
棘胸蛙两性形态特征在个体发育过程中的变化
2014-08-02 01:03吴晓雯李晗慧焦振鑫罗来高许雪峰
滁州学院学报 2014年5期
吴晓雯,李晗慧,焦振鑫,罗来高,许雪峰
个体大小的性别差异在动物界广泛存在,也有一些假说致力于解释两性大小异形(Sexual size dimorphism, SSD)[1]。生育力选择(Fecundity selection)被认为是促成偏向雌性为较大个体的两性异形的驱动力[2]。雌体的窝卵(仔)数,或后代大小和繁殖成功率通常都随着雌体的个体大小增加而增加[3-5],这种情况见于一些卵生的外温动物,如鱼类[6],爬行动物[7-8]和两栖动物[9-11]。偏向雄性为较大个体的两性异形被认为受性选择(Sexual selection)的影响,如由于雄性间的竞争或雌性选择,个体较大的雄性通常能提高交配成功率[12]。
两性异形可能是由两性不同的生长格局所导致的,选择作用在雄性和雌性生长上将会导致成体两性在形态特征上的变化。尽管在个体发育过程中两性间形态特征的变化可能会表现出发育上的冲突,但也可能在这个阶段解决这种冲突。成体基于遗传基础的两性形态特征的进化则是非常缓慢的,而且这些差异不仅在相近物种中进化,而且也可能发生在种群间甚至一个有机体的不同特征间[13]。
研究生长过程形态特征变化的细节对理解潜在的两性异形的进化的最终机制是很有必要的。成体相似的两性异形格局能由不同的生长过程产生[14]。缺乏个体发育和生长期间选择作用的详细知识就不能理解两性异形进化上的变化。然而,到目前为止还鲜有蛙类个体发育过程的两性形态特征变异的研究,一个可能的原因是在野外动物的跟踪上仍然是有很大的困难,而直接获取野外的动物又不能明确界定动物的发育年龄。
本研究通过在实验室饲养棘胸蛙,用动物标签技术跟踪动物来阐述棘胸蛙个体发育过程的两性形态特征变化,以期解释棘胸蛙个体发育过程两性异形的进化驱动力。
