
城市公园植被特征对陆生鸟类集团的影响
2015-01-19 05:48丁由中裴恩乐王小明王正寰
生态学报 2015年14期
杨 刚, 许 洁, 王 勇, 丁由中, 袁 晓, 裴恩乐, 马 波, 王小明, 王正寰,*
1 华东师范大学生命科学学院,上海 200062 2 华东师范大学上海市城市化过程与恢复重点实验室,上海 200241 3 上海市野生动植物保护管理站,上海 200023 4 上海科技馆,上海 200127
城市公园植被特征对陆生鸟类集团的影响
杨 刚1,2, 许 洁1,2, 王 勇1,2, 丁由中1,2, 袁 晓3, 裴恩乐3, 马 波3, 王小明4, 王正寰1,2,*
1 华东师范大学生命科学学院,上海 200062 2 华东师范大学上海市城市化过程与恢复重点实验室,上海 200241 3 上海市野生动植物保护管理站,上海 200023 4 上海科技馆,上海 200127
城市公共绿地是城市生态系统中重要的鸟类栖息地,其植被特征对鸟类集团存在显著影响。在通过分析植被特征对陆生鸟类集团的作用,从而为公园合理配置植被来提高其作为野生动物栖息地的生态服务功能提供理论基础。2009 年10 月至2011 年10 月,采用样线法对上海滨江森林公园进行鸟类调查,利用主成分分析划分鸟类集团,用高度定义植被层次,用卡方检验分析鸟类行为在植被层次上的差异。结果表明,滨江森林公园陆生鸟类群落在乔木层的栖息行为和运动行为频次显著多于其在灌木层和地被层的行为频次,在地被层的取食行为频次显著多于其在乔木层和灌木层的行为频次。陆生鸟类可划分为8 个鸟类集团,鸟类集团之间存在栖息、运动和取食空间生态位的重叠。食虫拾取集团、杂食拾取集团、食肉飞取集团和植食拾取集团在栖息、运动和取食空间生态位上均存在较高的重叠度,其通过食性分离各自空间生态位。食虫探取集团和食虫飞取集团互为栖息空间生态位重叠度最高集团,其通过取食方式的不同来实现生态位的分离。根据公园植被特征对鸟类集团的影响结果对上海市公园绿地植被配置提出了建议。
植被特征;鸟类集团;生态位重叠;主成分分析;鸟类行为
城市绿地作为城市野生动物尤其是城市鸟类重要的栖息地,对维持城市生态系统的稳定起着至关重要的作用[1-2]。近年来,研究城市鸟类群落结构尤其是鸟类集团成为城市生态学研究的热点之一[3-5]。鸟类集团是指以相似方式利用相同等级生境资源的鸟类类群[6]。研究集团之间或集团内物种之间的相互关系,对于研究群落构成,物种之间的竞争和资源分割有着重要意义[7]。
上海作为中国城市化率最高的城市,鸟类资源丰富,从20 世纪初至今共记录到鸟类438种,自2000年以来共整理记录到鸟类373 种[8]。上海城市绿地作为野生鸟类的主要栖息地之一,为鸟类直接或间接的提供食物,甚至成为很多鸟类,尤其是陆生鸟类的唯一避难所[9-11]。因此,公共绿地的建设和保护一直是上海生态城市建设的重要组成部分,同时城市陆生鸟类保护也长期受到学术界的关注[12-15]。然而,公共绿地的植被配置主要考虑景观和休闲的需要,这限制了其作为野生动物栖息地的生态服务功能[16-17]。研究表明,不同鸟类集团对于植被群落结构的要求并不一致,自然林地中食虫鸟类集团、筑地面巢鸟类集团和树洞巢鸟类集团数量显著多于人工林,而在城市环境中,这些鸟类的数量显著少于人工林[5]。