
工业属性普鲁兰酶的开发及其催化性能改善的研究进展
2013-10-25 06:25聂尧,严伟,徐岩
生物加工过程 2013年1期
聂 尧,严 伟,徐 岩
(江南大学 生物工程学院 教育部工业生物技术重点实验室,无锡 214122)
普鲁兰酶属于淀粉脱支酶,可切割普鲁兰和支链淀粉的α-1,6-糖苷键。它作用于支链淀粉时,可切下支链淀粉的整个支链,形成直链淀粉。当它作用于普鲁兰时,主要产物为麦芽三糖。普鲁兰酶是重要的工业用酶,在生产糖浆的淀粉加工工业中,通常与其他的淀粉水解酶(如α-淀粉酶、β-淀粉酶和葡糖淀粉酶)复合使用[1]。因此,研究可水解α-1,6-糖苷键的普鲁兰酶有利于高效获得具有结构多样性和性质独特的多糖,在食品、制药、能源和高聚物材料等领域有重要的商业价值[2],是工业上提高生产效率、增加经济效益的有效途径。而且,作为研究糖类物质结构的工具,普鲁兰酶也受到了极大的关注[3]。
根据水解的糖苷键类型,普鲁兰酶可以被分为两类:I型普鲁兰酶(EC 3.2.1.41,专一性切割普鲁兰的α-1,6-糖苷键,又称限制性糊精酶、支链淀粉-6-葡萄糖苷水解酶)和II型普鲁兰酶(EC 3.2.1.3,切割淀粉及相关多糖的α-1,4和α-1,6糖苷键,又称淀粉普鲁兰酶)[4]。
尽管国内外研究学者已经对普鲁兰酶进行了大量研究,但目前能够满足工业需求具有工业属性的普鲁兰酶的开发仍较为有限。微生物来源普鲁兰酶的分泌、诱导[5]和表达研究的开展以及嗜酸性分解普鲁兰多糖芽胞杆菌 (Bacillus acidopullulyticus)普鲁兰酶3D结构[6]和肺炎克雷伯氏菌(Klebsiella pneumoniae)普鲁兰酶晶体结构的解析[7]为普鲁兰酶的分子改造和高效应用提供了新的依据。
本文主要综述了普鲁兰酶在淀粉糖浆工业中的应用及产酶微生物的开发、异源表达、酶分子结构功能和性质改良等方面的研究内容,以期为普鲁兰酶的高效表达、酶的分子改造及催化性能改善提供理论依据。
1 普鲁兰酶在淀粉糖浆工业中的应用
当前,普鲁兰酶主要应用于淀粉糖浆工业。在糖浆工业中,使用的大部分淀粉质原料含有约75%~85%的支链淀粉,如玉米中支链淀粉约占74%、马铃薯占76%[8]。支链淀粉是一种高度支链化的多糖,平均每20~25个D-葡萄糖单元就有一个分支点,因此支链淀粉含有4% ~5%的α-1,6糖苷键[9]。通常酶法水解淀粉分为液化和糖化两个过程。在液化过程中需使用内作用的α-淀粉酶切割淀粉的α-1,4糖苷键,使其成为相对分子质量相对较低的糊精;在糖化过程中需添加葡萄糖淀粉酶(即糖化酶)将糊精完全分解为葡萄糖或添加β-淀粉酶将糊精分解为麦芽糖[10]。以酶法(α-淀粉酶和糖化酶)生产葡萄糖的最高转化率DE值为96%,这是酶法生产糖浆的极限数值(DE值是指还原糖(以葡萄糖计)占糖浆干物质量的百分比)。这是由于淀粉糖化过程中使用的糖化酶(即葡萄糖淀粉酶)对α-1,6糖苷键的作用效率低,而β-淀粉酶遇到α-1,6分支点则停止作用[9],因此支链淀粉中的α-1,6糖苷键对糖浆的生产无疑是障碍。若在糖化过程中加入可作用于α-1,6糖苷键的脱支酶,则糖化的效率可显著提高。
普鲁兰酶是比较理想的脱支酶。在糖化过程中,普鲁兰酶与糖化酶共同作用,普鲁兰酶切割分支点,糖化酶只需切割线性的低聚糖,可显著地提高淀粉的水解效率,不仅降低糖化酶用量,还可使DE值提高到97%以上,提高水解产物葡萄糖或麦芽糖的质量和纯度。由于淀粉的糖化过程在较高的温度(55~60℃)、微酸性条件(pH 4.5~5.5)下进行,因此应用于糖浆工业的普鲁兰酶需要有较好的耐热耐酸性能[10]。
Norman等[9]将微生物普鲁兰酶与黑曲霉糖化酶联用,研究了普鲁兰酶的添加对糖化过程的影响,发现未加入普鲁兰酶时,D-葡萄糖最大质量分数为96%,而使用对照组一半用量的葡萄糖淀粉酶和0.25 U/g干物质量的普鲁兰酶,D-葡萄糖最大质量分数提高0.5%,继续提高普鲁兰酶用量可使葡萄糖质量分数提高1.5%。并且双酶联用可增大糖化过程的底物浓度。在未加普鲁兰酶的对照组中,当底物干物质量分数为32%时,得到96%的D-葡萄糖,而使用对照组一半用量的葡萄糖淀粉酶和和1 U/g干物质量的普鲁兰酶时,可在高达37%的底物干物质量分数下获得相同水平的D-葡萄糖。另外,典型的糖化反应需要48~96 h,而通过加入普鲁兰酶则可缩短反应时间。当添加0.4 U/g干物质量的普鲁兰酶时,获得相同水平D-葡萄糖的反应时间甚至可缩短到30 h左右。