1 材料和方法
实验用棘胸蛙均为2010年所产的后代,我们将棘胸蛙饲养在常年通水的水泥池中,喂食黄粉虫。亚成体形态特征为从完成变态发育到饲养至六个月时(2010年10月中旬至2011年3月中旬)测得,成体为饲养至第一个繁殖季节前(2012年4月中旬)测得。用数显游标卡尺测量棘胸蛙亚成体体长(吻端至泄殖腔的距离,snout-urostyle length, SUL)、头长(吻端至下颌关节后缘的距离)、头宽(头部最宽处距离)、腹长(前肢后缘到后肢前缘的距离)、前肢长(肩关节至第三趾末端的距离)、后肢长(体后端正中部位至第四趾末端的距离)和体重(湿重)。每一个动物在形态特征测量完成后都对其皮下注射PIT-tag标签标注动物,然后饲养在水泥池中,于2012年4月中旬将已标注的成活棘胸蛙用 PIT-tag标签阅读器读出标签编号,然后按照上述实验方法再一次测量其形态特征数据,用于后续分析。
所有的数据在作进一步分析前,用Kolmogoror Simirnov和 Bartlett(Statistica 6.0 Software)分别检验数据正态性和方差同质性。用线性回归分析(linear regression)其它六个形态特征与体长的相关性,用单因子方差分析(one-way ANOVA)比较两性间体长的差异,用以体长为协变量的单因子协方差分析(one-way ANCOVA)比较两性间其它形态特征的差异,用Tukey’s检验比较两性间形态特征的差异。描述性统计值用平均值± 1标准误表示,显著性水平设置为α = 0.05。
2 结果
