
利用不同方法测定红松人工林叶面积指数的季节动态
2014-05-05 06:17王宝琦刘志理戚玉娇金光泽
生态学报 2014年8期
王宝琦,刘志理,戚玉娇,金光泽,*
(1.东北林业大学生态研究中心,哈尔滨 150040;2.东北林业大学林学院,哈尔滨 150040)
叶面积指数(LAI)指单位水平地面面积上总叶面积的一半[1],是量化植物冠层结构最常用参数之一,是森林生态系统中生态生理学模型的基本林冠结构参数[2],决定着众多植被林冠机能,如光合、呼吸、蒸腾作用及截留降雨[3-5]。LAI的季节变异是森林生态系统生产力、林下植被更新及其多样性的决定因素[6]。准确地测定LAI对于评估生态系统对气候变化、生物地球化学循环的响应至关重要[7]。
直接测定LAI的方法经典、成熟且相对精确,主要包括破坏性取样法、异速生长方程法、凋落物法[8]。破坏性取样法是将一定地面范围内的所有叶子取下并计算其叶面积,广泛应用于农作物及牧草LAI的测定,也应用于冠幅较小树木LAI的测定,但费时费力,且具有破坏性[9]。异速生长方程法是根据叶片生物量与胸径或边材面积的关系建立方程[10-12],结合不同树种的比叶面积(SLA)计算 LAI,结果相对精确,但也具有一定的破坏性,且不易获得LAI季节动态数据。凋落物法,避免了破坏性取样,多适用于测定落叶林型 LAI[13,9],近年来也有利用该方法测定常绿林 LAI的研究报道[2,14],但其结果的准确性还需要探讨。间接测定法通常是利用光学仪器在不破坏林木、不干扰冠层结构的前提下,简便、迅速地获取一些参数来反演LAI,且能提供LAI的季节动态数据而逐渐发展并被广泛应用。但光学仪器法测定LAI时忽略了林冠中存在的集聚效应,且不能有效区分叶片和木质部分,因此该方法测定的LAI被认为是有效LAI(Le)[15],其精度需要通过直接法测定值进行检验和校准。
关于直接法和间接法的对比研究报道较多。李轩然等[16]利用胸径与生物量的异速生长方程结合各树种的SLA,得到千烟洲针叶林LAI,且与光学仪器测定的LAI进行对比,结果表明光学仪器法测定值存在低估现象;马泽清等[17]利用异速生长方程及光学仪器法测定了几种南方人工林的LAI,得到光学仪器法测定值明显低于异速生长方程法;Jonckheere等[18]对比分析了异速生长方程与光学仪器法测定的LAI,结果证明前者测定的 LAI是后者的2倍;Nasahara等[19]结合展叶调查与凋落物法直接测定了日本中部寒温带落叶阔叶林LAI的季节动态,相对于该方法,光学仪器测定值最高低估程度达50%;Majasalmi等[12]通过光学仪器和异速生长方程法测定了欧洲赤松(Pinus sylvestris)和挪威云杉(Picea abies)等林型的LAI,并指出对光学仪器测定值进行针簇水平上集聚效应的校正后效果更佳。然而国内利用不同方法测定常绿针叶林LAI季节动态的研究尚少。
红松(Pinus koraiensis)人工林是我国东北东部山区主要人工林类型之一,本研究利用异速生长方程法、凋落物法、综合法(本研究将结合校正光学仪器值和凋落物测定LAI的方法称为综合法)及光学仪器法分别测定小兴安岭地区红松人工林LAI及其季节变化特征,以期为相对准确、有效地测定类似林型冠层LAI季节动态提供参考,并为该区域的森林生产力及生态进程模拟提供基础参数和科学依据。
1 材料和方法
1.1 研究区域概况
