
禽流感血凝素HA超表达载体的构建及高表达百脉根的获得
2014-03-26 08:38张伟伟袁蓓张占路赵杨敏徐秉良吴燕民
草业学报 2014年4期
张伟伟,袁蓓,张占路,赵杨敏,徐秉良,吴燕民*
(1.甘肃农业大学草业学院,甘肃 兰州730070;2.中国农业科学院生物技术研究所,北京100081;3.深圳市农科集团有限公司,广东 深圳518040)
自2003年到2013年,高致病性禽流感H5N1共引起人患病630例,致死375人,大规模捕杀家禽及候鸟数量更是数以亿计,给世界各地造成严重的经济损失和巨大的生命安全威胁。早在1992年,Mason等[1]提出了“植物可食疫苗”的概念,成功利用烟草(Nicotianatabacum)表达了乙肝表面抗原(HBsAg)基因。近十年已有大量的抗原被植物表达,包括抵御 HIV的2G12抗体[2]、乙肝表面抗原[3-5]、霍乱毒素B亚基[6]、大肠杆菌热敏毒素B亚基[7-8]、狂犬病毒G蛋白[9]等,但是目前禽流感植物疫苗的报道还是较少且抗原蛋白表达量低限制了植物疫苗进一步的发展[10]。
大量研究表明,影响重组药用蛋白在植物中表达量的因素包括很多方面。一是转录水平,启动子的选择是提高外源基因表达水平的关键。常用的启动子分为组成型启动子、组织特异性启动子、诱导型启动子和人工合成启动子。前3种启动子都可以实现外源基因的表达与调控,但人工合成启动子能够进一步提高外源基因的表达水平。如重复CaMV35启动子的增强子序列,构建双35S启动子,可显著提高外源基因的表达水平[11-12]。终止子通过终止转录和对mRNA 3′端的加工调节转录水平,进而影响外源基因在植物中的表达水平。Nagaya等[13]分别比较了不同终止子,发现热击蛋白18.2终止子能有效提高外源基因表达水平。二是翻译水平,研究表明通过增强mRNA的稳定性[14-15],按照植物偏爱密码子进行密码子优化可有效提高重组蛋白的表达水平[16]。三是翻译后水平,重组蛋白合成后,利用不同定位信号可以定位于不同的亚细胞结构中,亚细胞结构的生理生化环境、存储空间等多种因素都影响着蛋白的积累水平。为外源蛋白选择合适的分拣部位,有助于其高浓度富集。例如在重组蛋白C-添加内质网滞留信号序列-KDEL,可以增加外源蛋白在内质网中的积累水平[17]。郑琳琳等[18]证明可通过构建超表达载体有效提高外源基因的表达水平。
本研究选择豆科全价饲料牧草百脉根(Lotuscorniculatus)[19]作为受体植物,在已建立的,以磷酸甘露糖异构酶基因(PMI)为筛选标记的,安全高效的百脉根遗传转化体系的基础上,利用已获得的禽流感血凝素(HA)和大肠杆菌不耐热毒素B亚基(LTB)融合基因,通过添加强启动子,增强子Ω序列、KDEL序列及加长poly(A)序列等多种调控元件,构建HA-LTB高效融合植物表达载体,以期提高HA抗原蛋白表达量,建立外源融合抗原蛋白高水平表达平台,为禽流感可饲疫苗的研制奠定基础。
1 材料与方法
1.1 材料
1.1.1 植物材料 百脉根:栽培品种里奥(L.corniculatuscv.Leo)由中国农业科学院生物技术研究所作物与分子实验室提供。
