different growth periods in different concentrations of nutrients
2.2.4 菹草衰亡期附着物总量明显高于生长期 菹草附着物总量在快速生长期、稳定期和衰亡期亦存在显著差异(P <0.05)(图2d),衰亡期最高,快速生长期最低,在不同营养盐浓度下均存在这一趋势.衰亡期附着物总量为稳定期的1.106 ~1.717 倍,为快速生长期的2.280 ~3.584 倍.
3 讨论
3.1 氮、磷负荷对菹草附着物的影响
本研究发现,氮、磷负荷对菹草茎叶附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量有显著影响.随着氮、磷负荷的升高,菹草茎叶附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量显著升高,氮、磷负荷最大的T10 组附着物量也最大(图1),各浓度间差异均达显著水平(P <0.05).菹草生长的不同阶段,附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量与水体总氮和总磷负荷存在显著相关.Özkan 等[19]在土耳其浅水湖泊中通过中试实验设计了 3 种氮负荷(0.55 ±0.17、2.20 ±0.97、9.20 ±5.45 mg/L TN)和 2 种磷负荷(55 ±19.2、73 ±22.9 μg/L TP),亦发现平均附着物生物量随营养盐浓度升高,且在最高氮和最高磷负荷时达到峰值(0.92 mg Chl.a mg/g(DW));Luttenton 在北密歇根的道格拉斯湖通过人工调控营养盐发现附着藻的生物体积与水体氮浓度显著相关[20];而Liboriussen 等通过对丹麦 13 个湖泊研究发现附着物平均生物量与水体总磷浓度相关,在 60 ~200 μg/L 达到峰值[21];Chen 等[22]通过室内模拟3 种营养盐负荷(TN 0.1 mg/L,TP 0.01 mg/L;TN 1 mg/L,TP 0.1 mg/L;TN 10 mg/L,TP 1.0 mg/L),亦发现随营养盐负荷增高菹草附着藻大量繁殖,本研究结果与其一致,随氮、磷负荷增加附着物Chl.a 含量逐渐升高.本研究附着物的Chl.a 含量和附着有机物含量分别为0.463 ~4.765 和10.269 ~94.886 mg/g(DW),秦伯强等测得梅梁湾中金鱼藻和马来眼子菜的分别为3.843 ~17.609、3.129 ~3.720 mg/g(DW)和24.898 ~26.951、22.897 ~29.200 mg/g(DW)[9],与菹草快速生长期 T10 组、稳定期 T3 组、衰亡期 T1 组的相当.
自然水体中的营养物质可以表现出多种形式,其中有利于附着物生长的P 主要为可溶性有效磷,附着物可利用的N 主要为可溶性无机态氮(如NH4+-N、NO3--N 和NO2--N 等).研究表明适合附着物生长的最佳的 C∶N∶P 摩尔浓度比为 119∶17∶1.如果 N∶P <13 或 C∶N >10,则附着物表现出 N 限制;如果 N∶P >22 或 C∶P >280,则附着物表现出P 限制[23].本研究中,N∶P 依照太湖富营养化水体设计为20 ~25,有利于附着物的生长.在富营养化水体中,附着物可表现出对营养元素的过量吸收,其对水体的吸收速率可达TP 的2 倍以上[24],硝态氮和铵态氮浓度的升高导致丝状藻的大量繁殖[25-26],本研究也发现较高氮、磷负荷的处理中丝状藻较多.沉水植物叶片和水体的养分交换可能因密集的附着物受到抑制,进而影响沉水植物的生长.由此可见,氮、磷负荷对附着物有重要影响,因此恢复高等水生植物来净化湖泊水质,首先要降低营养盐负荷[27].
3.2 菹草生长周期内附着物的动态变化机理
沉水植物茎叶表面附着物的种类组成和数量不仅受水体营养盐负荷[19,21]、植物种类[13,28]及水动力学特征[29]等因素的影响,同时还受生长阶段[12,30]的影响.本研究选择沉水植物菹草生长的3 个典型阶段进行研究,发现附着物的Chl.a 含量、附着有机物量、附着无机物量和附着物总量在快速生长期、稳定期和衰亡期均存在显著差异,均表现为衰亡期最高,快速生长期最低,这与苏胜齐等[30]的研究结果一致.Havens 等亦发现水草在其不同生长阶段,单位面积附着物的生物量及种群结构存在显著差异[31].Rimes 等亦发现沉水植物豆瓣菜(Nasturtium officinale)、Apium nodzjlorum、Glyceria juitans 附着菌类密度 1月末至 2、3月降低,4—6月增加[15].Dijk 也发现蓖齿眼子菜(Potamogeton pectinatus L.)附着物具明显的季节变化,附着物量在5-6月增加,7月降低或保持同等水平,随后增加到一个较高水平[17].沉水植物附着物存在时间分布差异,由于沉水植物本身就有一个生长、成熟、衰亡的过程,所以附着物的时间变化较为复杂.