棘胸蛙亚成体体长两性间差异不显著,而成体的体长雄性大于雌性(表1和表2)。棘胸蛙亚成体和成体雌雄两性的头长、头宽、腹长、前肢长、后肢长和体重都随体长的增长而增长(linear regression, allP< 0.001)。ANCOVA 去除 SUL 差异的影响后,雄性亚成体的头宽和前肢长都要大于雌性,亚成体两性间的头长、腹长、后肢长和体重差异不显著;而去除SUL差异影响后成体的局部形态特征除了头长两性间没有差异外,其它形态变量差异显著,雌性在腹长上大于雄性,而头宽、前肢长、后肢长和体重雄性都大于雌性(表1 和表 2)。
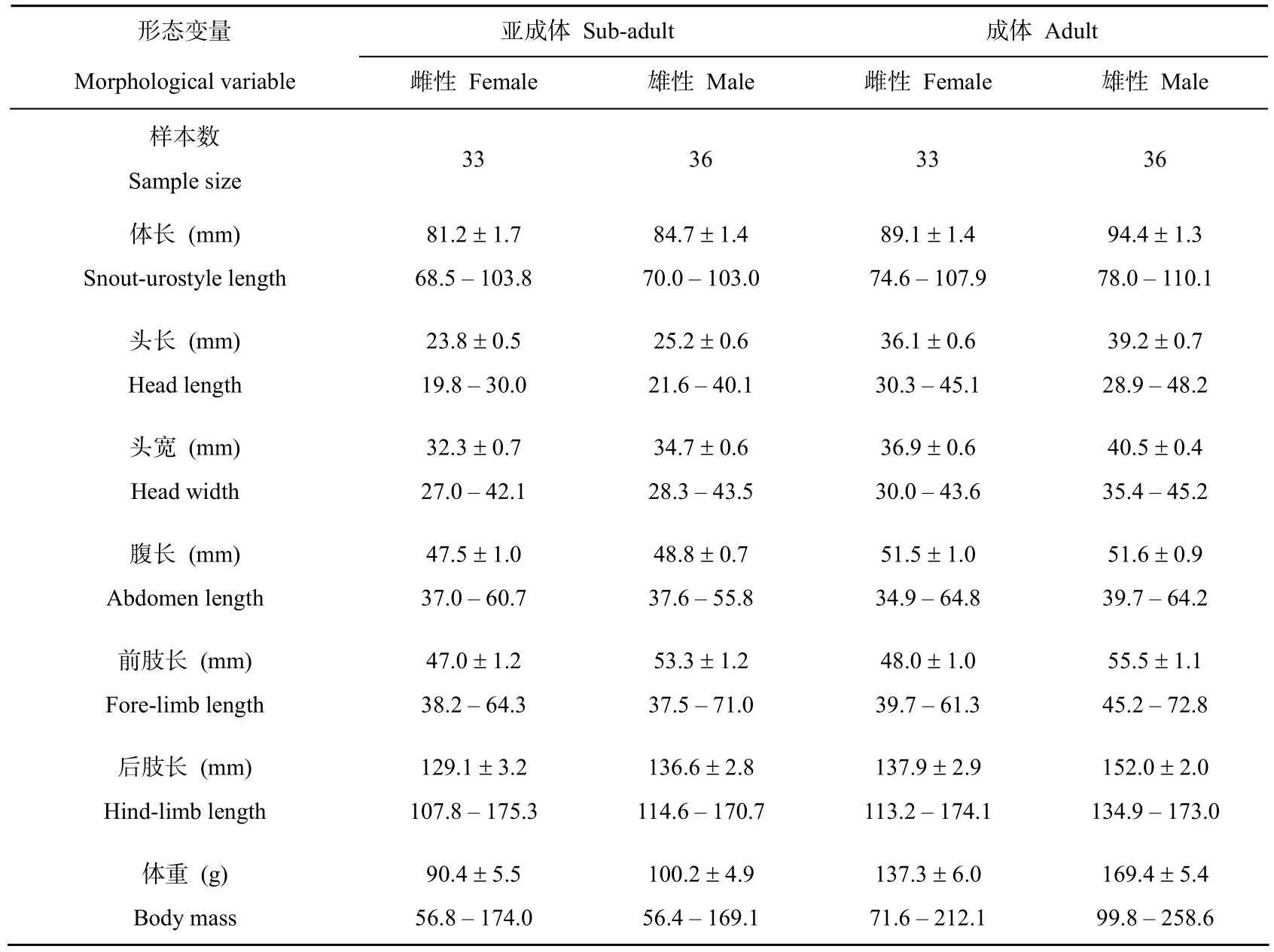
表1 棘胸蛙形态特征的描述性统计值
3 讨论
无尾两栖类动物中 90%的种类雌性的体长都大于雄性[15]。本研究中,棘胸蛙亚成体两性在个体大小上差异不显著,而发育到性成熟后,雄性的体长却比雌性的要大(见表1)。因此,棘胸蛙是一种偏向于雄性为较大个体的 SSD模式两栖动物。
棘胸蛙成体雄性具有较大体长这一结果与该种已有研究的结果相似[16]。棘胸蛙多生活于海波600m以上的山区具有临时性池塘的石壁上,而这样的生境不易获得,因此,棘胸蛙的雄性成体体形较大,可以在雄-雄竞争中具有较大的优势,保护其领域不被侵袭。另外,雄性具有较大体长可能会获得更多的交配机会,这样选择作用就有利于选择体形较大的雄性,以提高雄性的繁殖成功率[15,17-18]。在本研究中,与提高雄性繁殖成功率的形态特征变量均在选择作用力下得到强化,如头大小强化了雄性的配偶获得能力,前肢长强化了雄性抱对的能力,后肢长强化了其运动能力(表1和表 2),这一研究与爪蟾的结果相似[19]。尽管棘胸蛙雌性成体体型较小,但是腹长这一与雌性生育力直接相关的形态特征雌性却大于雄性(表 1和表2),生育力选择作用支持雌性向较大腹长的方向进化以利于提高繁殖输出并强化其繁殖成功率。这种结果见于其他的一些动物类群[8]。
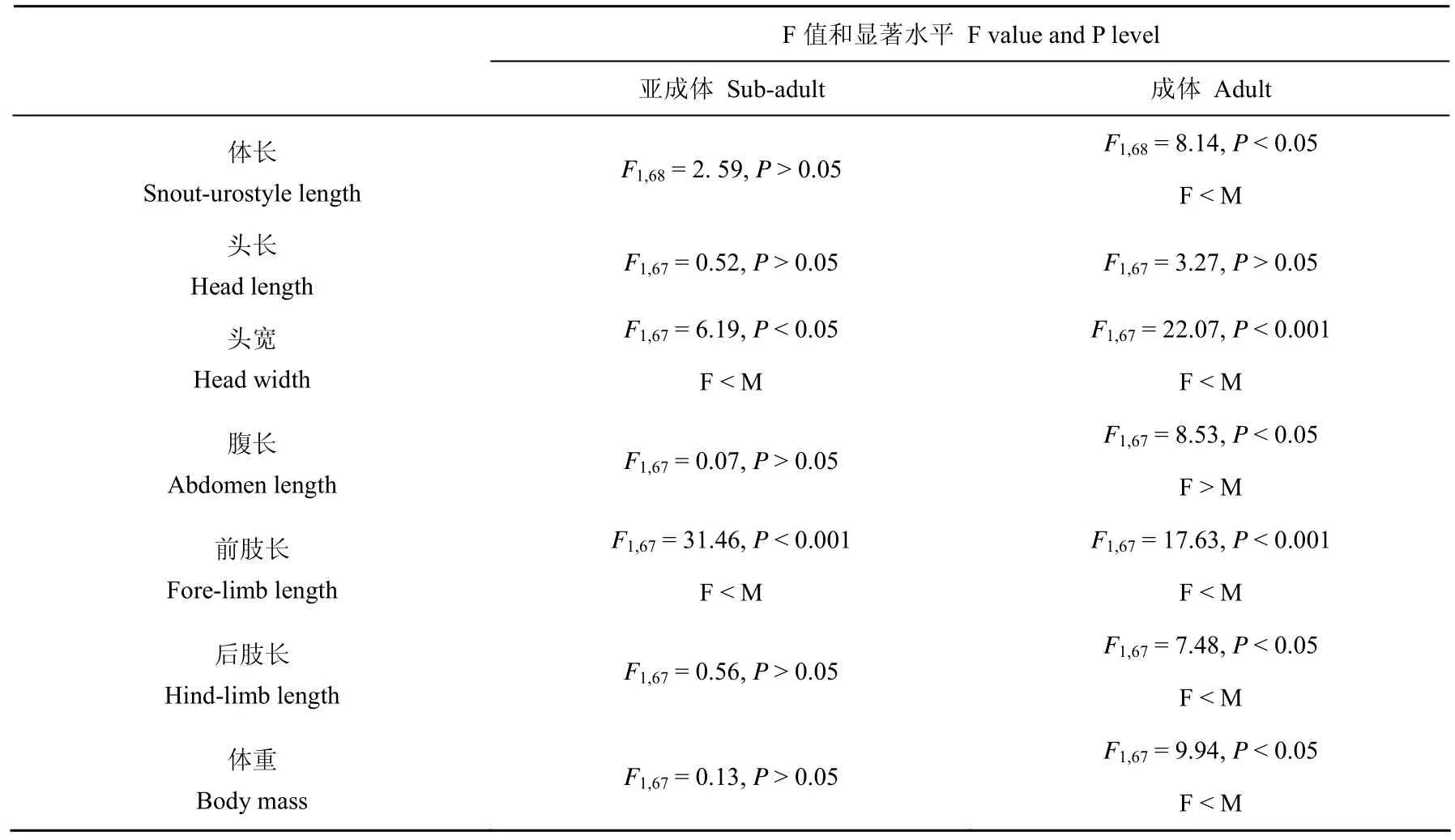
表2 棘胸蛙两性间形态特征的单因素方差和单因素协方差分析结果
