
侧柏林土壤呼吸速率对不同模拟降雨量的响应
2014-09-27 11:24朱梦洵饶良懿李会杰
水土保持通报 2014年2期
朱梦洵,饶良懿,李会杰,苏 赛
(1北京林业大学 水土保持学院,北京100083;2.水土保持与荒漠化防治教育部重点试验室,北京100083)
土壤是陆地生态系统中最大的碳库,数据显示土壤中碳约占全球碳储量的67%[1]。土壤呼吸是土壤中碳以CO2形式释放到大气中的过程,土壤呼吸的变化对大气中CO2的平衡产生很大的影响[2],研究土壤呼吸及其影响因子与全球气候变化的响应关系具有重要意义。其中降雨对土壤呼吸的作用相对复杂,包括一系列物理、生物及生物化学过程。研究资料[3]表明,20世纪全球范围内强降雨事件的频率和强度均有所增加。大气环流模型预测,21世纪全球总降雨量将不会发生很大变化,但是极端降雨事件将会不断增加[4]。全球气候变化对区域尺度的降雨格局产生了很大的影响,近年来降雨量、降雨频率、降雨季节分配在一些地区有不同的改变[5-6]。降雨量及降雨频率的不同将直接导致土壤水分状况发生变化,而土壤水分状况对研究土壤呼吸的影响机理甚至是预测未来土壤碳储量变化都具有重要意义[7]。然而预测森林中土壤碳存贮量和流通量,不仅需要了解森林土壤呼吸对土壤平均含水量的依赖关系,而且要了解土壤碳流失量对降雨模式改变的响应[8]。为此,本研究以不同林龄侧柏为例,通过设置不同降雨量(5,10,20,50mm)及连续相同频率的模拟降雨来观测土壤呼吸的变异规律,主要的研究目的包括3个方面:(1)30a林龄侧柏林土壤呼吸对不同模拟降雨量的响应;(2)10a林龄侧柏林土壤呼吸对不同模拟降雨量的响应;(3)不同模拟降雨条件下林龄对土壤呼吸速率的影响及影响因素。
1 材料与方法
1.1 研究区概况
研究样地设立于北京市鹫峰国家森林公园。鹫峰森林公园地处华北平原西部,位于太行山北部,燕山东端,距北京市西北约30km,属北京大西山范围。坐标为39°54′N,116°28′E,公园面积811.17hm2,海拔在50~1 100m。年平均气温为11.6℃,年降水量650~750mm,其中夏季降水量占全年降水量的74%,年蒸发量为1 800mm,干燥度1.9。公园森林覆盖率高达96.2%,共有陆地植物110科313属684种,公园四季气候明显,夏季炎热多雨,冬季寒冷干燥,具有暖温带的气候特征。鹫峰森林公园周围的天然植被是典型的暖温带落叶阔叶林,现在多为次生林和人工林,目前群落类型主要有:油松(Pinus tabulaef ormis)林、侧柏(Platycladus orientalis)林、栓皮栎(Quercus variabilis)林、栓皮栎与槲栎(Quercus aliena)混交林、刺槐(Robinia pseudoacacia)林等[9]。
1.2 模拟降雨试验
在鹫峰森林公园分别选取10m×10m的侧柏成林和幼林观测样地。分别在每个观测样地内选取灌木和草本层尽量一致的5棵生长状况良好的侧柏,每棵侧柏周围布设6个0.5m×0.5m的样方,对小样方使用人工模拟降雨设备(有均匀小孔的洒水壶)模拟降雨,其中1—4号样方分别做梯度为5,10,20,50mm的降雨强度处理,且每周浇一次水。另外两个样方分别做连续相同频率降雨处理(即50mm降雨分5次实施,同样1周为1个模拟降雨周期)和空白对照处理(CK),即6种处理5组重复。
1.3 土壤呼吸测定
从2012年8—10月每次上午10:00开始进行土壤呼吸的测定。土壤呼吸速率采用静态密闭气室法(碱液吸收法)[10]测定。密闭气室采用规格一致的下端开口的塑料透明保鲜盒(直径18cm,高30cm),于试验开始前一个月埋入样方土壤内插入10cm深,并盖土砸实以防止漏气。测定前一天贴地面剪除保鲜盒内地表植被,将内盛有20ml浓度为1mol/L的NaOH溶液的玻璃广口瓶(测定时打开瓶盖,高10 cm,内径6cm)放在保鲜盒内离地面2cm的三脚支架上并扣上保鲜盒盖子。放置24h后取出广口瓶迅速带回试验室用标准盐酸溶液进行滴定。
