
白藜芦醇对白血病K562细胞增殖、凋亡的影响及机制探讨
2013-07-05 09:39隋涛,白雪,马立,李青
山东医药 2013年43期
隋 涛 ,白 雪,马 立,李 青
(天津市第一中心医院,天津300192)
白藜芦醇是一种具有多种生物活性的天然植物成分,自然界许多植物中有白藜芦醇及其苷类。白藜芦醇是植物在遇到真菌感染、紫外线照射时所产生的植物补体,对植物起保护作用。近年研究[1,2]发现,白藜芦醇在人体内具有调节脂质代谢、抑制血小板聚集、抗炎、抗肿瘤等多种生物活性和药理作用,具有很大的药用价值。然而,对于白藜芦醇的抗肿瘤效果争议则很大,低浓度或高浓度的白藜芦醇对不同的肿瘤有抑制或促进作用[3~5]。为了解白藜芦醇在抗白血病中的作用,我们观察了白藜芦醇对人白血病细胞K562生长的影响,并对其相关作用机制进行了探讨。现将结果报告如下。
1 材料和方法
1.1 主要试剂 白藜芦醇购自Sigma公司,胎牛血清和RPMI1640培养基购自Invitrogen公司,CKK-8试剂盒购自同仁化学研究所,Annexin V-FITC/PI双染凋亡检测试剂盒购自美国BD公司。白藜芦醇用DMSO溶解,实验时用RPMI1640稀释成不同浓度。
1.2 细胞培养 将K562细胞培养在含10%胎牛血清、100 U/mL青霉素、100μg/mL链霉素和2 mmol/L谷氨酰胺的 RPMI1640培养基内,于37℃、5%CO2、饱和湿度条件下培养,每2~3 d传代1次。细胞处于对数生长期时用于实验。
1.3 白藜芦醇对K562细胞增殖影响的观察 采用CCK-8法。将处于对数生长期的K562细胞以1×104/每孔接种于96孔板。实验组分别加入终浓度为5、10、15、30、60 μmol/L 的白藜芦醇,对照组加入等量的DMSO,每组设3个复孔。置37℃、5%CO2湿润培养箱中分别孵育 24、48、72、96 h,每孔加入20μL的CCK-8溶液,继续培养2 h后吸去上清液,加入DMSO 150μL,用酶标仪测定450 nm处吸光度值(A值)。按照公式计算细胞生长抑制率:细胞生长抑制率=(1-实验组A值/对照组A值)×100%。实验重复3次。
1.4 白藜芦醇对K562细胞凋亡影响的观察 采用AnnexinV-FITC/PI双染法。将对数生长期的K562细胞以3×106/每孔接种于24孔板,分别加入终浓度为 5、10、15、30、60 μmol/L 的白藜芦醇培养48 h后,PBS洗涤3次,加入5μL的AnnexinV-FITC和1μL的PI混均,避光孵育10 min,PBS洗涤1次,每管加入400μL的PBS,采用流式细胞仪检测凋亡细胞,计算细胞凋亡率。
1.5 白藜芦醇对K562细胞周期影响的观察 采用流式细胞仪检测K562细胞周期。将对数生长期的K562细胞以3×106/每孔接种于24孔板,分别加入终浓度为 5、10、15、30、60 μmol/L 的白藜芦醇培养48 h后PBS洗涤3次,加入70%冷乙醇固定细胞,4℃过夜后离心弃上清,PBS重悬细胞后加入PI染液,4°C避光孵育30 min,用流式细胞仪检测细胞周期,流式细胞仪自带软件分析细胞周期的分布。