Norman等的研究结果对淀粉糖浆工业有重要的经济意义,因为普鲁兰酶的加入首先减少了葡萄糖淀粉酶的用量,不仅节约了生产成本,而且葡萄糖淀粉酶用量的减少还可减少异麦芽糖的生成,提高产物纯度。其次,在更高的底物浓度下进行糖化,可大大降低产物蒸发时的成本。另外糖化反应时间的缩短可增加生产批次,大幅提高经济效益[9]。
2 普鲁兰酶产生菌种及其比较
普鲁兰酶主要存在于植物[11]和微生物中,前者的普鲁兰酶被称为R-酶[11]。自从1961年Wallenfels等[12]从肺炎克雷伯氏菌(Klebsiella pneumoniae)中发现普鲁兰酶,迄今为止,已有许多的微生物普鲁兰酶被发现和研究。文献中常见的普鲁兰酶产生菌株见表1。
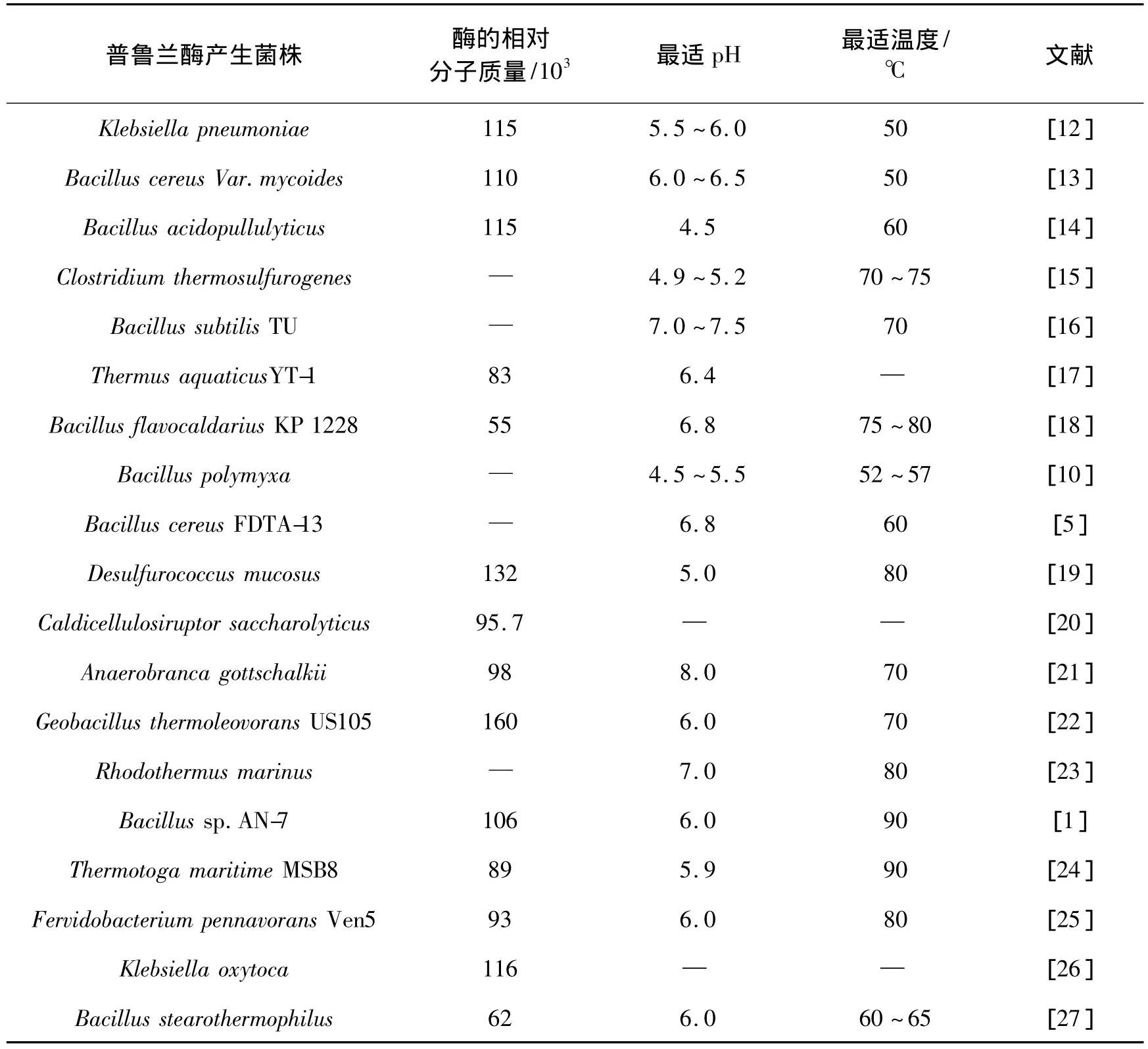
表1 普鲁兰酶产生菌株及其部分性质Table 1 Different microorganisms producing pullulanase and their partial properties
淀粉的糖化过程在57~63℃、pH 4.0~4.5的条件下进行,因此,要求普鲁兰酶需要有较好的耐热耐酸性能。由表1可以发现,除B.acidopullulyticus外,其他微生物的普鲁兰酶均不太符合糖化过程的要求,这也是B.acidopullulyticus可以被用于生产商品普鲁兰酶的原因之一。
淀粉的液化在95~105℃下进行,糖化条件则通常为60 ℃、pH 4.5 ~5.5[10],由液化转入糖化时需要降低温度并加酸降低糖浆的pH。若在液化的条件下可以同时进行糖化,既可以省去加酸碱调节pH的过程以简化工艺、降低成本,又可以有效地防止微生物污染,并且在液化的高温下,还可以获得较高的反应速率。因此,人们希望将液化和糖化过程合并,即在液化的条件下用耐高温的糖化酶和普鲁兰酶实现糖化。但早期所发现的普鲁兰酶的耐热性都较差,最适作用的温度偏低,因此不适合在液化过程的条件下作用。
为获得耐高温的普鲁兰酶,研究人员开始从极端嗜热微生物中筛选普鲁兰酶产生菌。获得的普鲁兰酶生 产 菌 种 包 括 D.mucosus[19]、A.gottschalkii[21]、G.thermoleovorans US105[22]、R.marinus[23]、Bacillus sp.AN-7[1]、T.maritime MSB8[24]、F.pennavorans Ven5[25]等。这些微生物的普鲁兰酶最适温度均在70℃以上,部分甚至达到90℃,基本符合在液化过程高温下水解淀粉的要求。
3 普鲁兰酶的异源表达
虽然学者们获得了许多产普鲁兰酶的微生物,但这些原始菌种的大多数发酵酶活很低,无法应用于工业生产。自20世纪90年代以来,采用基因工程技术将普鲁兰酶编码基因克隆进宿主中,构建高产的重组菌,已被许多学者采用,并且已获得了一些产酶能力强于原始菌的重组菌(表2)。常用的宿主菌为大肠杆菌和枯草芽胞杆菌,也有在酵母中表达的报道[28-29],但由于普鲁兰酶主要来源于原核微生物,因此普鲁兰酶的真核表达报道极少。

表2 普鲁兰酶基因在原核系统中的异源表达Table 2 Heterologous expression of pullulanase genes in prokaryotic systems
3.1 普鲁兰酶在大肠杆菌中的表达
Takizawa等[31]将 K.aerogenes 70 的普鲁兰酶基因pul克隆至pBR322质粒,并构建了重组大肠杆菌。该重组大肠杆菌产生比野生型K.aerogenes W70多3~7倍的普鲁兰酶。将相同的含有 pul基因的pBR322转入野生型K.aerogenes W70,则使普鲁兰酶总产量提高20~40倍,胞内酶水平比原始W70菌高100~150倍。这证明了pBR322上的pul基因可以在大肠杆菌和K.aerogenes中表达,并且普鲁兰酶产量可大幅提高。Bertoldo等[25]将编码极端嗜热厌氧菌F.pennavorans Ven5的Ⅰ型普鲁兰酶基因pulA克隆至大肠杆菌。不带信号肽的pulA基因被亚克隆进表达载体pSE420,并在trc启动子控制下在大肠杆菌中过量表达。