1.4 数据处理
试验数据使用Excel软件进行初步整理,使用SPSS 18.0统计软件对不同林龄侧柏林土壤呼吸与不同模拟降雨量进行多因素方差分析。
2 结果与分析
2.1 不同模拟降雨量对30a侧柏林土壤呼吸速率的影响
采用箱线图直观地对比了对照处理(CK)和5种模拟降雨量条件下30a侧柏林土壤呼吸速率的差异(图1)。由图1可知,5,10和20mm降雨量处理后的30a侧柏林土壤呼吸速率的总体值较高,中位数值均高于CK(对照组),反映该3种降雨处理对30年生侧柏林的土壤呼吸速率有促进作用。50mm降雨量处理后的30年生侧柏林土壤呼吸速率的中位数值比CK低,这可能是由于50mm降雨量使土壤水分达到过饱和状态,氧气向土壤传输受到阻碍,导致土壤呼吸速率降低。与陈胜全等[11]研究的当土壤处于过干或者过湿状态时,土壤呼吸会受到抑制的结论一致。图1中与其它处理对比,持续相同降雨频率处理后的30年生侧柏林土壤呼吸速率中位数最低,说明土壤呼吸速率在连续降雨处理后明显受到抑制。
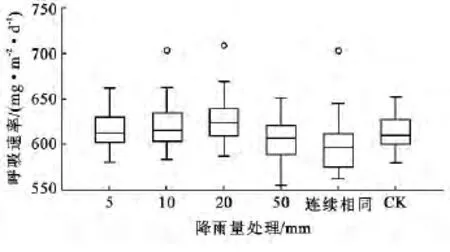
图1 30年生侧柏林在5种降雨量处理下的土壤呼吸速率对比箱线图
当地气象数据显示,首次模拟降水前研究区附近降雨稀少,土壤中水分含量相对较低。10和20mm及相同频率降雨处理组中的土壤呼吸速率均出现离群值(图1),而该3种模拟降雨最高值均出现在测量起始日(图2)。造成这种现象的原因可能是,突然施加的人工降雨对土壤水分进行了补偿,激发了土壤中的根系及微生物的生理活性,改变了呼吸底物的有效性,从而使土壤呼吸速率突然增大。随着周期性人工降雨的施加,降雨对该土壤水分的激发效应消失,所以不再出现离群值。该趋势与其他学者的研究结果相一致[12]。CK组的土壤呼吸速率在9月1日(645.61mg/(m2·d)出现了峰值,而其它处理组土壤呼吸速率相对较低(图2),气象资料显示在9月1日发生了天然降雨事件,这改善了对照组的土壤水分状况,使土壤呼吸作用得以促进并且该效应持续了3 d。这与Wu等[13]研究结果一致,在土壤面临干旱胁迫时,降雨会使土壤微生物量和土壤呼吸值激增。同时该次天然降雨事件对其他降雨处理组的土壤呼吸产生了相对抑制的作用。
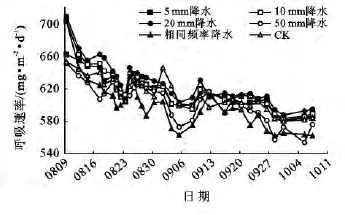
图2 30年生侧柏林在不同模拟降雨处理下土壤呼吸速率的变化
2.2 不同模拟降雨量对10年生侧柏林土壤呼吸速率的影响
由图3可以看出,在5mm降雨量处理下10年生侧柏林土壤呼吸速率的整体值最高。另外其它降雨处理的中位数值均低于CK对照组。各雨量处理的中位数值由大到小的顺序为:5mm雨量处理〔600.26g/(m2·d)〕>CK组〔592.64mg/(m2·d)〕>10mm雨量处理〔583.20mg/(m2·d)〕>20mm雨量处理〔577.94mg/(m2·d)〕>相同降雨频〔572.06 mg/(m2·d)〕>50mm雨量处理〔571.68mg/(m2·d)〕。说明各处理中5mm雨量处理对10年生侧柏林的土壤呼吸速率起到促进作用,其余模拟降雨量均对10年生侧柏林土壤呼吸速率造成抑制效果。原因可能是10年生侧柏林与30年生侧柏林相比,根呼吸、微生物活性或其他底物控制因素对降雨量比较敏感,5mm的模拟降雨处理后已使其土壤接近最大持水量,其他过多的降雨反而会在不同程度上抑制10年生侧柏林的土壤呼吸作用。
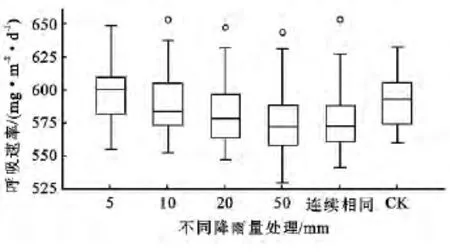