1.6 K562细胞中与细胞凋亡和周期相关的蛋白检测 采用Western blot法。将对数生长期的K562细胞以4×106/每瓶接种于25 cm2的培养瓶中,用终浓度为60μmol/L的白藜芦醇处理48 h后按常规方法裂解K562细胞,提取细胞总蛋白。BCA法进行蛋白浓度定量,取30μg总蛋白进行SDS-PAGE电泳,电泳后将SDS-PAGE凝胶上的蛋白转移至PVDF膜上。5%脱脂奶粉室温封闭 1 h,加入1∶1 000稀释的一抗(Bcl-2、Bcl-xl、Bax、Cyclin D1)4℃孵育过夜,加辣根过氧化物酶标记的二抗,室温孵育1 h后ECL显影,测各蛋白灰度值。以β-actin作为内参照。对照组细胞以DMSO代替白藜芦醇进行培养,操作同上。
1.7 统计学方法 采用SPSS16.0统计软件。计量资料组间比较用方差分析,两组间比较用t检验。P≤0.05为差异有统计学意义。
2 结果
2.1 白藜芦醇对K562细胞增殖的影响 随着白藜芦醇浓度逐渐增加和作用时间的延长,K562细胞的生长抑制率上升(P均<0.05),见表1。
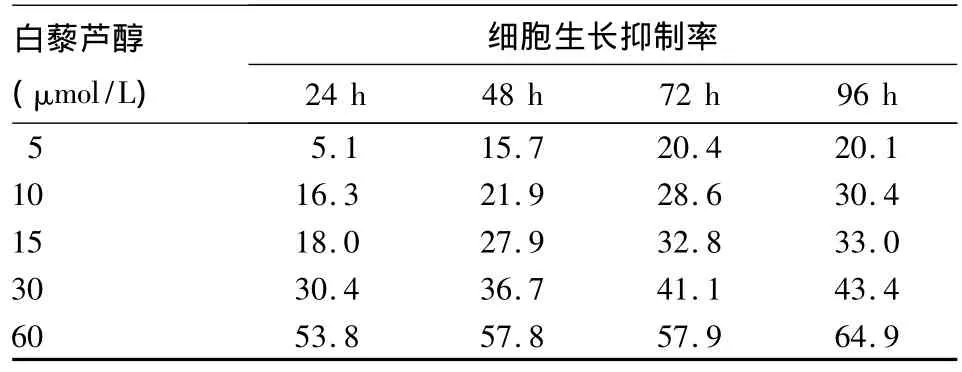
表1 白藜芦醇对K562细胞增殖的影响(%)
2.2 白藜芦醇对K562细胞凋亡的影响 K562细胞经不同浓度白藜芦醇作用48 h后,细胞凋亡显著增加,且以早期凋亡为主,K562细胞的凋亡率随白藜芦醇浓度的增高而上升(P<0.05),且晚期凋亡的K562细胞也逐渐增加,见表2。
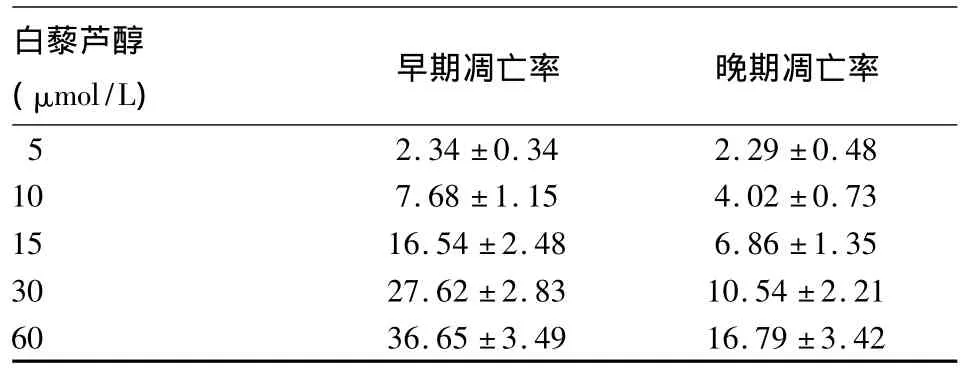
表2 白藜芦醇对K562细胞凋亡的影响(%,¯x±s)
2.3 白藜芦醇对K562细胞周期的影响 经白藜芦醇作用48 h后,K562细胞周期发生明显变化,随着白藜芦醇浓度的增加,G1期细胞逐渐增多(P<0.05),而 S期细胞逐渐减少(P <0.05),表明白藜芦醇可阻止K562细胞从G1期进入S期,进而抑制K562细胞的增殖,见表3。
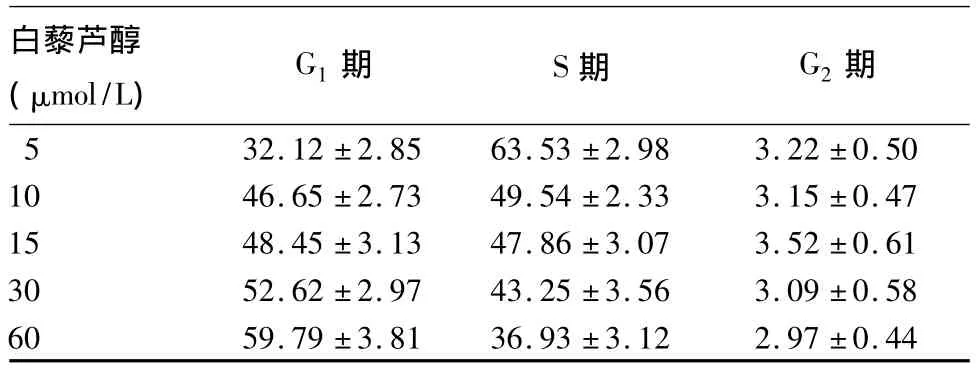
表3 白藜芦醇对K562细胞周期的影响(%,¯x±s)
2.4 白藜芦醇对K562细胞中凋亡相关蛋白表达的影响 实验组K562细胞经60μmol/L的白藜芦醇作用48 h后,抗凋亡蛋白Bcl-2和Bcl-xl表达减少(Bcl-2和Bcl-xl灰度与β-actin灰度比值分别为0.54±0.09 和0.36 ±0.11),而促凋亡蛋白 Bax表达增多(Bax灰度与β-actin灰度比值为1.46±0.28)。实验组凋亡相关蛋白的表达与对照组(Bcl-2、Bcl-xl、Bax灰度与 β-actin 灰度比值分别为 1.03 ±0.23、1.12 ±0.24、0.75 ±0.15)相比,P 均 <0.05。