Degraaf[18]对于美国城市郊区鸟类集团的研究表明,地面觅食植食性和杂食性鸟类,地面营巢鸟类和灌木层营巢鸟类倾向选择成熟自然生境;冠层营巢鸟类,树洞巢鸟类和树枝营巢鸟类倾向选择次生林生境,并且食虫鸟密度在次生林中最大。同时,有研究指出,通过改进植被结构可以有效提高城市绿地的鸟类多样性水平[19-20]。因此,研究陆生鸟类集团对植被结构的偏好,可以为提高城市化地区鸟类多样性水平提供依据,同时对城市公园作为鸟类栖息地生态服务功能的实现具有现实的指导意义。
本文以上海市典型的大型城市公共绿地——滨江森林公园为研究地点,通过分析鸟类群落组成和结构,研究公园植被特征对陆生鸟类集团的影响。本研究主要关注3 个科学问题:(1)通过数值分类方法可以将陆生鸟类群落分为哪些鸟类集团,(2)各陆生鸟类集团行为在植被垂直分层结构上有何差异,(3)各陆生鸟类集团间的生态位重叠度有何差异。通过解决以上科学问题,为公园合理配置植被,提高公园陆生鸟类多样性,在确保城市绿地休闲功能的同时,实现其作为野生动物栖息地的生态服务功能。
1 研究方法1.1 研究地点
上海位于亚洲大陆东缘,属于亚热带季风气候,年均降水量1213 mm,年平均气温16.9 ℃。上海位于澳大利亚至西伯利亚鸟类迁徙的中点,是候鸟的重要迁徙中转站[21]。上海2000 年以来已记录到鸟类中,陆生鸟类260 种,水鸟113 种[8]。
上海滨江森林公园(31°14’ N, 121°29’ E)位于上海市浦东新区高桥镇,黄浦江、长江和东海在此汇聚,该公园由28a的苗圃改建而成,于2007 年对公众开放。总面积约120 hm2,是一个典型的大型城市公园。公园的生境类型有常绿落叶阔叶混交林、常绿阔叶林、暖性针叶林、针阔混交林、落叶阔叶林、灌丛和疏林草坪。公园改建过程中共调查到木本植物160 余种,草本植物170 余种。常见乔木有落羽杉(Taxodiumdistichum)、池杉(T.ascendens)、枫杨(Pterocaryastenoptera)、香樟(Cinnamomumbodinieri)、广玉兰(Magnoliagrandiflora)、榉树(Zelkovaserrata)等;常见灌木有杜鹃(Rhododendronsimsii)、八角金盘(Fatsiajaponica)、珊瑚树(Viburnumodoratissimum)、卫矛(Euonymusalatus)、海桐(Pittosporumtobira)、垂丝海棠(Malushalliana)等;常见地被植物有诸葛菜(Orychophragmusviolaceus)、红花酢浆草(Oxaliscorymbosa)、麦冬(Ophiopogonjaponicus)、虎耳草(Saxifragastolonifera)、狗牙根(Cynodondactylon)等[22-23]。公园的2010 年至2012 年年均游客接待量为20 余万人次。
1.2 样线设置
在公园内随机设置40 个样点,样点间隔≥100 m。将相邻的2 个样点联接,联接的路线作为公园鸟类调查的样线,样线长度约5.5 km,样线穿过公园各种生境类型。
1.3 鸟类调查