野外调查在黑龙江省凉水国家级自然保护区(47°10'50″N、128°53'20″E)进行,保护区位于黑龙江省伊春市带岭区,属小兴安岭南部达里带岭支脉的东坡。地形比较复杂,最高山脉海拔707.3 m。本区属于温带大陆性季风气候,年均气温-0.3℃,年均最高气温7.5℃,年均最低气温-6.6℃。年平均降水量676 mm,且多集中在7月份。积雪期130—150 d,无霜期 100—120 d。
1.2 样点设置
设置3个20 m×30 m的红松人工林固定样地(物种组成见表1)。每个样地中随机布设6个样点,各样点处布设1个凋落物收集器,凋落物收集器是用径粗8 mm的铁丝和尼龙网围成(孔径1 mm,深0.5—0.6 m),网口为正方形,面积为0.5 m2,凋落网底离地面0.5 m左右;各凋落物收集器旁边固定3根30 cm长度的PVC管,用于确定光学仪器采集数据的位置。
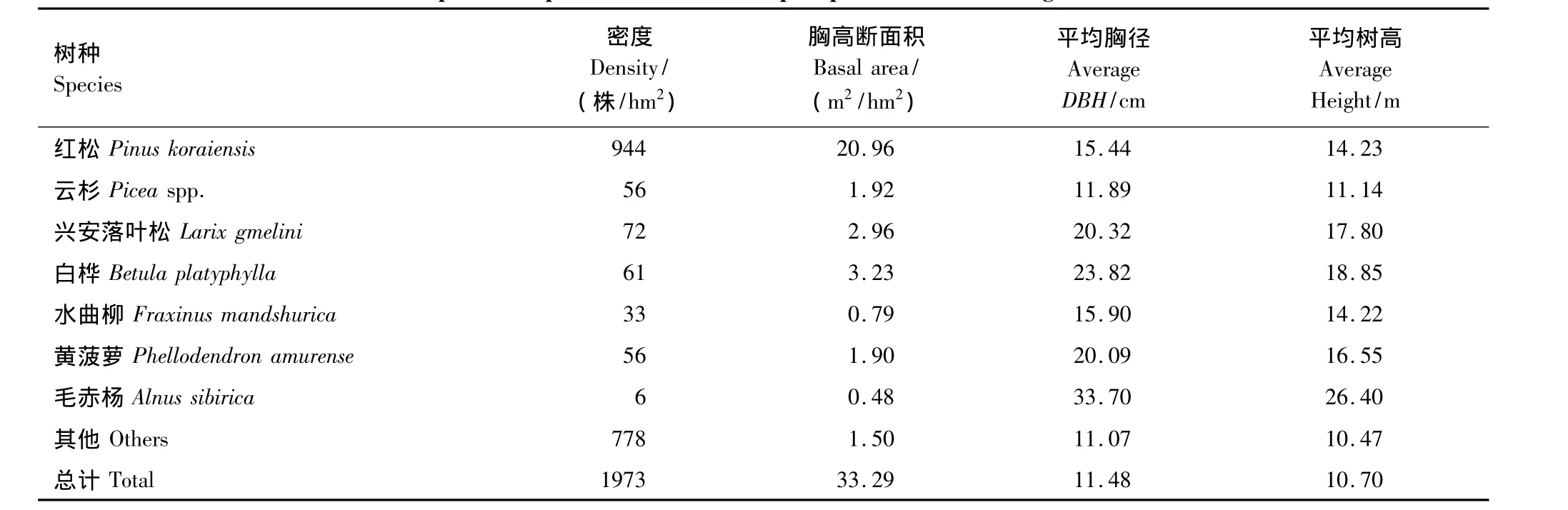
表1 小兴安岭红松人工林物种组成Table 1 Species composition of the Korean pine plantation in Xiaoxing'an Mountains
1.3 展叶调查
本研究中对红松,云杉(Picea spp.),兴安落叶松(Larix gmelini),白桦(Betula platyphylla)及水曲柳(Fraxinus mandshurica)五种主要树种的叶面积增长状况进行监测,监测日期为2012年5月1日、5月17日、5月24日、6月3日、6月11日、6月20日、7月4日、7月19日及8月1日。常绿树种和落叶树种的监测方式略有差异。对于常绿树种,各树种选择3棵样树,各样树上随机选择20个样枝,在各时期监测以下内容:新枝的长度;在新枝上随机选择5—10个样针测定其针叶长度。根据以下公式可得到常绿针叶树种各时期各样枝上的总叶面积:
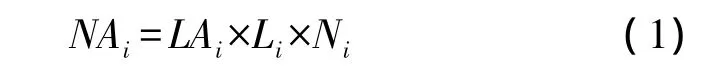
式中,NAi为树种i每个样枝上的总针叶面积;LAi为树种i每个针叶的叶面积;Li为树种i各调查时期样枝的长度;Ni为树种i单位样枝(1 cm)上针叶的数量。本研究中,假设红松针叶的横截面为等边三角形,云杉针叶的横截面为正方形,单个针叶面积具体计算公式参照Liu等[20]。
对于落叶树种,各树种选择3棵样树,各样树上随机选择1个样枝,在各时期监测各样枝上所有叶子的叶长和叶宽。根据以下公式可得到各样枝上的总叶面积:
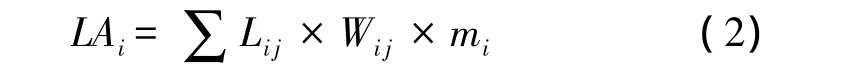
式中,LAi为树种i样枝上所有叶子的总叶面积;Li为树种i样叶j的叶长;Wi为树种i样叶j的叶宽;mi为树种i的叶面积的调整系数,本研究中主要落叶树种叶面积的调整系数参考Liu等[20],兴安落叶松是通过扫描法获得成熟时期单个叶片的真实叶面积,而后除以其长宽乘积,获得叶面积的调整系数,其值为0.63。
利用各树种叶面积最大时期各样枝上所有新生叶片的总叶面积,将其他时期样枝上所有新生叶片的总叶面积进行标准化,既得到各树种生长季节新生叶的叶面积增加比例,本研究中落叶树种利用叶面积增加比例代替LAI增加比例。
1.4 光学仪器法
半球摄影法(光学仪器法)采用Winscanopy2006冠层分析仪(Regent,Instruments Inc.,Quebec,Canada)采集数据,主要由数码相机(Coolpix 4500,Nikon,Tokyo,Japan)和 180°鱼眼镜头(Nikon FCE8)组成。生长季节图像采集时间与展叶调查同步,9—11月,每两周采集1次。采集数据时避免直射光,选择阴天或日出、日落前后,保持镜头离地面1.3 m。采集图像时,相机和鱼眼镜头保持水平,设置自动曝光状态。