1.1.2 菌种及载体 大肠杆菌DH5α、根癌农杆菌LBA4404由本实验室保存,pCAMBIA2301、pMD19-T商业载体购于 Takara生物公司,pMD-2E+35S、pMD-Ω由本实验室保存,pMD-Nos-poly(A)由本实验室设计经金为智生物公司合成,pCAMBIA2301-PMI-HA-LTB-MARs植物融合表达载体由本实验室保存。
1.1.3 工具酶和主要生物学试剂EcoRⅠ、HindⅢ、MluⅠ等限制性内切酶、T4DNA连接酶、Ex Taq Polymerase、LA Taq Polymerase购自Takara生物公司;DNA Marker购自北京全式金生物技术有限公司(TransGen Biotech);高纯质粒小量制备试剂盒、通用植物总RNA提取试剂盒购自百泰克生物公司(Bioteke);反转录试剂盒购于Invitrogen公司;Gel/PCR Extraction kit购自Biomiga生物公司;大肠杆菌热敏毒素B亚基酶联免疫分析试剂盒购自R&D system。
1.1.4 培养基 B5培养基(phytotech B5media)、MS培养基(Murashige Skoog medium)购自美国 Phyto Technology生物技术公司,YEB培养基(agrobacterium rhizogene medium)、LB培养基(Luria-Bertani bacterial medium)、植物凝胶(phytagel)、甘露糖购自北京拜尔迪生物公司(Biodee),1mg/mL的BSA标准品(Bovine Serum Albumin Standard Ampules)购自GeneStar公司,其他试剂均为国产分析纯。
1.1.5 引物与测序 利用Primer 5.0引物设计软件,根据相关基因序列及需要添加酶切位点序列、基因片段序列设计、载体构建过程中所有测序工作,由北京奥科生物技术有限公司完成,引物序列见表1。
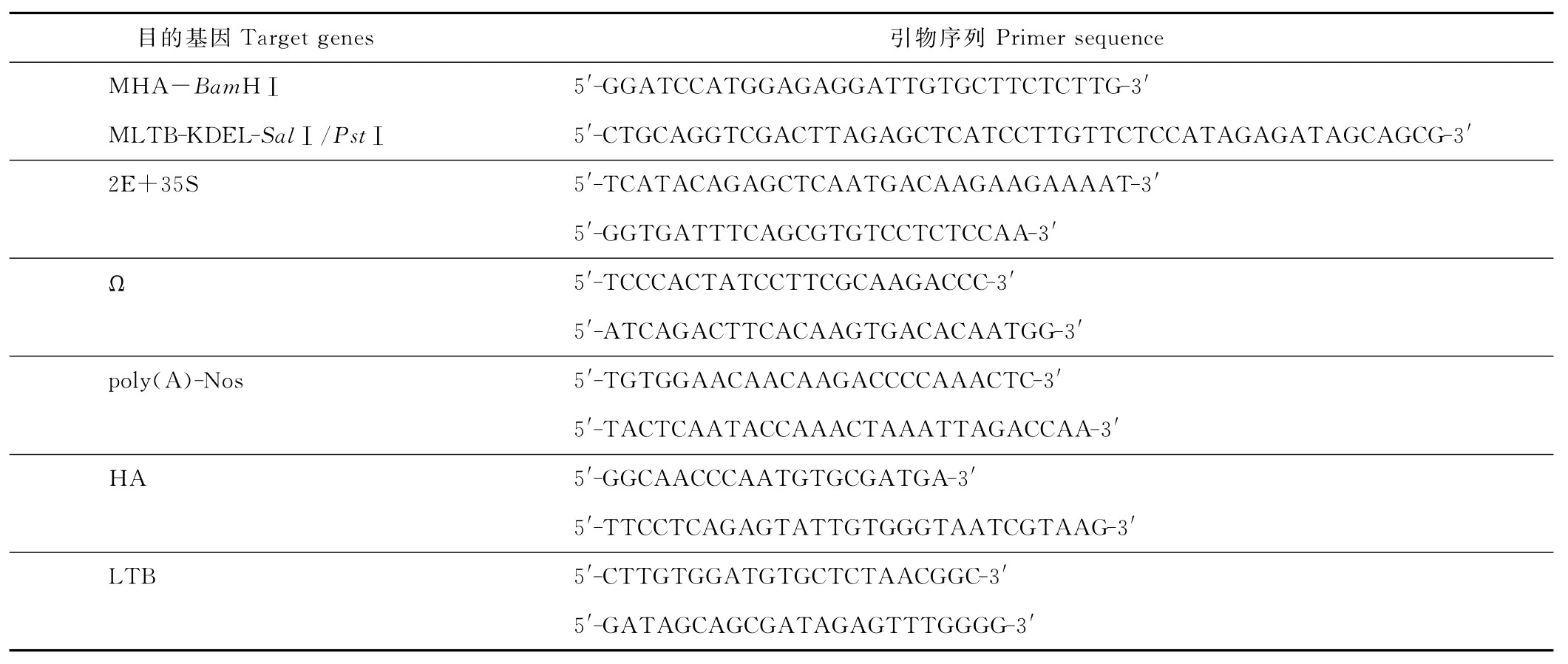
表1 目的基因的引物序列Table 1 Primer sequence of target genes
1.2 试验方法
1.2.1 高效植物表达载体的设计和构建 在已有载体pCAMBIA2301-PMI-MARs和HA-LTB融合基因的基础上,首先通过引物设计在LTB后添加KDEL序列,并利用HA-BamHⅠ引物和LTB-KDEL-SalⅠ/PstⅠ引物进行PCR扩增,回收产物及载体,经BamHⅠ和PstⅠ酶切后通过T4连接酶连接,连接产物转化大肠杆菌,经PCR及酶切验证;将载体中的CaMV35S启动子置换为2E+35S启动子,载体和pMD-2E+35S都经XbaI和HindⅢ双酶切,T4连接酶连接后转化大肠杆菌,经PCR及酶切验证;然后添加Ω增强子序列,载体和pMD-Ω经BamHⅠ和HindⅢ双酶切,T4连接酶连接后转化大肠杆菌,经PCR验证;最后添加长度为100bp的poly(A)与NOS串联序列。载体和pMD-Nos-poly(A)经KpnⅠ和SphⅠ双酶切,分别回收载体大片段和Nos-poly(A)片段,T4连接酶连接后转化大肠杆菌,经PCR及酶切验证。对验证结果正确的质粒进行测序,获得HA-LTB融合植物表达载体如图1。