2.5 白藜芦醇对 K562细胞中细胞周期蛋白Cyclin D1表达的影响 实验组K562细胞经60 μmol/L的白藜芦醇作用48 h后,Cyclin D1表达显著降低(Cyclin D1灰度与β-actin灰度比值为0.43±0.13),与对照组(Cyclin D1灰度与 β-actin灰度比值为1.16 ±0.27)相比,P <0.05。
3 讨论
近年研究[6~11]发现,白藜芦醇能诱导多种肿瘤细胞的凋亡,如卵巢癌细胞、胃肠肿瘤细胞、肝癌细胞、肾癌细胞及胶质瘤细胞等。因此,白藜芦醇在肿瘤治疗方面的价值逐渐引起人们的关注[12]。本研究结果显示,白藜芦醇以时间和剂量依赖的方式抑制白血病K562细胞的增殖并诱导其凋亡。细胞凋亡是细胞的一种主动性死亡过程,是受细胞内源性基因、酶和特定信号通路调控的一个级联激活过程,相关基因包括促凋亡基因和凋亡抑制基因。Bcl-2家族是最重要的凋亡调节因子之一,Bcl-xl与Bcl-2具有相似的抑制细胞凋亡的功能,而Bax则是凋亡促进因子,它们通过调节线粒体的功能来调控细胞凋亡。当细胞受到凋亡刺激后是否凋亡取决于Bcl-2/Bax、Bcl-xl/Bax 平衡。文献[13]报道,白藜芦醇通过改变Bcl-2蛋白家族的表达降低多种肿瘤细胞线粒体膜电位,增加细胞色素C的释放,进而活化Caspases。本研究结果显示,白血病K562细胞经白藜芦醇作用后其Bcl-2与Bcl-xl的表达水平降低,而Bax的表达水平升高。提示白藜芦醇在不同类型的肿瘤细胞通过共同的信号途径诱导细胞凋亡,包括K562细胞。
Cyclin D1是重要的细胞周期调节蛋白,在G0~G1期细胞中表达,通过与细胞周期依赖性蛋白激酶CDK4和CDK6结合,促进细胞从G1期进入S期,使细胞发生增殖与分裂[14]。Cyclin D1表达上调是诱导细胞周期调控异常导致肿瘤发生的主要原因之一。研究[15]证实,多种肿瘤细胞中Cyclin D1过度表达。本研究结果显示,白藜芦醇作用后K562细胞中处于G1期的细胞增多,而处于S期的细胞减少,同时其Cyclin D1表达下降。提示白藜芦醇通过下调Cyclin D1的表达,延迟K562细胞从G1期进入S期,发挥其抑制白血病K562细胞增殖的作用。
总之,白藜芦醇在体外可以抑制白血病K562细胞增殖并诱导其凋亡。其抗K562细胞增殖的效果可能与下调Cyclin D1的表达进而延迟细胞周期从G1期进入S期有关,其诱导K562细胞凋亡可能与上调Bax表达、下调Bcl-xl和Bcl-2表达有关。
[1]Muqbil I,Beck FW,Bao B,et al.Old wine in a new bottle:the warburg effect and anticancer mechanisms of resveratrol[J].Curr Pharm Des,2012,18(12):1645-1654.