1.5 凋落物法
分别于2011、2012年的5月初至9月,每月收集1次凋落物,9—11月每两周收集1次,收集完放入写好标签的塑料袋内,带回实验室,按树种将凋落叶分开。分别称重后,及时将选取的样品在65℃下烘干至恒重,测其干重,结合各树种的SLA,得到各树种在各样点各时期因凋落产生的LAI,各树种SLA参照Liu等[20]的研究结果。生长季节(5月初至9月)将每月凋落物总量按照展叶调查日期等分,获得各个展叶调查监测时期因凋落产生的LAI。
落叶树种,通过累加落叶季节(8—11月)的凋落叶产生的LAI可得到全年最大时期(8月初)的LAI,最大值结合生长季节新生叶LAI的增加比例可得到生长季节各时期的LAI,最大值依次减去落叶季节凋落叶产生的LAI可得到落叶季节各时期的LAI,从而得到各落叶树种LAI的季节动态(5—11月)。
常绿针叶树种全年均有凋落现象,因此生长季节各时期的LAI可通过以下公式计算:

式中,LAI5-1为5月1日常绿针叶树种林冠上的LAI,此时林冠上几乎不存在落叶树种的叶片,只存在常绿针叶,可根据上一年常绿针叶8月份的LAI值减去8—次年5月份凋落针叶产生的LAI获得,而本研究通过先测定一定周期内(以11—次年11月份为周期,即1a)凋落针叶产生的LAI,乘以针叶的存活周期即得到常绿针叶树种最大时期的LAI,本研究区域内红松的平均存活周期为4a[21-22],云杉的平均存活周期为 5a[22];ΔLAI为各时期通过展叶增加的LAI;LAIlitter为展叶测定周期内针叶凋落产生的LAI。
常绿针叶树种通过展叶增加的总LAI(ΔLAItotal)通过以下公式得到:
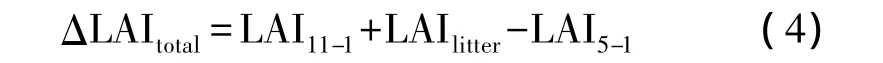
式中,LAI11-1为11月初常绿针叶树种林冠上的LAI,根据最大时期LAI减去8—11月针叶凋落产生的LAI获得;LAIlitter为5—11月针叶凋落产生的LAI;LAI5-1为 5月1日常绿针叶树种林冠上的 LAI。ΔLAItotal结合常绿针叶生长季节新生针叶LAI增加比例即可得到各树种各时期通过展叶增加的LAI(ΔLAI),然后结合公式(3)即可得到各时期的总LAI,同时利用最大时期总LAI将其标准化,可得到针叶树种各生长季节总LAI的增加比例。
常绿针叶树种落叶季节LAI的动态变化可通过各针叶树种最大时期LAI减去落叶季节针叶凋落产生的LAI获得,结合生长季节动态变化即可得到各针叶树种LAI的季节动态。综合各针、阔叶树种即可得到整个林分LAI的季节动态。
1.6 异速生长方程法
各树种的异速生长方程采用Wang[23](以下为异速生长方程-A)和董利虎等[24](以下为异速生长方程-B)的关于叶生物量与胸高直径之间的相关关系,结合SLA得到各树种最大时期的LAI,而后结合生长季节各个时期总LAI的增加比例及落叶季节的凋落物数据,可得到LAI的季节动态。本文在计算常绿针叶树种最大时期LAI时,没有考虑LAI的年际差异。
1.7 综合法
大部分光学仪器计算LAI是在林冠内叶片随机分布的假设下,且忽略了树干等木质部分的影响,因此为得到准确LAI需要进行合理校正,经过前人理论的发展及验证,LAI(L)计算如下[25]:

式中,Le为有效叶面积指数;α为木质部分面积占总面积的比率;γE为针叶针面积与簇面积的比率,用于量化针簇内集聚现象,本研究中红松及云杉的γE根据Liu等[20]方法测定,分别为1.77及1.28,再根据两树种的胸高断面积比例加权得到该林分的针簇比;ΩE为集聚指数,用于量化冠层水平集聚效应。