图1 融合植物表达载体HA-LTB结构示意图Fig.1 Flow chart of the construction of HA-LTB vector
1.2.2 百脉根的遗传转化 将携带目的载体的农杆菌LBA4404涂板,挑取单菌落,28℃摇床振荡培养以活化菌体,使细菌达到对数生长期(OD600为0.5~0.6)。8000r/min离心10min,菌体用 MS液体培养基重悬,使OD600为 0.5~0.6[20]。
取播种后约8d的百脉根幼苗,以百脉根子叶作为外植体在子叶柄部位剪切,放入培养皿中。将混均匀的菌液倒入放有子叶的培养皿中,浸泡30min。然后将其中的子叶转入有滤纸的MS培养基上,25℃暗培养3~4d,将暗培养后的愈伤组织转接到加抗生素的筛选培养基中培养约1周。转移到MS+Cef 100mg/L的生根培养基上。每两周换一次新的培养基,快速生根,直至成苗[21]。
1.2.3 转基因百脉根的分子检测 再生植株PCR分子检测,根据HA和LTB基因全长序列设计特异引物(表1),以再生百脉根植株基因组为模板,按以下体系进行PCR反应:在20μL PCR反应体系中加入12.75μL dd H2O,2μL 10×buffer,2μL dNTP,基因上下游引物各1μL,ExTaq酶0.25μL,1μL模板;反应程序为:94℃预变性5min,然后94℃变性45s,58℃退火45s,72℃延伸1min,30个循环,最后72℃延伸10min。
转基因植株的RT-PCR检测,参照“通用植物总RNA提取试剂盒”说明进行百脉根RNA的提取:按照Invitrogen公司反转录试剂盒说明书进行反转录及PCR反应。
转基因植株的ELISA检测,百脉根总可溶性蛋白的提取:超净台内取100mg转基因百脉根叶片放入2.0 mL Ep管中,迅速置于液氮内,打样机上破碎,加入500μL蛋白提取液,室温12000r/min离心10min,取上清,即为百脉根总可溶性蛋白粗提液,用于考马斯亮蓝蛋白定量;LTB蛋白的定量依照大肠杆菌热敏毒素B亚基酶联免疫试剂盒说明书操作。
2 结果与分析
2.1 高效植物表达载体的构建
2.1.1 添加KDEL序列 在已有载体pCAMBIA2301-PMI-MARs和HA-LTB融合基因的基础上,通过引物设计在LTB后添加KDEL序列。利用HA-BamHⅠ引物和LTB-KDEL-SalⅠ/PstⅠ引物(表1)以构建载体质粒为模板进行PCR验证,得到1条长约2000bp的条带,采用BamHⅠ和PstⅠ对构建好的植物表达载体进行双酶切鉴定,琼脂糖凝胶电泳得到1条约2000bp的条带。PCR及酶切验证表明插入基因片段与HA-LTBKDEL序列2062bp大小相符,且测序结果与HA-LTB-KDEL序列也完全匹配,载体已正确插入KDEL序列。