该不带信号肽的普鲁兰酶酶活比原始菌株产生的普鲁兰酶高122倍,比鸟枪法获得的克隆子高40倍。Zouari等[22]通过使用2种不同的信号肽,使G.thermoleovorans US105(PUL US105)的Ⅰ型普鲁兰酶在大肠杆菌周质空间或胞外高效分泌。其中,带有Ssamy信号肽的重组大肠杆菌用于考察普鲁兰酶分布,质粒上的Ssamy-pul US105先由lac启动子控制,酶活测定显示胞外酶活最高,胞内只有胞外的10%,周质无酶活,此时普鲁兰酶酶活相对较低。然后将lac启动子替换成trc启动子,结果比酶活提高8倍(12 U/mg)。
但将普鲁兰酶编码基因克隆至异源宿主并不能总是获得过量表达的重组菌。Michaelis等[37]将K.pneumoniae的pulA基因克隆至大肠杆菌,pulA基因的表达受到麦芽糖调节子的malT调节基因控制。载有单个pulA基因拷贝的重组菌的普鲁兰酶表达量只有野生型K.pneumoniae的10% ~20%,可能的原因是由于大肠杆菌的malT蛋白对pulA启动子活性不高导致pulA基因在大肠杆菌中转录效率不高。另一个可能的原因是普鲁兰酶在大肠杆菌细胞膜中的不适当定位导致在大肠杆菌中不稳定。
3.2 普鲁兰酶在枯草芽胞杆菌中的表达
宿主菌的安全性是食品级酶制剂发展的最重要因素之一。目前,有几种微生物通常被认为是安全的,如 E.coli K-12,B.subtilis和A.niger,它们被广泛地用作生产食品级酶的宿主细胞[38]。枯草芽胞杆菌(B.subtilis)作为外源基因表达的宿主,具有如下优点:不含内毒素,为非致病的土壤微生物;有很多噬菌体和质粒可作为枯草芽胞杆菌的克隆载体,并且有可利用的强启动子;具有很强的分泌蛋白质的能力,分泌的蛋白质具有天然活性,且大多数蛋白被分泌到胞外[39]。普鲁兰酶主要应用于食品工业,因此常用枯草芽胞杆菌作为表达普鲁兰酶的宿主菌。
Duffner等[19]将超高温厌氧古菌 D.mucosus的普鲁兰酶基因apuA克隆至质粒pJA803,并在宿主菌B.subtilis DN1885中表达。apuA基因由α-淀粉酶启动子(PamyM)控制。收集发酵上清液,经过阴离子交换、疏水层析和凝胶过滤后,得到的纯普鲁兰酶比酶活为26 U/mg。重组活性蛋白的表观相对分子质量为6.6×104,与D.mucosus的原始酶相同。该高温古菌普鲁兰酶在枯草杆菌中表达量较低(15 mg/L),原因可能有mRNA稳定性、密码子偏好、蛋白质折叠与分泌。进一步研究发现:D.mucosus普鲁兰酶有典型的古细菌密码子偏好,N端的22个氨基酸有9个大肠杆菌的稀有密码子,因此密码子偏好性可能是表达水平低的重要原因。
4 普鲁兰酶结构与功能分析
4.1 普鲁兰酶的分子结构
普鲁兰酶的一级结构可分为4个结构域:A、B、C、F[40-41]。结构域A有一些区域与8个α螺旋和8个β链一致,形成中心的催化的(β/α)8-折叠筒。一个小的结构域B以环的形式插入到第3个α螺旋和第3个β链间[42-43]。结构域B参与形成活性中心和折叠筒的壁。结构域C有8个β链,而且,结构域C连接到结构域A的C-末端,有时结构域F连接到结构域A的N-末端[44-45]。结构域F有一个短的α螺旋和6个β链,折叠为形成β-三明治基序的β-反平行片层[46]。
嗜酸性分解普鲁兰多糖芽胞杆菌(B.acidopullulyticus)普鲁兰酶的3D结构已获得解析[6]。该普鲁兰酶(BaPul13A)的成熟蛋白含921个氨基酸,由CBM41-X45a-X25-X45b-CBM48-GH13_14等部分形成了多结构域的蛋白质结构(图1)。其中CBM表示糖类结合结构域,X表示未知功能的组件,GH13_14为CAZY GH13结构域。该酶的3D结构见图2。CBM41 N端结构域在晶体中高度无序,无法模拟其结构,与其他酶比对后推测该结构域可能有糖原/淀粉结合功能。X45a-X25-X45b功能未知,CBM48为糖原/淀粉结合结构域,GH13_14为催化组件,属GH13家族,其自身含有α-淀粉酶共有的3个典型的A、B、C结构域。因此,BaPul13A成熟蛋白为含有7个结构域的蛋白质,其3个催化结构域连接到4个N端辅助的结构域。
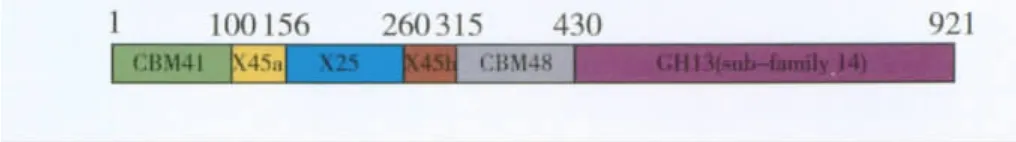