本研究通过公式(5)得到5月初及11月初常绿针叶树种较准确的总LAI,根据各树种胸高断面积比例得到各常绿针叶树种该时期的LAI,通过公式(3,4),结合生长季节LAI增加比例及凋落物数据即可得到常绿针叶树种LAI的季节动态;落叶树种的季节动态与凋落物法相同。综合各针、阔叶树种即可得到整个林分LAI的季节动态。
1.8 数据处理
利用 Photoshop CS 8.01(Adobe Systems Inc,USA)软件中的仿制像章工具消除5月初及11月初半球摄影图像中树干等木质部分的影响(用α值表示)[20],对两个时期的值进行t检验。半球摄影图像采用广泛应用的 DHP(DigitalHemispherical Photography)软件处理,处理时提取30—60°天顶角范围内的LAI。5月初及11月初冠层水平上的集聚效应(用 ΩE值表示)采用 DHP-TRACWin 软件[26-27]计算得到的ΩE值校正,校正时选用天顶角40—45°测量ΩE,对两个时期的值进行t检验。
2 结果与分析
2.1 木质部分所占比例(α)及集聚指数(ΩE)
5月及11月初,木质部分产生的误差之间不存在显著差异(P>0.05)(表 2),且均占总 LAI的13%,表明此时常绿树种产生的LAI占光学仪器测定LAI值的最大比例为87%;5月初各样点冠层水平上的集聚效应略大于11月初(P>0.05),分别为0.90±0.04、0.92±0.03。
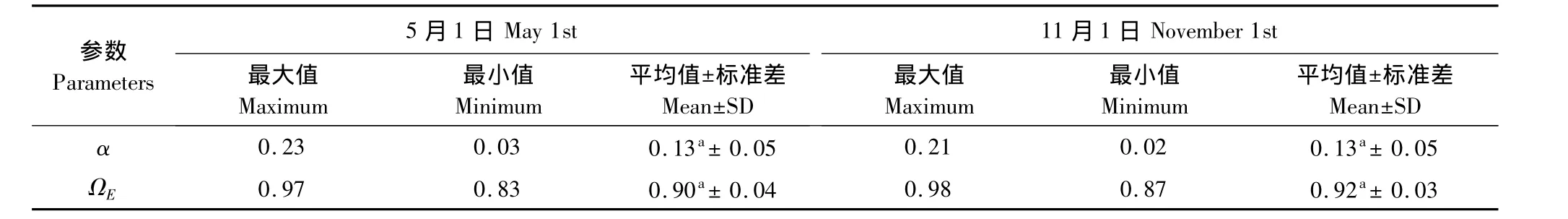
表2 小兴安岭红松人工林木质部分所占比例及集聚指数(n=18)Table 2 The woody-to-total area ratio and clumping index in the Korean pine plantation in Xiaoxing'an Mountains(n=18)
2.2 主要树种叶面积增加比例
总体而言,叶面积增加比例树种间存在明显差异(表3)。针叶树种中,红松展叶起始时间迟于兴安落叶松,长势较平缓,展叶几乎持续整个调查期,6月中旬进入生长高峰期,持续约1个月;兴安落叶松于5月上半月进入展叶高峰期,至6月初展叶已完成全年叶面积增长量的89%;而云杉5月后半月开始展叶,且快速生长持续约两周,6月上旬展叶已完成96%,7月初停止生长。阔叶树种中,水曲柳展叶起始时间较晚,6月初进入生长高峰期,持续约一个月,7月初展叶已完成97%;白桦叶增长相对平缓,展叶几乎持续整个调查期,即5—8月。
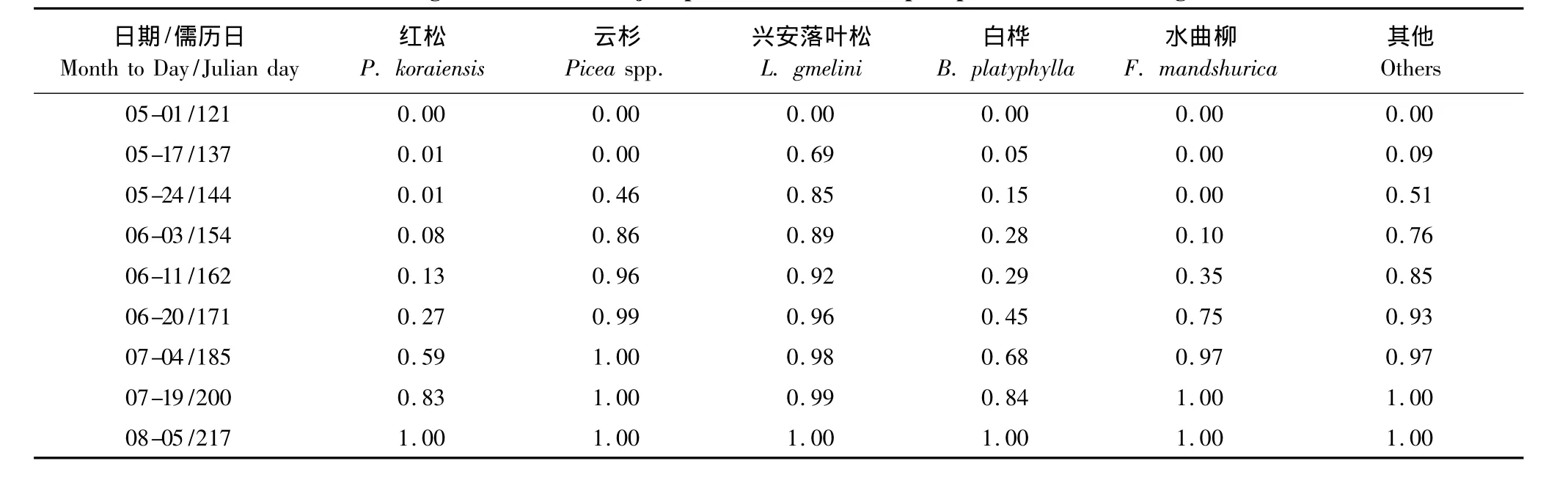
表3 小兴安岭红松人工林主要树种叶面积增加比例Table 3 Leaf area growth ratio for major species in the Korean pine plantation in Xiaoxing'an Mountains
2.3 凋落物动态
红松人工林内主要组成树种的叶凋落物候有明显差异(图1)。红松存在两个明显的落叶高峰期,在6月最先达到第1个落叶高峰,第2个出现在9月下半月,在10月后仍存在叶凋落现象,这与常绿针叶树种自身特性有关。而白桦、水曲柳、兴安落叶松及其他树种均存在一个明显落叶高峰期,其中白桦最先在8月份达到落叶高峰期,且是唯一在6、7月份即出现明显凋落现象的落叶树种,11月初叶凋落基本结束;水曲柳叶凋落持续时间最短,9月上半月进入落叶高峰期,9月下半月叶凋落量次之,至10月中旬叶凋落基本结束;其他树种叶凋落较少,且凋落较平缓;兴安落叶松落叶较晚,8月份开始出现凋落现象,且至10月初凋落叶仅占总量的11.88%,10月上半月进入落叶高峰期,持续约2周,10月下半月仍出现少量叶凋落。