2.1.2 置换启动子 将载体pCAMBIA2301中的CaMV35S启动子置换为2E+35S启动子。已构建载体经XbaI和HindⅢ双酶切,经PCR及酶切验证,结果表明PCR及酶切获得1条长约700bp的条带,与785bp的2E+35S启动子大小相符,将验证结果正确的质粒进行测序,且测序结果与2E+35S启动子序列也完全匹配,载体已正确插入2E+35S启动子序列。
2.1.3 添加Ω增强子序列 在已有载体pCAMBIA2301-PMI-MARs基础上添加Ω增强子序列,经PCR及酶切验证,结果表明PCR获得1条长约100bp的条带与75bp的Ω增强子序列大小相符,将验证结果正确的质粒进行测序,且测序结果与Ω增强子序列也完全匹配,载体已正确添加Ω增强子序列。
2.1.4 添加多聚poly(A)序列 已构建载体经KpnⅠ和SphⅠ双酶切,经PCR及酶切验证,结果表明PCR及酶切获得1条长约500bp的条带与poly(A)-Nos大小相符,将验证结果正确的质粒进行测序,且测序结果与poly(A)-Nos序列也完全匹配,载体已正确插入poly(A)-Nos序列。
通过对已构建载体进行PCR、双酶切鉴定及测序验证,结果表明载体已正确插入MHA-MLTB-KDEL序列、2E+35S启动子、Ω增强子序列、poly(A)-Nos序列,证明HA-LTB融合表达载体构建成功。
2.2 百脉根的遗传转化
参照实验室已建立的以磷酸甘露糖为筛选标记的百脉根安全高效遗传转化体系,以百脉根七日龄苗子叶(含下胚轴)为外植体,农杆菌介导法转化百脉根,经暗培养,分化培养和选择培养,最终获得抗性植株91株(图2)。
2.3 转基因百脉根的分子检测
2.3.1 PCR检测 获得百脉根抗性植株后,提取其基因组DNA,分别进行HA及LTB基因PCR检测,结果显示共获得21株转基因阳性百脉根植株,转化率达到23.1%,检测结果分别如图3-1和图3-2。

图2 百脉根的遗传转化Fig.2 Genetic transformation of L.corniculatus

图3 转基因百脉根PCR检测Fig.3 PCR assay of transgenic L.corniculatus
2.3.2 RT-PCR检测 为验证PCR阳性百脉根植株中HA及LTB基因在转录水平的表达,提取转基因植株总RNA,按照Invitrogen公司反转录试剂盒说明书进行反转录,并对cDNA进行PCR检测,结果如图4和5,说明HA及LTB基因在mRNA水平成功表达。
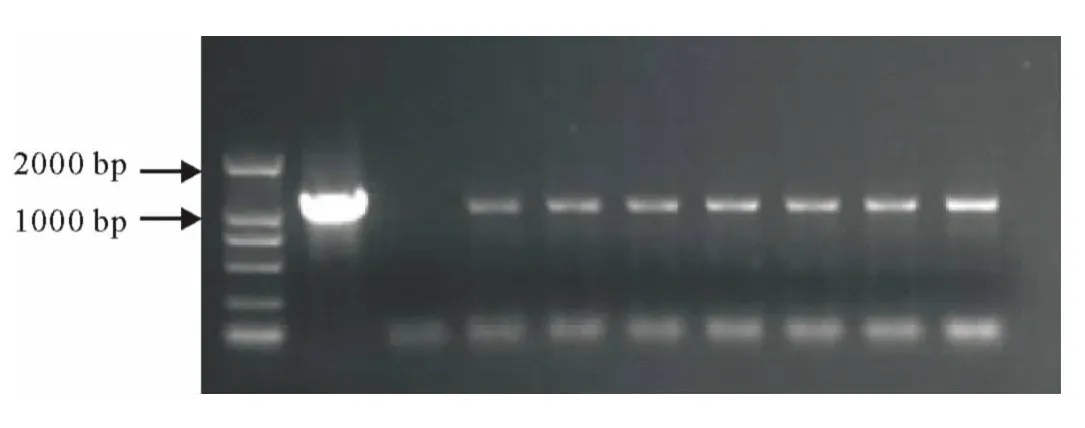
图4 HA基因RT-PCR检测Fig.4 RT-PCR of HA gene
图5 LTB基因RT-PCR检测Fig.5 RT-PCR of LTB gene
2.3.3 ELISA检测 将浓度分别为0.1mg/mL的牛血清白蛋白标准品(BSA)进行倍比稀释,在595nm吸光值下绘制BSA蛋白标准曲线。利用蛋白提取液提取转基因百脉根植株的总可溶性蛋白,经考马斯亮蓝染色后,测定其在595nm下的吸光值,并带入BSA蛋白标准曲线,计算转基因百脉根中总可溶性蛋白浓度。采用蛋白定量中同样的方法,将LTB标准品进行倍比稀释,进行LTB酶联免疫反应,绘制LTB标准曲线(图6),计算LTB抗原蛋白浓度。