棘胸蛙在亚成体时两性还保持着几乎同步的生长速率,而在亚成体至性成熟这一个体发育阶段,雌性与雄性呈现出异速的生长速率,那么,为什么棘胸蛙在个体发育过程中表现出不同的两性个体大小异形格局,且为什么棘胸蛙两性形态特征大小的差异为什么发生在个体发育的后期?已有研究者在其它动物类群中开展了个体发育过程的两性异形的实验研究,并形成了特定个体发育时期的两性冲突的进化选择理论[14]。这个理论包括:第一、两性形态特征大小差异出现在个体发育的后期能把两性间的冲突降到最低的程度。这一理论已经在其他物种的研究上得到支持[20]。我们推测,棘胸蛙两性间在个体发育的早期阶段保持均质的个体发育速度也可能能够减少两性间的冲突。第二、个体发育过程中两性间的冲突的强度依赖于生长周期的长度。进而言之,这种强度依赖于选择作用在促进或削弱两性的同一表型时这一周期的相对时间和这一时期选择作用的相对强度。通常来说,两性大小异形受选择作用在成体期两性形态特征的大小差异作用在繁殖成功率上的贡献的影响。因此,在棘胸蛙亚成体至成体个体发育阶段,选择作用强化了能影响繁殖成功率的形态特征,如雌性向着较大的腹长和雄性向着较大的体长、头大小及与提高雄性生育力有关的性状特征上进化。
[1] Lu D; Zhou CQ; Zhao LJ; Liao WB. Pattern of sexual size dimorphism supports the inverse of Rensch’s rule in two frog species [J]. Animal Biology, 2014, 64 (1): 87-95.
[2] Darwin C. The Descent of Man and Selection in Relation to Sex [M]. John Murray, London. 1871.