图3 10年生侧柏林在5种降雨处理下的土壤呼吸速率对比箱线图
在箱线图中10,20,50mm和连续相同频率降雨处理下的土壤呼吸速率均出现了极值(图3),除CK对照组外,其余处理组的土壤呼吸速率最高值均出现在起始测量日(图4)。可能是由于测量前土壤处于相对干旱的状态,突然的人工降雨对10年生侧柏林同样起到了极大的促进作用,使得10,20,50mm及相同频率降雨处理下的土壤呼吸速率在首次测量日出现了极大离群值(图3)。8月15日之前,所有模拟降雨处理组的呼吸速率均在不同程度上高于CK对照组,但是随着模拟降雨的周期性重复,土壤水分条件得到逐步的改善,施加降雨对10年生侧柏林土壤呼吸速率的促进趋势减缓,甚至自8月15日后,20,50mm及相同频率降雨处理对10a侧柏林的土壤呼吸速率起到抑制作用趋势(图4)。从8月24日起10 mm降雨处理对10年生侧柏林土壤呼吸速率的影响开始有抑制趋势(图4)。这可能是由于随着降雨量的增多土壤水分含量相应增加,土壤孔隙度降低,土壤透气性变差,影响了土壤微生物的活性,从而抑制了10年生侧柏林的土壤呼吸作用。由50mm与连续相同频率降雨处理的土壤呼吸速率趋势线对比可知(图4),土壤呼吸速率不只是受降雨总量的影响,还与降雨频率相关。
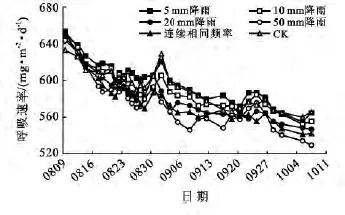
图4 10年生侧柏林在不同模拟降雨处理下土壤呼吸速率的变化
2.3 林龄对侧柏林土壤呼吸速率在不同降雨条件下的影响
用每个处理中5个重复测定的密闭气室土壤呼吸速率的平均值作为该样地中当次测量的土壤呼吸速率。对分别进行不同模拟降雨处理的两种林龄的侧柏林土壤呼吸速率进行了多因素方差分析。分析结果表明,林分年龄的不同对侧柏林土壤呼吸速率存在显著的影响(p<0.001)。不同降雨处理也对侧柏林土壤呼吸速率的影响具有显著差异(p<0.001)。同时不同林龄和不同降雨量间的交互作用对侧柏林土壤呼吸速率有较显著影响(p=0.028)。
不同林龄间通过对照处理(CK)可观察到(图5),30年生侧柏林的平均土壤呼吸速率﹝611.68mg/(m2·d)﹞明显高于10年生侧柏林的平均土壤呼吸速率﹝591.58mg/(m2·d)﹞。林龄对侧柏林的土壤呼吸速率有显著影响。同时图1还显示相同林龄内,与对照处理(CK)相比不同模拟降雨处理组,对各林龄侧柏林的土壤呼吸速率有促进也有抑制作用,但30年生侧柏林在5组不同模拟降雨处理下的平均土壤呼吸总是高于其相同模拟降雨处理下的10年生侧柏林的平均土壤呼吸速率。这说明相同林龄内不同降雨量对侧柏林土壤呼吸速率有不同的影响,但不同降雨量并未对原本30年生侧柏林土壤呼吸速率大于10年生侧柏林土壤呼吸速率的这一整体趋势造成改变性影响。
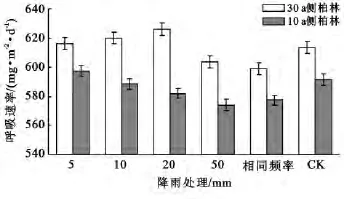
图5 相同降雨处理下不同林龄间侧柏林土壤呼吸速率的关系
3 结果讨论
3.1 土壤呼吸和不同降雨量间的关系
降水既影响土壤含水量,又可以通过冲刷或淋溶作用促进地上的有机残体向地下运输,从而促进微生物的活性与种群数量增加[12,14]。在两种林龄的CK组中,9月1日的天然降雨均对侧柏林的土壤呼吸速率均产生促进作用。这是由于降雨过后,土壤中水分迅速增加,土壤中的微生物数量会在雨后激增[15],因此降雨会对土壤呼吸速率有所促进。也有文献认为是由于降雨促进了根呼吸作用从而使土壤呼吸量有所增加。不同林龄侧柏林土壤呼吸对降雨量有不同的响应,5种模拟降雨处理中,5,10和20mm模拟降雨量对30年生侧柏林土壤呼吸速率均有促进作用(图2)。而对10年生侧柏林施加同样模拟降雨后发现只有5mm降雨量对其土壤呼吸速率有促进作用,其余处理组与CK比较均出现抑制作用(图4)。这符合Zhang等[4]研究的过大降雨量或过高降雨频率会减少土壤中碳排放的结论。可能是由于当土壤含水量在一定的阈值内变化时,呼吸速率会随着其增加而增大,但是当土壤含水量的变化超过这一阈值时,呼吸强度反而会随着土壤水分的增加而减小[11]。从两种林龄侧柏林对降雨量需求的对比中发现,10年生侧柏林对降雨量的需求小于30年生侧柏林。这可能是由于随着林龄的增大,30年生侧柏林的凋落物、根生物量以微生物种群数量大于10年生侧柏林,因此30年生侧柏林对土壤水分的需求较大。