最终计算出21株阳性百脉根总可溶性蛋白中LTB抗原蛋白的含量(图7),其中12号百脉根LTB表达量最高,达到0.086μg/mg(蛋白粗提液),结果表明LTB与HA蛋白融合不影响各自蛋白的免疫活性,LTB表达量最高达到0.086μg/mg(蛋白粗提液),HA抗原蛋白的分子量是LTB的3倍,由此可以推算出HA在百脉根中的最高表达量约为0.25μg/mg(蛋白粗提液)。
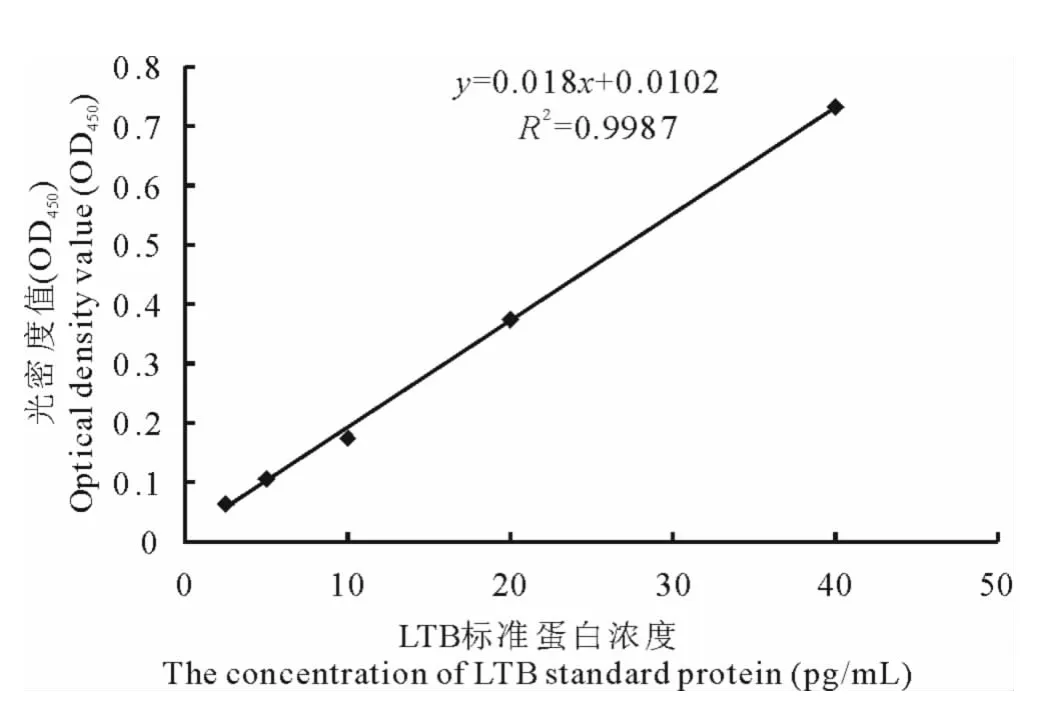
图6 LTB蛋白标准曲线Fig.6 Standard curve of LTB protein
图7 转基因百脉根中LTB蛋白含量Fig.7 LTB content in transgenic L.corniculatus
3 讨论
选择一个最佳启动子高表达整合到植物中的外源基因是构建高效植物表达载体的一项重要工作。CaMV35S启动子因其强表达和组织特异性表达特点而被广泛应用于双子叶转基因植物。在CaMV35S启动子的-46至-105序列中含有增强子序列,将CaMV35S启动子的-46至-105序列中含有增强子元件重复串联,构建双35S启动子,可显著提高外源基因的表达水平[11-12]。Wally等[22]研究结果表明D35s启动子的表达量相比于CaMV35S启动子平均提高了3~10倍,进一步表明添加双35S启动子有助于提高外源基因及外源蛋白表达量。
真核基因的5′-3′-非翻译序列(UTR)对保持mRNA的稳定性和翻译水平有重要的作用。Gallie等[23]研究结果表明在5′端添加Ω元件能提高基因的翻译效率。朱祯[24]报道烟草花叶病毒(TMV)的126ku蛋白基因的Ω因子(63bp)的存在可以使Gus RNA的转译活性提高数十倍。毛立群和郭三堆[25]将pBI121GUS基因的3′端插入了长度分别为25,50及100个dA残基的poly(A)序列并转入烟草中,发现GUS基因的活性分别提高了27,37和47倍,表明poly(A)序列的长短与翻译水平呈相关。Shapiro等[26]和Gallie等[23]分别报道称poly(A)尾巴对转录后翻译起增强作用,且增强的程度与poly(A)尾巴的长度正相关。因此,添加5′-3′-非翻译序列在一定程度解决外源基因在植物体内表达量低和稳定性差的问题。
引导肽可以利用分泌途径将外源蛋白引向特定的部位,从而提高重组蛋白的积累水平。外源基因的表达产物会受细胞内的蛋白酶的降解,而在接入引导肽序列则会及时对蛋白进行定位,还可以提高蛋白的含量。Wandelt等[27]将编码豌豆(Pisumsativum)种子VICILIN蛋白的基因和内质网定位信号KDEL的编码序列连接转化植物,发现在转基因烟草和紫花苜蓿(Medicagosativa)中带有KDEL序列的蛋白含量分别是对照的20和100倍。Sojikul等[28]及Thanavala等[29]均证明在重组蛋白C-添加内质网滞留信号序列KDEL,可以增加外源蛋白在内质网中的积累水平,从而提高外源蛋白在转基因植株中的表达量。因此构建含KDEL序列的表达载体可能对提高外源蛋白的稳定性和积累水平有一定作用。