图1 各调查时期红松人工林主要树种凋落物产生的LAIFig.1 LAI estimated from litter of major species in the Korean pine plantation during the investigation periods
2.4 主要树种的最大叶面积指数(LAImax)特征
总体看来,红松人工林内红松LAImax时期的LAI在所有树种中占绝对优势,主要源于红松为优势树种(相对优势度为62.97%)(图2)。不同方法测定红松LAI的排列顺序为:异速生长方程法-B(6.46)>凋落物法(5.59)>异速生长方程法-A(3.92)>综合法(2.40)。对于所有落叶树种,异速生长方程法-B测定的LAI大于其他方法。综合法测定的LAI普遍较低,常绿针叶树种尤为明显。
2.5 不同方法测定LAI季节动态
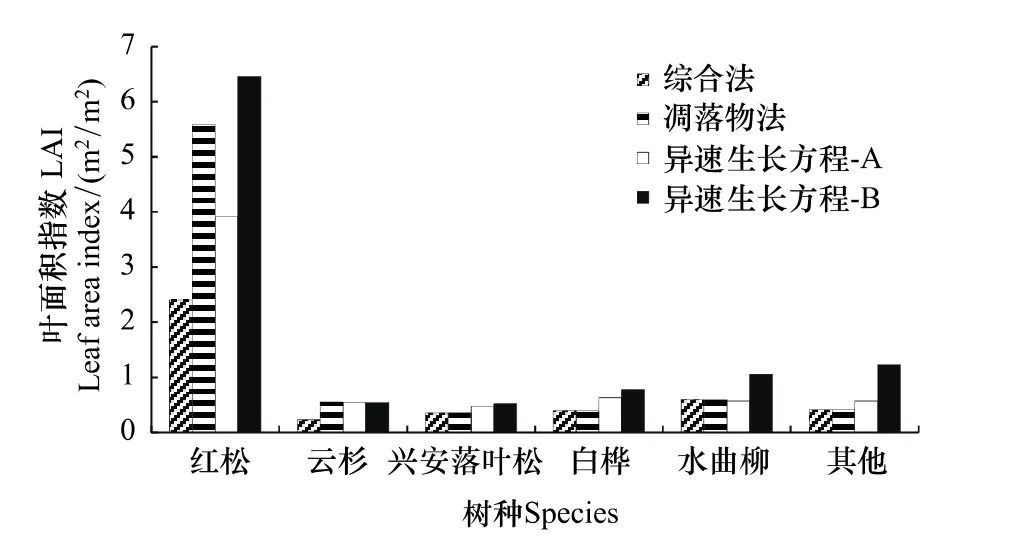
图2 不同方法测定红松人工林主要树种LAImaxFig.2 The annual maximum LAI for major species of Korean pine plantation derived from different methods

图3 不同方法测定红松人工林LAI的季节动态Fig.3 Seasonality of LAI of Korean pine plantation from different methods
整个调查期内,不同方法测定的LAI均呈单峰型,5月初值最小,持续增长至8月初达到峰值,9月中旬前LAI减小幅度较小,后因进入落叶高峰期LAI迅速减小(图3)。光学仪器法测定的红松人工林LAI明显低于其他测定方法,相对于综合法,光学仪器法最大时期低估58.86%,整个调查期平均低估48.90%;相对于凋落物法,分别低估 77.06%、75.50%;相对于异速生长方法-A,分别低估 72.93%、70.18%;相对于异速生长方法-B,分别低估82.86%、81.69%。在整个调查期中,不同方法测定LAI值差异较大,排列顺序为异速生长方法-B>凋落物法>异速生长方法-A>综合法>光学仪器法,其峰值分别为10.58、7.90、6.70、4.41 和 1.81;最小值分别为 5.87、4.07、3.33、1.38 和 0.81。
3 讨论
目前,因光学仪器法能方便、快捷地测定LAI而被广泛应用,随着高分辨率数码相机技术的发展,使得半球摄影法更具优势[28-29],而这种技术因自身限制存在一定误差也得到广泛认同[30-31]。误差主要源于木质部分和集聚效应,对于针叶林,集聚效应不仅存在于冠层水平,也存在于针簇水平,本研究有效地量化了以上因素产生的误差。
对于落叶林型,以往研究多利用背景值的方法去除树干等木质部分产生的影响,而本研究中利用Photoshop软件有效地量化了红松人工林的α,5月和11月的α值均为13%,其他学者也得到类似结论,如 Deblonde等[11]测定挪威松(Pinus resinosa)林的α值为8%—12%、斑克松(Pinus banksiana)林的α值为10%—33%;Gower等[32]测定北方森林的 α值为5%—35%;而刘志理等[33]同样利用Photoshop软件方法测定了小兴安岭白桦次生林11月的α值为57%,可见树种是影响α值的重要因子之一。本研究中红松人工林内冠层水平的集聚指数ΩE由被广泛应用的DHP-TRACWin软件直接获得;通过实地采样测定红松人工林的簇内集聚指数γE为1.73,与其他学者的研究结果相符,如Chen等[25]测定的北方针叶林 γE值为 1.4—1.8;Bréda[9]测得针叶林的γE值为1.2—2.0。