2009 年10 月至2011 年10 月,选择晴朗无风的工作日,在鸟类活动的高峰时段(7:00—10:00; 16:00—18:00)用双筒望远镜(10×42 mm)对公园鸟类进行调查,每月调查2 次。采用样线法进行鸟类调查,以1.5—2.0 km/h的速度沿样线前进, 记录样线左右两侧各25 m之内直接观察到的鸟类个体,记录其在植被上的位置和行为,对于飞过和仅以鸣声分辨的鸟类不做记录。鸟类行为划分为3 种:
(1)栖息 鸟类没有明显的位置移动,停留在某个位置休憩,鸣叫,整理羽毛等;
(2)运动 鸟类在多个位置移动,追逐,嬉戏,搜寻食物,打斗等;
(3)取食 鸟类啄取食物,正在进食昆虫,蠕虫,果实或其他食物资源。
1.4 鸟类集团分类
根据鸟类食性、取食方式以及在植被群落各层的活动情况对陆生鸟类进行鸟类集团划分。参考赵正阶[24]及本研究的观察,鸟类食性分为4 种:
(1)食虫 食物以昆虫及其幼虫为主;
(2)杂食 兼食昆虫和植物材料;
(3)植食 以植物材料为食;
(4)食肉 以鱼类、鸟类、哺乳动物等脊椎动物为食。
参考王勇等[25]、刘菡等[26]及本研究的观察,将鸟类取食方式分为3 种:
(1)拾取 用喙直接从地面或叶层表面啄取静止或移动缓慢的食物;
(2)探取 用喙啄破外壳或爪穿破树干、粗枝表面, 探取树干、粗枝表面下的食物;
(3)飞取 在空中追赶捕食移动迅速的猎物。
按照高度[27]划分植被层次,记录鸟类所处的植被层次:
(1)乔木层h>2.5 m;
(2)灌木层0.5 m≤h≤2.5 m;
(3)地被层h<0.5 m。
如此, 可对每种鸟进行食性、取食方式和植被层次评估, 符合变量要求的赋值1, 其余的赋值0。使用主成分分析(PCA)进行降维计算, 最终确定鸟类集团的种类。
1.5 数据分析
对各鸟种变量的得分情况进行主成分分析(PCA), 截取特征值≥1的主成分用于最终的降维分析。在各主成分中选取载荷量绝对值≥0.6的变量作为该主成分的显著变量[28]。本研究最终确定使用4 个主成分(PC1—PC4)。根据各主成分中显著变量的构成确定该主成分主要反映的鸟类集团信息,将各鸟种的变量数据分别代入4 个主成分, 以其在各主成分中的最终得分来判断该鸟种的鸟类集团归属。用卡方检验分析整体鸟类群落以及陆生鸟类集团在乔木层,灌木层和地被层的分布是否具有显著差异。采用Morisita-Horn指数[29]:
式中,Oik代表种i的资源利用曲线与种k的资源利用曲线的重叠指数,Pij代表种i在第j个资源状态下的个体数占该种所用个体数的比例。计算不同鸟类集团间的空间生态位重叠度(栖息,运动和取食)。
2 结果与分析
2.1 植被特征对鸟类群落的影响

图1 滨江森林公园鸟类行为在植被结构上的分布
本次研究共记录到陆生鸟类65 种(表2),鸟类行为6965 次(图1)。鸟类群落在乔木层的栖息行为和运动行为频次显著多于其在灌木层和地被层的行为频次(栖息行为:χ2= 3440.51,P<0.001;运动行为:χ2= 148.57,P<0.001),而鸟类群落在地被层的取食行为频次显著多于其在乔木层和灌木层的行为频次(χ2= 27.78,P<0.001)。
2.2 鸟类集团分类
基于鸟类位置,食性和取食方式的PCA分析结果显示,前4 个主成分(PC1—PC4)的特征值均大于1,累积贡献率达到69.824%,因此截取前4 个主成分进入降维分析。在PC1中,显著的变量(载荷量绝对值≥0.6的变量)为食肉,飞取和探取,因此该主成分定义为食肉的食性以及飞取和探取的取食方式。按照此定义原则,PC2定义为食虫和杂食的鸟类食性,PC3定义为拾取和探取的取食方式,PC4主要定义为植食食性(表1)。