菹草快速生长期生长旺盛,可能通过分泌化感物质如藻类抗生素抑制附着藻和菌类的增长,或通过保持较高的光合作用组织替代速率[32]来限制附着物生长,使其保持相对较低水平.随着水环境温度的逐渐上升,氮、磷负荷的持续时间增加,沉水植物菹草生物量和表面积增加,但代谢速率逐渐降低,分泌化感物质的速度减缓,附着藻生物量得以持续增加,附着细菌因环境条件适宜继续增长,同时,在菹草叶面逐渐产生大量白色颗粒物,朱端卫等认为可能是水体中钙盐的淀积[33],附着无机物所占比例开始增大,附着物总量较快速生长期有较大提高.T10 组(TN 12.0 mg/L,TP 1.0 mg/L)由于营养盐负荷较高,提前进入衰亡期,到植物衰亡阶段,植物生长几乎停止,茎开始出现断裂,叶片开始出现孔洞,此阶段沉水植物分泌化感物质的能力降到最低,附着细菌通过改变茎叶表面的渗透性促进茎和叶片分解[34],同时叶片通过自溶作用分泌出大量的溶解性有机物,促进附着藻类和菌类的生长[35],水温的增加更适合附着细菌的生长和繁殖,菹草叶面白色颗粒物大面积积累,附着物量达到最大.
由此可见,附着物在沉水植物生长周期内存在明显的动态变化,在沉水植物生长和恢复过程中起着重要作用.在浅水富营养化湖泊植物生态修复过程中,首先应降低水体营养盐负荷以减少附着物积累,增加水体透明度以增强植物对光的吸收利用,然后引进适应性强的先锋水生植物物种,逐步构建以水生高等植物为优势群落的水生生态系统,从而实现湖泊生态系统的健康稳定发展.
[1]Phillips GL,Eminson D,Moss B.A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters.Aquatic Botany,1978,4(1):103-126.
[2]Kemp WM,Boynton WR,Twilley RR et al.The decline of submerged vascular plants in upper Chesapeake Bay:Summary of results concerning possible causes.Marine Technology Society Journal,1983,17(2):78-89.
[3]Sand-Jensen K,Riis T,Vestergaard O et al.Macrophyte decline in Danish lakes and streams over the past 100 years.Journal of Ecology,2000,88(6):1030-1040.
[4]Twilley RR,Kemp WM,Staver KW et al.Nutrient enrichment of estuarine submersed vascular plant communities.Part 1.Algal growth and effects on production of plants and associated communities.Marine Ecology-Progress Series,1985,23(5):179-191.
[5]Sand-Jensen K,Revsbech NP.Photosynthesis and light adaptation in epiphyte-macrophyte associations measured by oxygen microelectrodes.Limnology and Oceanography,1987,32(2):452-457.
[6]Sand-Jensen K.Epiphyte shading:its role in resulting depth distribution of submerged aquatic macrophytes.Folia Geobotanica and Phytotaxonomica,1990,25:315-320.
[7]MacLeod NA,Barton DR.Effects of light intensity,water velocity,and species composition on carbon and nitrogen stable isotope ratios in periphyton.Canadian Journal of Fisheries and Aquatic Sciences,1998,55(8):1919-1925.
[8]Asaeda T,Sultana M,Manatunge J et al.The effect of epiphytic algae on the growth and production of Potamogeton perfoliatus L.in two light conditions.Environmental and Experimental Botany,2004,52(3):225-238.
[9]秦伯强,宋玉芝,高 光.附着生物在浅水富营养化湖泊藻-草型生态系统转化过程中的作用.中国科学:C 辑:生命科学,2006,36(3):283-288.
[10]宋玉芝,秦伯强,高 光.附着生物对太湖沉水植物影响的初步研究.应用生态学报,2007,18(4):928-932.
[11]宋玉芝,黄 瑾,秦伯强.附着生物对太湖常见的两种沉水植物快速光曲线的影响.湖泊科学,2010,22(6):935-940.
[12]Morin JON,Kimball KD.Relationship of macrophyte-mediated changes in the water column to periphyton composition and abundance.Freshwater Biology,1983,13(5):403-414.
[13]Pip E,Robinson G.A comparison of algal periphyton composition on eleven species of submerged macrophytes.Aquatic E-cology,1984,18(2):109-118.