为提高禽流感血凝素表达量,本研究利用已有的HA-LTB融合基因,通过设计和添加多种调控元件,即双增强子CaMV35S启动子、Ω序列、加长poly(A)序列、内质网滞留信号KDEL序列等,经PCR、双酶切及测序鉴定,证明HA-LTB融合基因高效植物表达载体构建成功。利用农杆菌介导方法转化百脉根,获得抗性植株91株,经PCR,RT-PCR检测,得到转基因阳性植株21株。通过夹心ELISA检测,结果表明LTB最高表达量达到0.086 μg/mg(蛋白粗提液),HA最高表达量达到0.25μg/mg(蛋白粗提液),即占总可溶性蛋白的0.025%,显著高于Guo等[30]于2012年报道的HA抗原蛋白表达量为总可溶性蛋白的0.00786%,已接近国际先进水平。本研究通过载体的设计和构建大幅度提高了禽流感血凝素在百脉根中的表达量,实现了外源抗原蛋白在豆科牧草百脉根中的高水平表达,为动物实验及禽流感可饲用疫苗的研究奠定了基础。
[1] Mason H S,Lam D M,Arntzen C J.Expression of hepatitis B surface antigen in transgenic plants[J].Proceedings of the National Academy of Sciences,1992,89(24):11745-11749.
[2] Holland T,Sack M,Rademacher T,etal.Optimal nitrogen supply as a key to increased and sustained production of a monoclonal full-size antibody in BY-2suspension culture[J].Biotechnology and Bioengineering,2010,107(2):278-289.
[3] Unni S C,Soniya E V.TransgenicCucumissativusexpressing the hepatitis B surface antigen[J].Plant Molecular Biology Reporter,2010,28(4):627-634.
[4] He Z M,Jiang X L,Qi Y,etal.Assessment of the utility of the tomato fruit-specific E8promoter for driving vaccine antigen expression[J].Genetica,2008,133(2):207-214.
[5] Sunil Kumar G B,Ganapathi T R,Srinivas L,etal.Expression of hepatitis B surface antigen in potato hairy roots[J].Plant Science,2006,170(5):918-925.