图1 BaPul13A分子结构示意[6]Fig.1 Schematic diagram of the modular structure of BaPul13A[6]
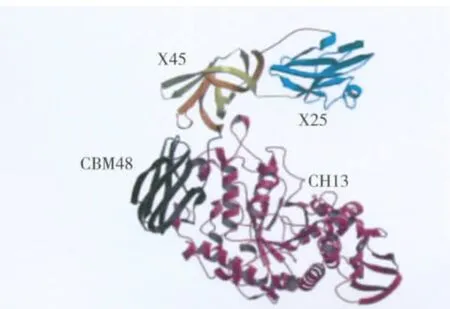
图2 BaPul13A的3D结构Fig.2 3-D structure of BaPul13A
4.2 普鲁兰酶的重要氨基酸位点
Liebl等[47]研究发现,几乎所有的普鲁兰酶都含有高度保守的区域Ⅰ~Ⅳ,这些区域组成一个活性中心和共有的底物结合位点。此外,Kuriki等[48]发现普鲁兰酶催化位点中的2种氨基酸:精氨酸(Arg)和谷氨酸(Glu),对酶的催化活力十分重要。他们还发现,Asp-206、His-210和His-296在底物结合位点发挥重要作用[49]。
为探索普鲁兰酶的功能位点,Yamashita等[50]替换了 K.aerogenes普鲁兰酶重要位点的氨基酸His-607、Asp-677、His-682 和 His-833,这些氨基酸分别位于普鲁兰酶的4个保守区。His-607、Asp-677和His-833的氨基酸替换导致酶活完全丧失。相比之下,His-682的替换仍保留了酶活。测定这些突变体对普鲁兰酶的竞争性抑制剂——α-或β-环糊精的结合亲和力后发现,His-833的突变不影响普鲁兰酶对α-环糊精的亲和力,而His-607和Asp-677的突变体导致丧失其对普鲁兰的结合能力。这些结果表明K.aerogenes普鲁兰酶的His-607、Asp-677位点参与普鲁兰酶与底物的结合,而His-833位点与催化有关。
4.3 普鲁兰酶的保守序列
许多普鲁兰酶的氨基酸序列中都有4个保守区。所有的Ⅰ型普鲁兰酶都具有共有序列YNWGYDP[50],YNWGYDP 基序位于酶的中部,分析显示该基序可能与底物结合或催化活力相关。Ⅱ型普鲁兰酶含有与Ⅰ型酶不同的特异序列,2种普鲁兰酶的序列的最大不同在于共有序列Ⅱ和Ⅳ。Ⅱ型普鲁兰酶的共有序列Ⅱ可被分为两部分,其中第二部分位于共有序列Ⅲ和Ⅳ中间[4]。另外,Berloldo等[51]研究显示,古细菌Ⅱ型普鲁兰酶不含YNWGYDP共有序列及保守区域Ⅰ~Ⅳ。
4.4 普鲁兰酶中脯氨酸含量与热稳定性的关系
酶分子的热稳定性通常与其氨基酸组成和含量有关。Suzuki等[52]研究发现,增加 β-转角和总的疏水性氨基酸中脯氨酸的频率,芽胞杆菌的寡聚-1,6-葡萄糖苷酶的热稳定性可得到提高。Matthew等[53]的实验亦证明了上述发现。Suzuki等[54]研究噬菌体T4溶菌酶时,通过在一个β-转角处用脯氨酸(Pro)替换了丙氨酸(Ala),降低了酶分子骨架的折叠熵,从而提高了溶菌酶的热稳定性。因此,该发现被称为脯氨酸理论。
Suzuki等[18]比较 K.pneumoniae、B.acidopullulyticus和B.flavocaldarius的普鲁兰酶时发现,脯氨酸理论适用于这3种菌的普鲁兰酶:①以K.pneumoniae→B.acidopullulyticus→B.flavocaldarius的顺序,3 种细菌的普鲁兰酶脯氨酸含量依次升高,热稳定性也随之提高;②比较结构参数,发现极端嗜热菌B.flavocaldarius的普鲁兰酶仅在疏水相互作用上超过中温菌的普鲁兰酶。B.flavocaldarius普鲁兰酶含有更多的脯氨酸、甘氨酸、苯丙氨酸、精氨酸和亮氨酸,而丝氨酸、异亮氨酸、赖氨酸、苏氨酸、缬氨酸和天冬氨酸则比中温菌的普鲁兰酶少。大量的脯氨酸、苯丙氨酸和亮氨酸有助于增强疏水相互作用,精氨酸、组氨酸、赖氨酸和天冬氨酸几乎不影响离子键势能。疏水相互作用被认为是稳定嗜热酶最重要的因素[55]。
脯氨酸理论对蛋白质工程意义重大。利用该理论,可创造出热稳定性优良的普鲁兰酶变体,可为提高普鲁兰酶的热稳定性提供切实可行的策略。
5 改善普鲁兰酶热稳定性的其他方式
在淀粉糖化过程中,普鲁兰酶的热稳定性非常重要。良好的热稳定性允许提高糖化反应温度和底物浓度,并阻止微生物生长[56]。虽然研究人员从极端嗜热微生物中分离出了产耐高温普鲁兰酶的菌种,但这些菌种的普鲁兰酶热稳定性依然不太理想。如R.marinus的普鲁兰酶,虽然最适作用温度达到80℃,但在85℃下的半衰期只有30 min[23]。因此,需要通过一些技术手段增强普鲁兰酶的热稳定性。