以PC1和PC2以及PC3和PC4分别建立直角坐标系, 对65 种陆生鸟类进行数值分类归类,生成8 个鸟类集团(图2a、b),分别是: 食虫拾取集团(IC), 包括18 种鸟类, 代表物种有白鹡鸰(Motacillaalba)、斑鸫(Turdusnaumanni); 食虫探取集团(IP), 包括9 种鸟类, 代表物种有北红尾鸲(Phoenicurusauroreus)、红胁蓝尾鸲(Tarsigercyanurus);食虫飞取集团(IF),包括5种鸟类, 代表物种有北灰鹟(Muscicapadauurica)、灰纹鹟(M.griseisticta); 杂食性拾取集团(OC), 包括20 种鸟类, 代表物种有白头鹎(Pycnonotussinensis)、灰椋鸟(Sturnuscineraceus); 杂食性探取集团(OP), 仅有白眉鹀(Emberizatristrami); 植食拾取集团(HC), 包括5 种鸟类, 代表物种有黑尾蜡嘴雀(Eophonamigratoria)、金翅(Carduelissinica); 植食探取集团(HP), 仅包括燕雀(Fringillamontifringilla) 1 种;食肉飞取集团 (RF), 包括6 种鸟类,代表物种有雀鹰(Accipiternisus)、棕背伯劳(Laniusschach)(表2)。

表1 基于鸟类位置,食性和取食方式的PCA分析结果

图2 滨江森林公园鸟类集团分析
2.3 植被特征对鸟类集团的影响
2.3.1 植被特征对食虫拾取集团的影响
食虫拾取集团共记录行为1314 次,其中栖息行为1123 次,运动行为92 次,取食行为99 次(图3)。该鸟类集团在乔木层的栖息行为频次显著多于其在灌木层和地被层的行为频次(χ2= 729.36,P<0.001),在乔木层和灌木层的运动行为频次显著多于其在地被层的行为频次(χ2= 30.15,P<0.001),在地被层的取食行为频次显著多于其在乔木层和灌木层的行为频次(χ2= 58.61,P<0.001)。

表2 本研究记录到的滨江森林公园陆生鸟类名录及其所属鸟类集团
2.3.2 植被特征对食虫探取集团的影响
食虫探取集团共记录行为231 次,其中栖息行为194 次,运动行为16 次,取食行为21 次(图3)。该鸟类集团在乔木层,灌木层和地被层的栖息行为频次无显著差异(栖息行为:χ2= 2.98,P= 0.225,运动行为χ2= 5.38,P= 0.068,栖息行为:χ2= 5.43,P= 0.066)。
2.3.3 植被特征对食虫飞取集团的影响
食虫飞取集团共记录到栖息行为25 次,未观察到运动和取食行为(图3)。该鸟类集团在乔木层和地被层的栖息行为频次显著少于其在灌木层的行为频次(χ2= 7.28,P= 0.026)。
2.3.4 植被特征对杂食拾取集团的影响
杂食拾取集团共记录到行为3871 次,其中栖息行为3350 次,运动行为233 次,取食行为288 次(图3)。该鸟类集团在乔木层的栖息行为和运动行为频次显著多于其在灌木层和地被层的行为频次(栖息行为:χ2= 1829.80,P<0.001;运动行为χ2= 139.22,P<0.001),而在地被层和灌木层的取食行为频次显著多于其在乔木层的行为频次(χ2= 22.90,P<0.001)。