对于综合法,整个调查期内光学仪器法测定的LAI平均低估48.90%,刘志理等[34]利用相同方法测定小兴安岭阔叶红松林、谷地云冷杉林LAI的季节动态(7—11月),得到光学仪器法比综合法分别平均低估56.5%、22.8%;本研究中的低估程度高于谷地云冷杉林,主要源于谷地云冷杉林内常绿树种的簇内集聚效应弱于红松人工林。刘志理等[33]利用综合法模拟了小兴安岭白桦次生林LAI的季节动态(5—11月),效果较好(R2=0.87)。
以往研究中凋落物法多用于测定落叶林的LAI,而且无法提供生长季节 LAI的动态变化[3,13],然而结合一定周期内的凋落物及叶子存活周期来测定常绿针叶林最大时期的 LAI也有报道,如 Sprintsin等[2]估测了半干旱地中海白松(Pinus halepensis)种植园的LAI。本研究中利用凋落物法结合展叶调查直接测定红松人工林LAI的季节动态,其变化范围为3.58—7.90,季节性变异最高为 54.69%;而Chen[15]指出针叶林LAI的季节变化在25%—30%,低于本研究结果,主要源于本研究中红松人工林的林龄约为60a,林分处于生长阶段。相对于凋落物法,整个调查期内光学仪器法测定的LAI平均低估75.50%。
相对于异速生长方程法-A和-B,整个调查期内光学仪器法平均低估70.18%和81.69%。Küßner等[35]曾报道相对于异速生长方程法,光学仪器法低估程度为37%—82%;Gardingen等[36]报道相对于破坏性取样法,半球摄影法低估50%,其他学者也得到类似结果[37-38]。
本文中3种直接测定结果也存在一定差异,其中凋落物法测定红松人工林最大时期LAI为7.90,整个调查期均值为5.87,更接近于异速生长方程法-A的结果(分别为6.70、4.85)。选用不同的异速生长方程,其结果存在差异,主要源于:1)选择建立异速生长方程标准木的林分立地指数不同;2)树木叶子生物量存在明显的年际动态,即“丰歉年”,因此建立异速生长方程的时间不同将造成标准木的叶子生物量不同。利用凋落物法的长期定位研究,可弥补异速生长方程法的不足。
本文采用异速生长方程法、凋落物法、综合法和光学仪器法探讨了小兴安岭红松人工林LAI的季节动态,分析了不同方法之间的差异。在非破坏条件下,利用凋落物法结合展叶调查能直接、有效地测定常绿针叶林LAI的季节动态,这为将来进一步有效量化常绿针叶林LAI的季节及年际动态奠定了基础。
[1] Chen J M,Black T A.Defining leaf area index for non-flat leaves.Plant,Cell and Environment,1992,15(4):421-429.
[2] Sprintsin M,Cohen S,Maseyk K,Rotenberg E,Grünzweig J,Karnieli A,Berliner P,Yakir D.Long term and seasonal courses of leaf area index in a semi-arid forest plantation.Agricultural and Forest Meteorology,2011,151(5):565-574.