5.1 底物类似物对普鲁兰酶热稳定性的影响
Kusano等[56]发现,大于麦芽三糖醇的麦芽寡糖能够完全保护普鲁兰酶免遭热钝化,这可能是由于酶与底物类似物的相互作用稳定了普鲁兰酶的四级结构。鉴于此,Kusano等研究了D-葡萄糖醇(G1-OH)、麦芽糖醇(G2-OH)、麦芽三糖醇(G3-OH)、麦芽四糖醇(G4-OH)和麦芽五糖醇(G5-OH)对B.acidopullulyticus普鲁兰酶热稳定性的影响。研究发现,普鲁兰酶在0.56、0.28、0.14和0 mol/L的D-葡萄糖醇中60℃热处理90 min后,残余酶活依次下降,说明普鲁兰酶的热稳定性依赖于D-葡萄糖醇的浓度。同时还发现,随着作为热保护剂的糖醇的葡萄糖基增加,普鲁兰酶稳定性提高。另外,高浓度的麦芽糖醇也可稳定其他的普鲁兰酶。这些结果说明了糖醇的添加可提高普鲁兰酶的热稳定性,葡萄糖基数较多的糖醇可作为普鲁兰酶的热保护剂应用于糖浆工业。
5.2 固定化普鲁兰酶
酶的固定化是提高酶热稳定性的有效措施之一,而且使用固定化酶可实现工业酶促过程的连续操作。目前科研人员已通过固定化技术成功提高了普鲁兰酶热稳定性。
Singh等[2]将 B.acidopullulyticus普鲁兰酶共价固定在通用软化树脂Duolite XAD761上。固定化普鲁兰酶显示了比游离酶更好的温度特性。pH 5.5下最适温度达到60℃,比游离酶高10℃;65℃下热处理6 h后保留了85%的酶活,70℃下处理3 h残余32.38%的活力,而70℃下游离酶则迅速失活。固定化普鲁兰酶对普鲁兰、可溶性淀粉和糊精的最大反应速率分别为4.0、4.17和5.00 U/g,均高于游离酶。循环使用25批后,固定化普鲁兰酶的酶活下降到初始的一半,表明该固定化普鲁兰酶具有较好的重复利用能力。
Kuroiwa等[57]通过普鲁兰酶的氨基和凝胶表面的醛基多点结合,将K.pneumoniae的普鲁兰酶用琼脂凝胶固定化。50℃热处理8 h后,游离酶完全失活,而相比之下固定化的酶则相当稳定,热处理48 h后残余30%的酶活。并且发现,一个普鲁兰酶分子连接的醛基密度为17.9个醛基/3600×10-20m2时固定化酶的热稳定性最好,在pH 6.0的磷酸缓冲液中45℃热处理200 h后依然保留60%的酶活。
6 结语
对于普鲁兰酶的研究,国内主要集中在产酶菌种的筛选与鉴定。国外则研究得较深入,目前已对普鲁兰酶基因与酶分子的结构、酶的定位与分泌机制有了较清楚的认识。但对普鲁兰酶在分子水平上进行改造,以获得酶学性质更加优良的普鲁兰酶变体,则研究较少。
从普鲁兰酶的研究现状来看,现有菌种产生的普鲁兰酶在酶学性质上依然不能完全满足工业需求,因此需要寻找更多酶学性质更好的产生菌种。自然界中超过99%的原核微生物无法在实验室中培养[58],其中含有大量普鲁兰酶产生菌。采用常规的方法无法获得这些非培养微生物的普鲁兰酶编码基因。宏基因组学已被用于从未培养微生物中寻找新的功能基因,并成功地获得了蛋白酶、甘油水解酶和氧化还原酶等多种酶类的编码基因。将宏基因组学应用于普鲁兰酶基因的寻找,将很可能获得更多新的普鲁兰酶编码基因[59]。
另一方面,虽然人们已获得产普鲁兰酶的基因工程菌,但普鲁兰酶表达水平依然普遍较低,所产的普鲁兰酶酶学性质仍有缺陷,并且重组芽胞杆菌的重组质粒稳定性不够理想。因此,今后的研究工作一方面需要寻找和构建更加优良的表达载体,采用信号肽选择、启动子改造、翻译起始区优化等手段实现重组菌普鲁兰酶的高效表达;另一方面,可以采用定向进化等分子改造手段改造普鲁兰酶本身,进一步改善其酶学特性,如提高普鲁兰酶对底物的亲和力、酶的热稳定性和pH耐受性等。若能通过上述手段获得耐高温普鲁兰酶高产重组菌株,将会对食品工业及相关领域产生深远的影响。
[1]Kunamneni A,Singh S.Improved high thermal stability of pullulanase from a newly isolated thermophilic Bacillus sp.AN-7[J].Enzyme Microb Technol,2006,39(7):1399-1404.