[3] Cadeddu G; Giacoma C; Castellano S. Sexual size dimorphism in the Tyrrhenian tree frog: a life-history perspective [J]. Journal of Zoology, 2012, 286 (4): 285 –292.
[4] Pincheira-Donoso D, Tregenza T. Fecundity selection and the evolution of reproductive output and sex-specific body size in the Liolaemus lizard adaptive radiation [J].Evolutionary Biology, 2011, 38 (2): 197 – 207.
[5] Roff DA. The evolution of life histories [M]. Chapman and Hall, London. 1992.
[6] Elgar M.A. Evolutionary compromise between a few large and many small eggs: comparative evidence in teleost fish[J]. Oikos, 1990, 59 (2): 283 – 287.
[7] Cox RM, Butler MA, John-Alder HB. The evolution of sexual size dimorphism in reptiles. In: Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism. (Ed. Fairbairn DJ, Blanckenhorn WU,Szekely T.) [M]. Oxford University Press, Oxford. 2007,38 - 49.
[8] Luo LG, Wu YL, Zhang ZY, Xu XF. Sexual size dimorphism and female reproduction in the white-striped grass lizard Takydromus wolteri [J]. Current Zoology,2012, 58 (2): 236 – 243.
[9] Kupfer A. Sexual size dimorphism in amphibians: an overview. In: Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism. (Ed. Fairbairn DJ,Blanckenhorn WU, Szekely T.) [M]. Oxford University Press, Oxford. 2007, 50 – 59.
[10] Kupfer A. Sexual size dimorphism in caecilian amphibians:analysis, review and directions for future research [J].Zoology, 2009, 112 (5): 362 – 369.
[11] Wells KD. The behavioral ecology of anuran communication.In: Hearing and sound communication in amphibians.Springer handbook of auditory research, (ed Narins PM,Feng AS.) [M]. Springer Verlag, Now York. 2007, 44 – 86.
[12] Andersson S. Sexual Selection. Princeton University Press,Princeton [M]. 1994.
[13] Badyaev AV. Growing apart: an ontogenetic perspective on the evolution of sexual size dimorphism [J]. Trends in Ecology Evolution, 2002, 17 (8): 369 – 378.
[14] Badyaev AV, Hill GE, Wittingham LA. The evolution of sexual size dimorphism in the house finch. IV. Population divergence in ontogeny of dimorphism [J]. Evolution,2001, 55 (12): 2534 – 2549.
[15] Kaplan RH, Salthe SN. The allomertry of reproduction: an empirical view in salamanders [J]. The American Naturalist, 1979, 113 (5): 671 – 689.
[16] Yu BG; Zheng RQ; Zhang Y; Liu CT. Geographic variation in body size and sexual size dimorphism in the giant spiny frog Paa spinosa (David, 1875) (Anura:Ranoidae) [J]. Journal of Natural History, 2010, 44(27-28): 1729 – 1741.
[17] 林植华, 计翔. 浙江丽水中国石龙子的食性、两性异形和雌性繁殖[J]. 生态学报, 2000, 20 (2): 304 – 310.
[18] 杜卫国, 计翔. 蓝尾石龙子的生长、两性异形和雌性繁殖[J]. 动物学研究, 2001, 22 (4): 279 – 289.
[19] Herrel A; Gonwouo LN; Fokam EB; Ngundu WI;Bonneaud C. Intersexual differences in body shape and locomotor performance in the aquatic frog, Xenopus tropicalis [J]. Journal of zoology, 2012, 287 (4): 311 –316.
[20] 罗来高. 影响虎纹蛙繁殖成功率的因子分析[D]. 南京:南京师范大学. 2010.
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
湖南畜牧兽医(2020年6期)2020-03-22
猪业科学(2016年1期)2016-03-30
中国造纸学报(2015年1期)2015-12-16
化工进展(2015年3期)2015-11-11
材料研究与应用(2015年4期)2015-08-23
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
中学生物学(2008年12期)2008-12-27