还可以通过图1和图3观察降雨量同样是50 mm但由于降雨频率不同(即一次性施加50mm和5次连续施加10mm降雨),两种林龄侧柏林土壤呼吸速率所产生的响应也不同。两种模拟降雨量相比,一次性施加50mm降雨量对10年生侧柏林的土壤呼吸速率产生的抑制作用大于连续相同频率降雨处理组,而30年生侧柏林却表现出相反的结果。Christopher[16]认为降雨频率的变化,可能比降雨量或其他对CO2通量产生影响的因素更加重要。
3.2 土壤呼吸和林龄间的关系
根据多因素方差分析可以看出,林龄对侧柏林土壤呼吸速率有显著的影响(p<0.01)。不同的模拟降雨量对侧柏林的土壤呼吸作用也产生了相应的影响,但与林龄的影响相比,降雨和林龄的交互作用对侧柏林土壤呼吸速率的影响程度相对减弱。相关文献[17]表明,根呼吸和微生物呼吸及土壤有机质的不同是导致林龄对土壤呼吸速率产生影响的主要因素。Ewel[18]对美国佛罗里达松树林的研究发现29年生松树林的土壤呼吸速率比9年生松树林的土壤呼吸速率高35%。同时还发现与9年生松树林相比29年生松树林含有高于其3倍的根生物量。这被认为是导致林龄对土壤呼吸速率产生影响的原因。研究者们[19]发现随着林龄的增加,地上生物量显著增加,甚至同理推断地下生物量同样也是随着林龄增加而增加。通过对各种森林的研究[20]证实,作为碳源之一的凋落物会随着林龄的增加而增加,而且凋落物在土壤呼吸各成分中的贡献率较大,达到17%~48%。同时根系分泌物和凋落物有机残体进入土壤会对微生物呼吸产生很大的影响。随着林龄的增加,30年生侧柏林生物量大于10年生侧柏林,这可能促进了微生物呼吸,从而导致30年生侧柏林的土壤呼吸速率显著高于10年生侧柏林。而且与土壤呼吸密切相关的细根生物量在一定范围内也是随着林龄的增加而增加[21],这可能是导致30年生侧柏林土壤呼吸速率高于10年生侧柏林的另一个原因。
通过3个月对侧柏林生长季后期的观测,得到30和10年生侧柏林的土壤呼吸速率平均值分别为611.7和571.6mg/(m2·d)(图5)。所观察到的大林龄侧柏林的土壤呼吸速率高于小林龄侧柏林土壤呼吸速率的趋势,符合 Wiseman等[18]在美国弗吉尼亚州对不同林龄火炬松土壤呼吸速率的研究结果。但是与Saiz[22]在爱尔兰得出的10年生云杉林土壤呼吸速率大于47年生云杉林的研究结果相反。表明林龄对土壤呼吸速率有显著影响,但是侧柏林土壤呼吸速率是否一直随着林龄增大而增大,目前为止还没有一致的研究结论。
4 结论
(1)林龄对侧柏林土壤呼吸速率有很大的影响。30年生侧柏林的土壤呼吸速率大于10年生侧柏林。
(2)相同林龄内降雨量在一定范围内会促进土壤呼吸,但超过一定阀值将会对土壤呼吸起到抑制作用。本研究中所模拟的各种降雨量尽管对同一林龄内的土壤呼吸产生了不同影响,但并未改变两林龄间原本30年生侧柏林土壤呼吸速率大于10年生侧柏林土壤呼吸速率的整体趋势。
[1]Jenkison D S,Adams D E,Wild A.Model estimates of CO2emissions from soil in response to global warming[J].Nature,351(6324):304-306.