[2]Singh R S,Saini G K,Kennedy J F.Covalent immobilization and thermodynamic characterization of pullulanase for the hydrolysis of pullulan in batch system[J].Carbohydr Polym,2010,81(2):252-259.
[3]Kitahata S,Tanimoto T,Ikuta A,et al.Synthesis of novel heterobranched beta-cyclodextrins from 42-O-beta-D-galactosylmaltose and beta-cyclodextrin by the reverse action of pullulanase,and isolation and characterization of the products[J].Biosci Biotechnol Biochem,2000,64(6):1223-1229.
[4]Domań-Pytka M,Bardowski J.Pullulan degrading enzymes of bacterial origin[J].Crit Rev Microbiol,2004,30(2):107-121.
[5]Nair S,Singhal R,Kamat M.Induction of pullulanase production in Bacillus cereus FDTA-13[J].Biores Technol,2007,98(4):856-859.
[6]Turkenburg J P,Brzozowski A M,Svendsen A,et al.Structure of a pullulanase from Bacillus acidopullulyticus[J].Proteins,2009,76(2):516-519.
[7]Mikami B,Iwamoto H,Malle D,et al.Crystal structure of pullulanase:evidence for parallel binding of oligosaccharides in the active site[J].J Mol Biol,2006,359(3):690-707.
[8]周念波,孙杰,王晶,等.普鲁兰酶在食品工业中的应用[J].食品工程,2008(2):18-20.
[9]Norman B E,Bagsvaerd.A novel debranching enzyme for application in the glucose syrup industry[J].Stárch:Starke,1982,34:340-346.
[10]Castro G R,Santopietro L M D,Sineriz F.Acid pullulanase from Bacillus polymyxa MIC-23[J].Appl Biochem Biotechnol,1992,37:227-233.
[11]Wu C,Colleoni C,Myers A M,et al.Enzymatic properties and regulation of ZPU1,the maize pullulanase-type starch debranching enzyme[J].Arch Biochem Biophys,2002,406(1):21-32.
[12]Wallenfels K,Bender H,Rached J.Pullulanase from Aerobacter aerogenes;production in a cell-bound state.Purification and properties of the enzyme[J].Biochem Biophys Res Commun,1966,22(3):254-261.
[13]Takasaki Y.Productions and utilizations of beta-amylase and pullulanase from Bacillus cereus var.mycoides[J].Agric Biol Chem,1976,40(8):1515-1522.