[6] Jani D,Meena L S,Rizwan-ul-Haq Q M,etal.Expression of cholera toxin B subunit in transgenic tomato plants[J].Transgenic Research,2002,11(5):447-454.
[7] Moravec T,Schmidt M A,Herman E M,etal.Production ofEscherichiacoliheat labile toxin(LT)B subunit in soybean seed and analysis of its immunogenicity as an oral vaccine[J].Vaccine,2007,25(9):1647-1657.
[8] Rosales-Mendoza S,Soria-Guerra R E,López-Revilla R,etal.Ingestion of transgenic carrots expressing theEscherichiacoliheat-labile enterotoxin B subunit protects mice against cholera toxin challenge[J].Plant Cell Reports,2008,27(1):79-84.
[9] Ashraf S,Singh P K,Yadav D K,etal.High level expression of surface glycoprotein of rabies virus in tobacco leaves and its immunoprotective activity in mice[J].Journal of Biotechnology,2005,119(1):1-14.
[10] 袁蓓,李京生,吴燕民.利用植物生产药用蛋白的进展、局限及应对策略[J].草业学报,2013,22(4):283-299.
[11] Hull A K,Criscuolo C J,Mett V,etal.Human-derived,plant-produced monoclonal antibody for the treatment of anthrax[J].Vac-cine,2005,23(17):2082-2086.
[12] Ruggiero F,Exposito J Y,Bournat P,etal.Triple helix assembly and processing of human collagen produced in transgenic tobacco plants[J].Febs Letters,2000,469:132-136.
[13] Nagaya S,Kawamura K,Shinmyo A,etal.The HSP terminator ofArabidopsisthalianaincreases gene expression in plant cells[J].Plant and Cell Physiology,2010,51:328-332.