[3] Cutini A,Matteucci G,Mugnozza G S.Estimation of leaf area index with the Li-Cor LAI 2000 in deciduous forests.Forest Ecology and Management,1998,105(1/3):55-65.
[4] Behera S K,Srivastava P,Pathre U V,Tuli R.An indirect method of estimating leaf area index in Jatropha curcas L.using LAI-2000 PlantCanopyAnalyzer. Agriculturaland Forest Meteorology,2010,150(2):307-311.
[5] Sea W B,Choler P,Beringer J,Weinmann R A,Hutley L B,Leuning R.Documenting improvement in leaf area index estimates from MODIS using hemispherical photos for Australian savannas.Agriculturaland ForestMeteorology, 2011, 151(11):1453-1461.
[6] Bequet R,Campioli M,Kint V,Vansteenkiste D,Muys B,Ceulemans R.Leaf area index development in temperate oak and beech forests is driven by stand characteristics and weather conditions.Trees-StructureandFunction, 2011, 25(5):935-946.
[7] Bequet R,Kint V,Campioli M,Vansteenkiste D,Muys B,CeulemansR. Influence ofstand,site and meteorological variables on the maximum leaf area index of beech,oak and Scots pine.European Journal of Forest Research,2012,131(2):283-295.
[8] Ryu Y,Sonnentag O,Nilson T,Vargas R,Kobayashi H,Wenk R,Baldocchi D D.How to quantify tree leaf area index in an open savanna ecosystem:a multi-instrument and multi-model approach.Agricultural and Forest Meteorology,2010,150(1):63-76.
[9] Bréda N J J.Ground-based measurements of leaf area index:a review of methods,instruments and current controversies.Journal of Experimental Botany,2003,54(392):2403-2417.
[10] Marshall J D,Waring R H.Comparison of methods of estimating leaf-area index in old-growth Douglas-fir.Ecology,1986,67(4):975-979.
[11] Deblonde G,Penner M,Royer A.Measuring leaf area index with the LI-COR LAI-2000 in pine stands.Ecology,1994,75(5):1507-1511.
[12] MajasalmiT, RautiainenM, StenbergP, Lukeš P.An assessment of ground reference methods for estimating LAI of boreal forests.Forest Ecology and Management,2013,292:10-18.
[13] Neumann H H, DenHartogG, Shaw R H.Leafarea measurements based on hemispheric photographs and leaf-litter collection in a deciduous forestduring autumn leaf-fall.Agricultural and Forest Meteorology,1989,45(3/4):325-345.
[14] Reich P B,Frelich L E,Voldseth R A,Bakken P,Adair C.Understorey diversity in southern boreal forests is regulated by productivity and its indirect impacts on resource availability and heterogeneity.Journal of Ecology,2012,100(2):539-545.
[15] Chen J M.Optically-based methods for measuring seasonal variation of leaf area index in boreal conifer stands.Agricultural and Forest Meteorology,1996,80(2/4):135-163.
[16] Li X R,Liu Q J,Cai Z,Ma Z Q.Specific leaf area and leaf area index of conifer plantations in Qianyanzhou station of subtropical China.Journal of Plant Ecology,2007,31(1):93-101.
[17] Ma Z Q,Liu Q J,Zeng H Q,Li X R,Chen Y R,Lin Y M,Zhang S H,Yang F T,Wang H Q.Estimation of leaf area index of planted forests in subtropical China by photogrammetry.Acta Ecologica Sinica,2008,28(5):1971-1980.
[18] Jonckheere I,Fleck S,Nackaerts K,Muys B,Coppin P,Weiss M,Baret F.Review of methods for in situ leaf area index determination:PartI. Theories,sensors and hemispherical photography.Agricultural and Forest Meteorology,2004,121(1/2):19-35.
[19] Nasahara K N,Muraoka H,Nagai S,Mikami H.Vertical integration of leaf area index in a Japanese deciduous broad-leaved forest.Agricultural and Forest Meteorology,2008,148(6/7):1136-1146.
[20] Liu Z L,Jin G Z,Qi Y J.Estimate of leaf area index in an oldgrowth mixed broadleaved-Korean pine forest in northeastern china.PLoS One,2012,7(3):e32155.
[21] Dai L M,Xu Z B,Zhang Y J,Chen H.Study on decomposition rate and fall of Pinus koraiensis needle.Acta Ecologica Sinica,2001,21(8):1296-1300.
[22] Luo T X.Patterns of Net Primary Productivity for Chinese Major Forest Types and Their Mathematical Models[D].Beijing:Chinese Academy of Sciences,1996.
[23] Wang C K.Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. ForestEcology and Management,2006,222(1/3):9-16.
[24] Dong L H,Li F R,Jia W W,Liu F X,Wang H Z.Compatible biomass models for main tree species with measurement error in Heilongjiang Province of Northeast China.Chinese Journal of Applied Ecology,2011,22(10):2653-2661.