[2]Wang Genxu,Qian Ju,Chen Guodong,et al.Soil organic carbon pool of grassland soils on the Qinghai—Tibetan Plateau and its global implication[J].Science of The Total Environment,2002,291(1/3):207-217.
[3]Easterling D R,Meehl G A,Parmesan C.Climate extremes:Observations,modeling,and impacts[J].Science,2000,289(5487):2068-2074.
[4]Tebaldi C,Hayhoe K,Arblaster J.Going to the extremes:An intercomparison of model-simulated historical and future changes in extreme events[J],Climatic Change,2007,82(1/2):233-234.
[5]王义东,王辉民,马泽清,等.土壤呼吸对降雨响应的研究进展[J].植物生态学报,2010,34(5):601-610.
[6]IPCC.Climate Change 2007:the Physical Science Basis[R].Geneva:Intergovernmental Panel on Climate Change,2007.
[7]邓东周,范志平,王红,等.土壤水分对土壤呼吸的影响[J].林业科学研究,2009,22(5):722-727.
[8]Zhang Jiaying,Lin Zhongbin,Zhang Renduo,et al.Effects of simulated rainfall events on soil carbon transformation[J].Australian Journal of Soil Research,2010,48(5):404-412.
[9]赵永泉,彭道黎.北京鹫峰公园主要人工林群落多样性研究[J].西南林学院学报,2008,28(1):17-18.
[10]陈四清,崔骁勇,周广胜,等.内蒙古锡林河流域大针茅草原土壤呼吸和凋落物分解的CO2排放速率研究[J].植物学报,1999,41(6):645-650
[11]陈胜全,李凌浩,韩兴国,等.水分对土壤呼吸的影响及机理[J].生态学报,2003,23(5):972-978.
[12]Huxman T E,Snyder K A,Tissue D.Precipitation pulses and carbon fluxes in semiarid and arid ecosystems[J].Oecologia,2004,141(2):254-268.
[13]Wu J,Brookes P C.The proportional mineralization of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil[J].Soil Biology and Biochemistry,2005,37(3):507-515.
[14]Schimel J P,Balser T C,Wallenstein M.Microbial stress-response physiology and its implications for ecosystem function[J].Ecology,2007,88(6):1386-1394.
[15]Gordon H,Haygarth P M,Bardgett R D.Drying and rewetting effects on soil microbial community composition and nutrient leaching[J].Soil Biology and Biochemistry,2008,40(2):302-311.
[16]Christopher W H,John M B,Philip A F,et al.Increased rainfall variability and reduced rainfall amount decreases soil CO2flux in a grassland ecosystem[J].Global Change Biology,2005,11(2):322-334.
[17]Gong Jirui,Ge Zhiwei,An Ran,et al.Soil respiration in poplar plantations in northern China at different forest ages[J].Plant Soil,2012,360(1):109-122.
[18]Ewel K C,Cropper J R,Gholz H L.Soil CO2evolution in Florida slash pine plantations(I):Changes through time[J].Journal of Forest Research,1987,17(4):325-329.
[19]Wiseman P E,Seiler J R.Soil CO2efflux across four age classes of plantation loblolly pine(Pinus taeda L.)on the Virginia Piedmont[J].Forest Ecology and Management,2004,192(2/3):297-311.
[20]Nouvellon Y,Epron D,Marsden C.Age-related changes in litter inputs explain annual trends in soil CO2effluxes over a full Eucalyptus rotation after afforestation of a tropical savannah[J].Biogeochemistry,2012,111(1):515-533.
[21]李凌浩,林鹏.武夷山甜槠林细根生物量和生长量研究[J].应用生态学报,1998,9(4):337-340
[22]Saiz G,Byrne K A,Bahl K B,et al.Stand age-related effects on soil respiration in a first rotation Sitka spruce chronosequence in central Ireland[J].Global Change Biology,2006,12(6):1007-1020.
猜你喜欢
现代装饰(2022年5期)2022-10-13
防护林科技(2022年5期)2022-08-22
防护林科技(2020年10期)2020-12-22
考试与评价·七年级版(2020年6期)2020-11-02
四川林业科技(2020年1期)2020-08-31
海峡姐妹(2017年7期)2017-07-31
高原山地气象研究(2016年2期)2016-11-10
快乐作文·低年级(2015年1期)2015-03-26
亚热带资源与环境学报(2015年1期)2015-01-22
塔里木大学学报(2014年3期)2014-03-11