[14] Anthonisen I L,Salvador M L,Klein U.Specific sequence elements in the 5′untranslated regions of rbcL and atpB gene mRNas stabilize transcripts in the chloroplast ofChlamydomonasreinhardtii[J].RNA,2001,7(7):1024-1033.
[15] Huang L F,Liu Y K,Lu C A,etal.Production of human serum albumin by sugar starvation induced promoter and rice cell culture[J].Transgenic Research,2005,14:569-581.
[16] Kang T J,Kim B G,Yang J Y,etal.Expression of a synthetic cholera toxin B subunit in tobacco using ubiquitin promoter and bar gene as a selectable marker[J].Molecular Biotechnology,2006,32:93-100.
[17] Ko K,Tekoah Y,Rudd P M,etal.Function and glycosylation of plant-derived antiviral monoclonal antibody[J].Proceedings of the National Academy of Sciences of the United States of America,2003,100:8013-8018.
[18] 郑琳琳,王佳,贺龙梅,等.唐古特白刺蛋白激酶基因NtCIPK2超表达载体构建及紫花苜蓿转化研究[J].草业学报,2013,22(6):223-229.
[19] 马江涛,王宗礼,黄东光,等.基因工程在牧草培育中的应用[J].草业学报,2010,19(6):248-262.
[20] 陶飞,马江涛,唐益雄,等.抗逆转录因子在百脉根中的功能分析[J].草业学报,2013,22(3):250-258.
[21] 刘建利,张占路,吴燕民,等.百脉根农杆菌快速高效遗传转化体系的建立[J].草业学报,2006,15(3):128-131.
[22] Wally O,Jayaraj J,PunjaZ K.Comparative expression ofβ-glucuronidase with five different promoters in transgenic carrot(DaucuscarotaL.)root and leaf tissues[J].Plant Cell Reports,2008,27(2):279-287.
[23] Gallie D R,Lucas W J,Walbot V.Visualizing mRNA expression in plant protoplasts:factors influencing efficient mRNA uptake and translation[J].The Plant Cell Online,1989,1(3):301-311.
[24] 朱祯.走向21世纪的植物分子生物学[M].北京:科学出版社,2000:121.
[25] 毛立群,郭三堆.Ω 序列和3′poly(dA)长度与基因表达效率的关系[J].植物学报,1998,40(12):1166-1168.
[26] Shapiro R A,Herrick D,Manrow R E,etal.Determinants of mRNA stability inDictyosteliumdiscoideumamoebae:differences in poly(A)tail length,ribosome loading,and mRNA size cannot account for the heterogeneity of mRNA decay rates[J].Molecular and Cellular Biology,1988,8(5):1957-1969.
[27] Wandelt C I,Khan M R I,Craig S,etal.Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants[J].The Plant Journal,1992,2(2):181-192.
[28] Sojikul P,Buehner N,Mason H S.A plant signal peptide-hepatitis B surface antigen fusion protein with enhanced stability and immunogenicity expressed in plant cells[J].Proceedings of the National Academy of Sciences,2003,100(5):2209-2214.
[29] Thanavala Y,Mahoney M,Pal S,etal.Immunogenicity in humans of an edible vaccine for hepatitis B[J].Proceedings of the National Academy of Sciences of the United States of America,2005,102(9):3378-3382.
[30] Guo Q Q,Zhang Z L,Jiang S J,etal.Expression of an avian influenza virus(H5N1)hemagglutinin gene in transgenicLotus corniculatus[J].Plant Molecular Biology Reporter,2012,30(5):1117-1124.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
山东农业科学(2021年8期)2021-09-12
中国生殖健康(2020年4期)2021-01-18
草业科学(2019年11期)2019-12-12
西藏农业科技(2019年3期)2019-11-04
科海故事博览·下旬刊(2019年6期)2019-04-16
中国生殖健康(2018年4期)2018-11-06
现代园艺(2018年3期)2018-02-10
海南师范大学学报(自然科学版)(2017年4期)2018-01-22
上海农业学报(2017年3期)2017-04-10