[25] Chen J M,Rich P M,Gower S T,Norman J M,Plummer S.Leaf area index of boreal forests:theory, techniques, and measurements.Journal of Geophysical Research,1997,102(D24):29429-29443.
[26] Leblanc S G,Chen J M,Fernandes R,Deering D W,Conley A.Methodology comparison for canopy structure parameters extraction from digital hemispherical photography in boreal forests.Agricultural and Forest Meteorology,2005,129(3/4):187-207.
[27] Macfarlane C,Hoffman M,Eamus D,Kerp N,Higginson S,McMurtrie R,Adams M.Estimation of leaf area index in eucalypt forestusing digital photography. Agricultural and Forest Meteorology,2007,143(3/4):176-188.
[28] Frazer G W,Fournier R A,Trofymow J A,Hall R J.A comparison of digital and film fisheye photography for analysis of forest canopy structure and gap light transmission.Agricultural and Forest Meteorology,2001,109(4):249-263.
[29] Zhao C Y,Qi J G,Shen W H,Zou S B.Estimating the canopy coverage of Picea crassifolia forest using digital hemispherical photography in Qilian Mountains.Acta Ecologica Sinica,2009,29(8):4196-4205.
[30] Eriksson H,Eklundh L,Hall K,Lindroth A.Estimating LAI in deciduous forest stands.Agricultural and Forest Meteorology,2005,129(1/2):27-37.
[31] Ryu Y,Nilson T,Kobayashi H,Sonnentag O,Law B E,Baldocchi D D.On the correct estimation of effective leaf area index:Does itrevealinformation on clumping effects?.Agricultural and Forest Meteorology,2010,150(3):463-472.
[32] Gower S T,Kucharik C J,Norman J M.Direct and indirect estimation of leaf area index,f(APAR),and net primary production of terrestrial ecosystems. Remote Sensing of Environment,1999,70(1):29-51.
[33] Liu Z L,Jin G Z.Estimation of leaf area index of secondary Betula platyphylla forestin Xiaoxing'an Mountains. Acta Ecologica Sinica,2013,33(8):2505-2513.
[34] Liu Z L,Jin G Z.Estimation of leaf area index of three forest types in Xiaoxing'an Mountains of Northeast China.Chinese Journal of Applied Ecology,2012,23(9):2437-2444.
[35] Küßner R, Mosandl R. Comparison of direct and indirect estimation of leaf area index in mature Norway spruce stands of eastern Germany.Canadian Journal of Forest Research,2000,30(3):440-447.
[36] Van Gardingen P R,Jackson G E,Hernandez-Daumas S,Russell G,Sharp L.Leaf area index estimates obtained for clumped canopies using hemispherical photography.Agricultural and Forest Meteorology,1999,94(3/4):243-257.
[37] Sommer K J,Lang A R G.Comparative analysis of two indirect methods of measuring leaf area index as applied to minimal and spur pruned grape vines.Australian Journal of Plant Physiology,1994,21(2):197-206.
[38] Brenner A J,Cueto Romero M,Garcia Haro J,Gilabert M A,Incoll L D,Martinez Fernandez J,Porter E,Pugnaire F I,Younis M T.A comparison of direct and indirect methods for measuring leaf and surface areas of individual bushes.Plant,Cell and Environment,1995,18(11):1332-1340.
参考文献:
[16] 李轩然,刘琪璟,蔡哲,马泽清.千烟洲针叶林的比叶面积及叶面积指数.植物生态学报,2007,31(1):93-101.
[17] 马泽清,刘琪璟,曾慧卿,李轩然,陈永瑞,林耀明,张时煌,杨风亭,汪宏清.南方人工林叶面积指数的摄影测量.生态学报,2008,28(5):1971-1980.
[21] 代力民,徐振邦,张扬建,陈华.红松针叶的凋落及其分解速率研究.生态学报,2001,21(8):1296-1300.
[22] 罗天祥.中国主要森林类型生物生产力格局及其数学模型[D].北京:中国科学院,1996.
[24] 董利虎,李凤日,贾炜玮,刘福香,王鹤智.含度量误差的黑龙江省主要树种生物量相容性模型.应用生态学报,2011,22(10):2653-2661.
[29] 赵传燕,齐家国,沈卫华,邹松兵.利用半球图像反演祁连山区青海云杉(Picea crassifolia)林盖度.生态学报,2009,29(8):4196-4205.
[33] 刘志理,金光泽.小兴安岭白桦次生林叶面积指数的估测.生态学报,2013,33(8):2505-2513.
[34] 刘志理,金光泽.小兴安岭三种林型叶面积指数的估测.应用生态学报,2012,23(9):2437-2444.
猜你喜欢
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
内蒙古林业科技(2021年2期)2021-07-08
林业科学(2020年10期)2020-11-30
生态学报(2020年21期)2020-11-24
山东林业科技(2019年6期)2020-01-02
生态学报(2019年21期)2019-12-18
作文周刊·小学四年级版(2019年8期)2019-04-23
现代营销(创富信息版)(2016年6期)2016-11-21
国际木业(2016年6期)2016-02-28