[14]Blindow I.The composition and density of epiphyton on several species of submerged macrophytes:The neutral substrate hypothesis tested.Aquatic Botany,1987,29(2):157-168.
[15]Rimes CA,Goulder R.Temporal variation in density of epiphytic bacteria on submerged vegetation in a calcareous stream.Letters in Applied Microbiology,1986,3(1):17-21.
[16]Pan Y,Stevenson RJ,Vaithiyanathan P et al.Changes in algal assemblages along observed and experimental phosphorus gradients in a subtropical wetland.U.S.A.Freshwater Biology,2000,44(2):339-353.
[17]Dijk GM.Dynamics and attenuation characteristics of periphyton upon artificial substratum under various light conditions and some additional observations on periphyton upon Potamogeton pectinatus L..Hydrobiologia,1993,252 (2):143-161.
[18]国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4 版.北京:中国环境科学出版社,2002:243-284.
[19]Özkan K,Jeppesen E,Johansson LS et al.The response of periphyton and submerged macrophytes to nitrogen and phos-phorus loading in shallow warm lakes:a mesocosm experiment.Freshwater Biology,2010,55(2):463-475.
[20]Luttenton MR.Response of a lentic periphyton community to nutrient enrichment at low N:P ratios.Journal of Phycology,2006,42(5):1007-1015.
[21]Liboriussen L,Jeppesen E.Structure,biomass,production and depth distribution of periphyton on artificial substratum in shallow lakes with contrasting nutrient concentrations.Freshwater Biology,2006,51(1):95-109.
[22]Chen C,Yin D,Yu B et al.Effect of epiphytic algae on photosynthetic function of Potamogeton crispus.Journal of Freshwater Ecology,2007,22(3):411-420.
[23]Rodusky AJ,Steinmann AD,East TL et al.Periphyton nutrient limitation and other potential growth-controlling factors in Lake Okeeehobee,USA.Hydrobiologia,2001,448(1/2/3):27-39.
[24]McCormick PV,oDell MB.Quantifying periphyton responses to phosphorus in the Florida Everglades:a synoptic-experimental approach.Journal of the North American Benthological Society,1996,15(4):450-468.
[25]Irfanullah HM,Moss R.Factors influencing the return of submerged plants to a clear-water,shallow temperate lake.A-quatic Botany,2004,80(3):177-191.
[26]Cao T,Ni LY,Xie P.Acute biochemical response of a submersed macrophyte,Potamogeton crispus L.,to hionium in an aquarium experiment.Journal of Freshwater Ecology,2004,19(2):279-284.
[27]Sagrario G,Maria A,Jeppesen E et al.Does high nitrogen loading prevent clear-water conditions in shallow lake at moderately high phosphorus concentrations?Freshwater Biology,2005,50(1):27-41.
[28]Jeppesen E,Sondergaard M,Meerhoff M.Shallow lake restoration by nutrient loading reduction some recent findings and challenges ahead.Hydrobiologia,2007,196(5):239-252.
[29]由文辉.淀山湖着生藻类群落结构与数量特征.环境科学,1999,20(5):59-62.
[30]苏胜齐,沈盎绿,姚维志.菹草着生藻类的群落结构与数量特征初步研究.西南农业大学学报,2002,24(3):255-258.
[31]Havens KE,Hauxwell J,Tyler AC et al.Complex interactions between autotrophs in shallow marine and freshwater ecosystems:implications for community responses to nutrient stress.Environmental Pollution,2001,113(1):95-107.
[32]Sand-Jensen K.Effect of epiphytes on eelgrass photosynthesis.Aquatic Botany,1977,3(1):55-63.
[33]朱端卫,朱 红,倪玲珊等.沉水植物驱动的水环境钙泵与水体磷循环的关系.湖泊科学,2012,24(3):355-361.
[34]Schreiber L,krimm U,Knoll D et al.Plant-microbe interactions:identification of epiphytic bacteria and their ability to alter leaf surface permeability.New Phytologist,2005,166(2):589-594.
[35]Baker JH,Orr DR.Distribution of epiphytic bacteria on freshwater plants.Journal of Ecology,1986,74(1):155-165.
猜你喜欢
草业学报(2022年5期)2022-07-08
草业学报(2021年3期)2021-03-22
疯狂英语·读写版(2021年2期)2021-03-01
江苏农业科学(2020年19期)2020-12-09
园林(2020年8期)2020-09-18
小学阅读指南·高年级版(2020年3期)2020-04-01
花卉(2020年12期)2020-01-09
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
草业学报(2018年10期)2018-10-19
草业科学(2018年7期)2018-07-26