[2]胡凌云,王海钠,裴俊俊,等.白藜芦醇防癌抗癌作用及其分子机制研究进展[J].山东医药,2010,50(5):111-112.
[3]Castillo-Pichardo L,Cubano LA,Dharmawardhane S.Dietary grape polyphenol resveratrol increases mammary tumor growth and metastasis in immunocompromised mice[J].BMC Complement Altern Med,2013,13(5):6.
[4]Zhu W,Qin W,Zhang K,et al.Trans-resveratrol alters mammary promoter hypermethylation in women at increased risk for breast cancer[J].Nutr Cancer,2012,64(3):393-400.
[5]Pozo-Guisado E,Merino JM,Mulero-Navarro S,et al.Resveratrol-induced apoptosis in MCF-7 human breast cancer cells involves a caspase-independent mechanism with downregulation of Bcl-2 and NF-kappa B[J].Int J Cancer,2005,115(1):74-84.
[6]Soto BL,Hank JA,Darjatmoko SR,et al.Anti-tumor and immunomodulatory activity of resveratrol in vitro and its potential for combining with cancer immunotherapy[J].Int Immunopharmacol,2011,11(11):1877-1886.
[7]Roy SK,Chen Q,Fu J,et al.Resveratrol inhibits growth of orthotopic pancreatic tumors through activation of FOXO transcription factors[J].PLoSOne,2011,6(9):25166.
[8]Wang Z,Li W,Meng X,et al.Resveratrol induces gastric cancer cell apoptosis via reactive oxygen species,but independent of sirtuin1[J].Clin Exp Pharmacol Physiol,2012,39(3):227-232.
[9]Vergara D,Simeone P,Toraldo D,et al.Resveratrol downregulates Akt/GSK and ERK signalling pathways in OVCAR-3 ovarian cancer cells[J].Mol Biosyst,2012,8(4):1078-1087.
[10]郭亮,张新恒,张楠,等.白藜芦醇对人肾癌细胞增殖和细胞周期的影响[J].山东医药,2011,51(39):20-21.
[11]Gagliano N,Aldini G,Colombo G,et al.The potential of resveratrol against human gliomas[J].Anticancer Drugs,2010,21(2):140-150.
[12]Hsieh TC,Wu JM.Resveratrol:biological and pharmaceutical properties as anticancer molecule[J].Biofactors,2010,36(5):360-369.
[13]Low IC,Chen ZX,Pervaiz S.Bcl-2 modulates resveratrol-induced ROS production by regulating mitochondrial respiration in tumor cells[J].Antioxid Redox Signal,2010,13(6):807-819.
[14] Musgrove EA,Caldon CE,Barraclough J,et al.Cyclin D as a therapeutic target in cancer[J].Nat Rev Cancer,2011,11(8):558-572.
猜你喜欢
国际呼吸杂志(2019年4期)2019-03-12
中成药(2018年8期)2018-08-29
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
华南农业大学学报(2015年5期)2015-12-04
医学研究杂志(2015年5期)2015-06-10
中国药理学通报(2014年2期)2014-05-09
现代检验医学杂志(2014年6期)2014-02-02