[14]SChülein M,H jer-Pedersen B.Characterization of a new class of thermophilic pullulanases from Bacillus acidopullulyticus[J].Ann New York Acad Sci,1984,434(1):271-274.
[15]Matuschek M,Burchhardt G,Sahm K,et al.Pullulanase of Thermoanaerobacterium thermosulfurigenesEM1(Clostridium thermosulfurogenes):molecular analysis of the gene,composite structure of the enzyme,and a common model for its attachment to the cell surface[J].J Bacteriol,1994,176(11):3295.
[16]Takasaki Y.Pullulanase-Amylase complex enzyme from Bacillus subtilis[J].Agric Biol Chem,1987,51(1):9-16.
[17]Plant A R,Morgan H W,Daniel R M.A highly stable pullulanase from Thermus aquaticus YT-1[J].Enzyme Microb Technol,1986,8:668-672.
[18]Suzuki Y,Hatagaki K,Oda H.A hyperthermostable pullulanase produced by an extreme thermophile,Bacillus flavocaldarius KP 1228,and evidence for the proline theory of increasing protein thermostability[J].Appl Microbiol Biotechnol,1991,34(6):707-714.
[19]Duffner F,Bertoldo C,Andersen J T,et al.A new Thermoactive Pullulanase from desulfurococcus mucosus cloning,sequencing,purification,and characterization of the recombinant enzyme after expression in Bacillus subtilis[J].J Bacteriol,2000,182(22):6331-6338.
[20]Albertson G D,McHale R H,Gibbs M D,et al.Cloning and sequence of a type I pullulanase from an extremely thermophilic anaerobic bacterium,Caldicellulosiruptor saccharolyticus[J].Biochim Biophys Acta,1997,1354(1):35-39.
[21]Bertoldo C,Armbrecht M,Becker F,et al.Cloning,sequencing,and characterization ofa heat-and Alkali-Stable type I pullulanase from Anaerobranca gottschalkii[J].Appl Environ Microbiol,2004,70(6):3407-3416.
[22]Zouari Ayadi D,Ben Ali M,Jemli S,et al.Heterologous expression,secretion and characterization of the Geobacillus thermoleovorans US105 type I pullulanase[J].Appl Microbiol Biotechnol,2008,78(3):473-481.
[23]Gomes I.Highly thermostable amylase and pullulanase of the extreme thermophilic eubacterium Rhodothermus marinus:production and partial characterization[J].Biores Technol,2003,90(2):207-214.
[24]Kriegshauser G,Liebl W.Pullulanase from the hyperthermophilic bacterium Thermotoga maritima:purification by beta-cyclodextrin affinity chromatography[J].J Chromatogr B:Biomed Sci Appl,2000,737(1/2):245-251.
[25]Bertoldo C,Duffner F,Jorgensen P L,et al.Pullulanase type I from Fervidobacterium pennavorans Ven5:cloning,sequencing,and expression of the gene and biochemical characterization of the recombinant enzyme[J].Appl Environ Microbiol,1999,65(5):2084-2091.
[26]Francetic O,Pugsley AP.Towards the identification of type II secretion signals in a nonacylated variant of pullulanase from Klebsiella oxytoca[J].J Bacteriol,2005,187(20):7045-7055.
[27]Kuriki T,Okada S,Imanaka T.New type of pullulanase from Bacillus stearothermophilus and molecular cloning and expression of the gene in Bacillus subtilis[J].J Bacteriol,1988,170(4):1554-1559.
[28]Janse B J,Pretorius I S.Expression of the Klebsiella pneumoniae pullulanase-encoding gene in Saccharomyces cerevisiae[J].Curr Genet,1993,24(1/2):32-37.
[29]Xu B,Yang Y J,Huang Z X.Cloning and overexpression of gene encoding the pullulanase from Bacillus naganoensis in Pichia pastoris[J].J Microbiol Biotechnol,2006,16(8):1185-1191.
[30]Ben Messaoud E,Ben Ammar Y,Mellouli L,et al.Thermostable pullulanase type I from new isolated Bacillus thermoleovorans US105:cloning,sequencing and expression of the gene in E.coli[J].Enzyme Microb Technol,2002,31(6):827-832.
[31]Takizawa N,Murooka Y.Cloning of the pullulanase gene and overproduction of pullulanase in Escherichia coli and Klebsiella aerogenes[J].Appl Microbiol Biotechnol,1985,49(2):294-298.
[32]Duffner F,Bertoldo C,Andersen J T,et al.A new thermoactive pullulanase from Desulfurococcus mucosus:cloning,sequencing,purification,and characterization of the recombinant enzyme after expression in Bacillus subtilis[J].J Bacteriol,2000,182(22):6331.
[33]Kurik T,Park IJH,OkadaS,etal.Purification and characterization ofThermostable Pullulanase from Bacillus stearothermophilus and molecular cloning and expression of the gene in Bacillus subtilis[J].Appl Environ Microbiol,1988,54(11):2881-2883.
[34]Martin W T,Phillip J B,Larry N A,et al.Pullulanase expression constructs containing α-amylase promoter and leader sequences:US,6300115B1[P].2001-10-09.
[35]Dennis M.Katkocin N W Z,Yang S S.Novel thermostable pullulanase enzyme and method for its production:US,4628028[P].1986-12-09.
[36]Philippe Deweer A A.Pullulanase producing microrganisms:US,5817498[P].1998-10-06.
[37]Michaelis S,Chapon C,D'Enfert C,et al.Characterization and expression of the structural gene for pullulanase,a maltoseinducible secreted protein of Klebsiella pneumoniae[J].J Bacteriol,1985,164(2):633-638.
[38]Choi C H,Kim S H,Jang J H,et al.Enzymatic synthesis of glycosylated puerarin using maltogenic amylase from Bacillus stearothermophilus expressed in Bacillus subtilis[J].J Sci Food Agric,2010,90(7):1179-1184.
[39]彭清忠,张惟材,朱厚础.枯草杆菌表达系统的研究进展[J].生物技术通讯.2001,12(3):220-225.
[40]Ohdan K,Kuriki T,Kaneko H,et al.Characteristics of two forms of α-amylase and structural implication [J].Appl Environ Microbiol,1999,65(10):4652-4658.
[41]Svensson B.Protein engineering in the α-amylase family:catalytic mechanism,substrate specificity,and stability[J].Plant Mole Biolo,1994,25(2):141-157.
[42]Janecek S,Svensson B,Henrissat B.Domain evolution in the αamylase family[J].J Mol Evolution,1997,45(3):322-331.
[43]Jespersen H M,MacGregor E A,Sierks M R,et al.Comparison of the domain-level organization of starch hydrolases and related enzymes[J].Biochem J,1991,280(Pt 1):51-55.
[44]Renz A,Schikora S,Schmid R,et al.cDNA sequence and heterologousexpression ofmonomeric spinach pullulanase:multiple isomeric forms arise from the same polypeptide[J].Biochem J,1998,331(Pt 3):937-945.
[45]Ryter A,Pugsley A.Cloning and expression in Escherichia coli of the Klebsiella pneumoniae genes for production,surface localization and secretion of the lipoprotein pullulanase[J].EMBO J,1987,6(11):3531-3538.
[46]Kashiwabara S,Ogawa S,Miyoshi N,et al.Three domains comprised in thermostable molecular weight 54,000 pullulanase of type I from Bacillus flavocaldarius KP1228 [J].Biosci,Biotechnol,Biochem,1999,63(10):1736-1748.
[47]Liebl W,Stemplinger I,Ruile P.Properties and gene structure of the Thermotoga maritima alpha-amylase AmyA,aputative lipoprotein of a hyperthermophilic bacterium[J].J Bacteriol,1997,179(3):941-948.
[48]Kuriki T,Imanaka T.The concept of the[alpha]-amylase family:structural similarity and common catalytic mechanism[J].J Biosci Bioeng,1999,87(5):557-565.
[49]Kuriki T,Imanaka T.Nucleotide sequence of the neopullulanase gene from Bacillus stearothermophilus[J].Microbiology,1989,135(6):1521-1528.
[50]Yamashita M,Matsumoto D,Murooka Y.Amino acid residues specific for the catalytic action towards α-1,6-glucosidic linkages in Klebsiella pullulanase[J].J Ferment Bioeng,1997,84(4):283-290.
[51]Bertoldo C,Antranikian G.Starch-hydrolyzing enzymes from thermophilic archaea and bacteria[J].Curr Opin Chem Biol,2002,6(2):151-160.
[52]SuzukiY,FujiiH,Uemura H,etal.Purification and characterization of extremely thermostable exo-oligo-1,6-glucosidase from a caldoactive Bacillus sp.KP 1228[J].Starch:Stärke,1987,39(1):17-23.
[53]MatthewsB,Nicholson H,BecktelW.Enhanced protein thermostability from site-directed mutations that decrease the entropy of unfolding[J].PNAS,1987,84(19):6663-6667.
[54]Suzuki Y.A general principle of increasing protein thermostability[J].Proc Jpn Acad Ser B,1989,65(6):146-148.
[55]Menéndez-AriasL,ArgosfP.Engineering protein thermal stability:Sequence statistics point to residue substitutions in αhelices[J].J Mol Biol,1989,206(2):397-406.
[56]Kusano S,Takahashi S,Fujimoto D,et al.Effects of reduced maltooligosaccharides on the thermal stability of pullulanase from Bacillus acidopullulyticus [J].Carbohydr Res,1990,199(1):83-89.
[57]Kuroiwa T,Shoda H,Ichikawa S,et al.Immobilization and stabilization of pullulanase from Klebsiella pneumoniae by a multipoint attachment method using activated agar gel supports[J].Process Biochem,2005,40(8):2637-2642.
[58]SchlossP D,Handelsman J.Metagenomics for studying unculturable microorganisms:cutting the Gordian knot[J].Genome Biol,2005,6(8):229.doi:10.1186/qb-2005-6-8-229.
[59]Schmeisser C,Steele H,Streit W R.Metagenomics,biotechnology with non-culturable microbes[J].Appl Microbiol Biotechnol,2007,75(5):955-962.
猜你喜欢
食品与发酵工业(2021年16期)2021-08-31
食品与药品(2020年1期)2020-03-10
现代食品科技(2018年10期)2018-11-06
中国塑料(2016年7期)2016-04-16
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
合成材料老化与应用(2015年4期)2015-07-25
应用化工(2014年1期)2014-08-16
原子与分子物理学报(2014年3期)2014-06-06
食品工业科技(2014年13期)2014-03-11
中国粮油学报(2014年8期)2014-02-06