2.3.5 植被特征对杂食探取集团的影响
杂食探取集团共记录到栖息行为8 次,未观察到运动行为和取食行为(图3)。该鸟类集团在乔木层和灌木层的栖息行为频次无显著差异(χ2= 2.00,P= 0.30)。

图3 滨江森林公园各集团鸟类行为在植被结构上的分布
2.3.6 植被特征对植食拾取集团的影响
植食拾取集团共记录到行为1132 次,其中栖息行为817 次,运动行为74 次,取食行为241 次(图3)。该鸟类集团在乔木层的栖息行为频次显著多于其在灌木层和地被层的行为频次(χ2= 920.24,P<0.001),在乔木层和灌木层的运动行为频次显著多于其在地被层的行为频次(χ2= 21.51,P<0.001),在地被层和乔木层的取食行为频次显著多于其在灌木层的行为频次(χ2= 22.54,P<0.001)。
2.3.7 植被特征对植食探取集团的影响
植食探取集团共记录到行为116 次,其中栖息行为70 次,运动行为1 次,取食行为45 次(图3)。该鸟类集团在乔木层的栖息行为频次显著多于其在灌木层的行为频次(χ2= 35.71,P<0.001),在乔木层的取食行为频次显著多于其在地被层的行为频次(χ2= 33.80,P<0.001)。
2.3.8 植被特征对食肉飞取集团的影响
食肉飞取集团共记录到行为268 次,其中栖息行为236 次,运动行为20 次,取食行为12 次(图3)。该鸟类集团在乔木层的栖息行为频次显著多于其在灌木层和地被层的行为频次(χ2= 111.87,P<0.001),在乔木层,灌木层和地被层的运动行为和取食行为频次无显著差异(运动行为:χ2= 6.10,P= 0.060,取食行为:χ2= 1.50,P= 0.598)。
2.4 陆生鸟类集团的空间生态位重叠度
对于栖息空间生态位,食虫拾取集团与杂食拾取集团互为重叠度最高的集团,为0.998;食虫探取集团和食虫飞取集团互为重叠度最高集团,为0.818;杂食探取集团与食虫拾取集团的重叠度最高,为0.985;植食拾取集团与植食探取集团互为重叠度最高集团,为0.991;食肉飞取集团与杂食拾取集团的重叠度最高,为0.994。
对于运动空间生态位,食虫拾取集团与食肉飞取集团互为重叠度最高的集团,为0.997;食虫探取集团和植食拾取集团互为重叠度最高集团,为0.989;杂食拾取集团与食肉飞取集团的重叠度最高,为0.914;植食探取集团与食虫拾取集团的重叠度最高,为0.289。
对于取食空间生态位,食虫拾取集团与杂食拾取集团重叠度最高,为0.757;食虫探取集团与杂食拾取集团重叠度最高,为0.761;杂食拾取集团与食肉飞取集团互为重叠度最高的集团,为0.834;植食拾取集团与食虫拾取集团重叠度最高,重叠度为0.593;植食探取集团与食虫探取集团重叠度最高,为0.171(表3)。
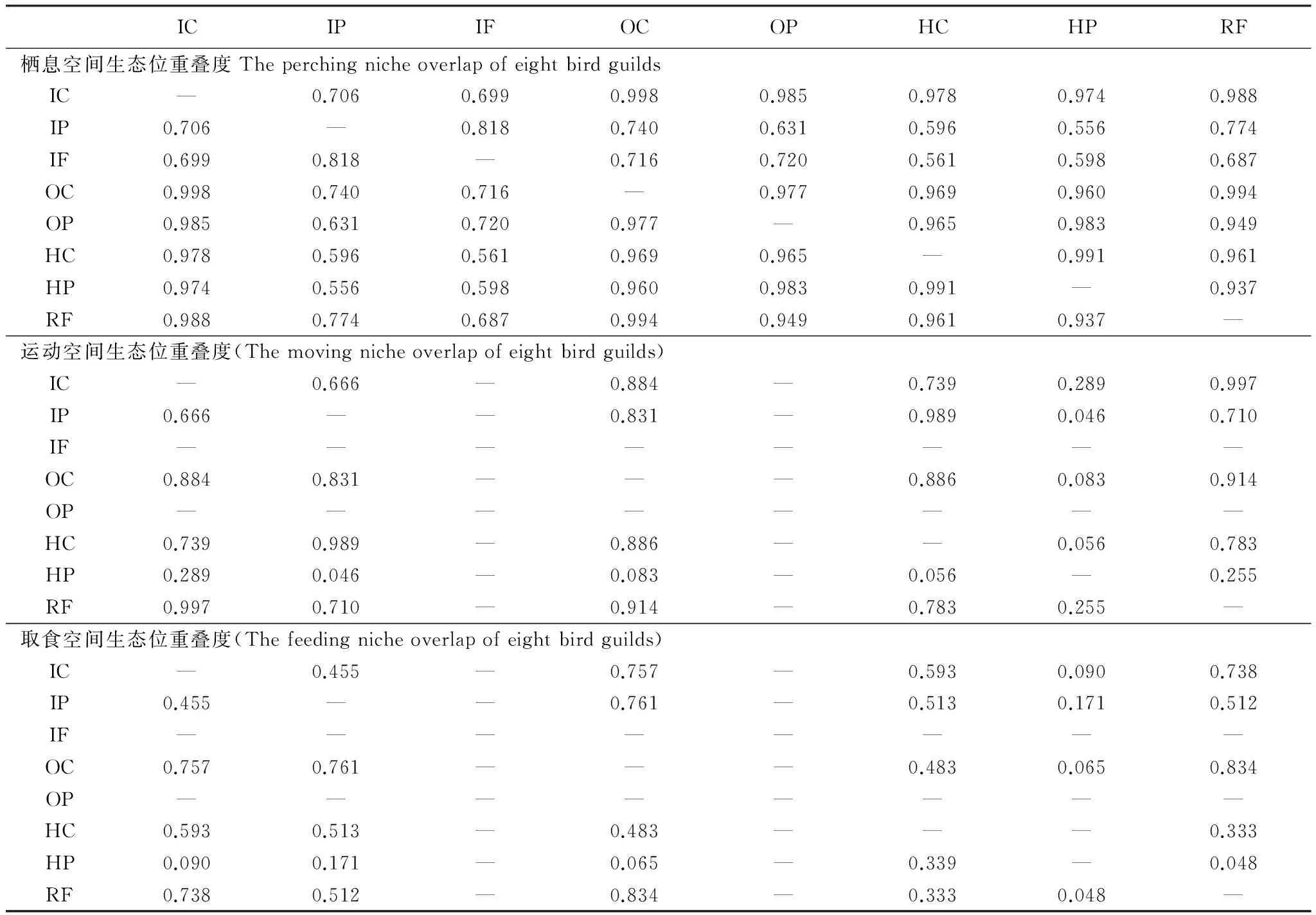
表3 8种鸟类集团间的空间生态位重叠度
3 讨论
3.1 公园特征对鸟类群落和鸟类集团的影响
公园陆生鸟类群落3 种行为在植被层次上存在显著差异,乔木层为鸟类群落提供主要的栖息和运动场所,而地被层为其提供主要的取食场所。但植被层次对8 种鸟类集团的作用存在差异:
(1)乔木层为食虫拾取集团、杂食拾取集团、植食拾取集团、植食探取集团和食肉飞取集团提供栖息场所;为食虫拾取集团、杂食拾取集团和植食拾取集团提供运动场所;为植食拾取集团和植食探取集团提供取食场所。Clergeau 等[30],Palomino和Carrascal[31]和Evans 等[32]的研究结果表明,乔木层作为重要的植被结构能够增加鸟类物种的丰富度和多样性水平,可以为大多数鸟类提供栖息,筑巢,隐蔽和取食的场所。就栖息而言,本研究中83.5%的灰椋鸟(Sturnuscineraceus)选择乔木层栖息。在观察到6 次灰椋鸟集群中,4 次集群于高大乔木,且最大集群数量达到80 只。就运动而言,所观察到的戴菊(Regulusregulus)求偶行为均在乔木层出现。就取食而言,本研究中的2 种植食性鸟类集团主要以公园高大乔木的成熟果实或枝叶为食。如黑尾蜡嘴雀(Eophonamigratoria)和燕雀(Fringillamontifringilla),其主要食物分别为落羽杉(Taxodiumdistichum)果实和乌桕(Sapiumsebiferum)果实。
(2)食虫拾取集团,杂食拾取集团和植食拾取集团均选择地被层为主要的取食场所。Pearman[33]的研究结果表明,地被层为大多数食虫鸟类和杂食性鸟类的提供了食物的来源,这与本研究中食虫拾取集团和杂食拾取集团取食行为主要发生在地被层的结果基本一致。然而,在自然林生态系统中,包括灌木层和地被层的林下带是大多数昆虫及其幼虫和土壤动物的栖息地[34-35],不但能为以上3 种鸟类集团提供取食场所,而且是大多数食虫鸟类和杂食性鸟类的食物来源地。在本研究中林下带作为鸟类取食场所的特性并不突出,只有3/8的鸟类集团以此为取食场所,因此在公园次生林的植被配置过程中,应提高林下带灌木和草本植物的盖度和多样性,增加昆虫的保有量为更多鸟类提供食物来源。
(3)灌木层为食虫拾取集团和植食拾取集团提供运动场所,为杂食拾取集团提供取食场所。多数鸟类集团未选择灌木层作为其运动和取食场所。然而,鸟类栖息行为主要在乔木层出现而其取食行为主要在地被层出现(图1),从乔木层到地被层或者逆向的鸟类活动必须经过灌木层才能实现,因此灌木层的屏障作用尤为明显。Camprodon和Brotons[19]和Hansson[36]已经证明了灌木层能够为鸟类提供筑巢的场所和庇护的屏障,从而能够显著增加鸟类的多样性水平和多度。而在瑞典城市绿地的研究表明,去除90%灌木层的人工林的鸟类多样性和多度显著少于未去除灌木的人工林[37]。因此灌木层作为重要的植被结构,在城市绿地中对鸟类群落的生态作用非常重要。仅有2/8的鸟类集团利用其作为运动场所,因此灌木层植被种植过程中,应增加灌木层盖度为鸟类群落提供屏障。
3.2 鸟类集团空间生态位的重叠与分离
对比8 种鸟类集团在植被层次上的行为频次(图3)和相互间空间生态位重叠度(表3)进行比较,结果表明鸟类集团的之间存在空间生态位的重叠和分离。
以食虫拾取集团、杂食拾取集团、食肉飞取集团和植食拾取集团为例。其在栖息、运动和取食空间生态位上均存在较高的重叠度(表3)并且以上4 种集团鸟类多度占到全部陆生鸟类多度的94.54%,说明其通过食性分离了各自的空间生态位[38]。由于以上4 种取食方式的鸟类集团在空间生态位上存在重叠,因此,一旦某一集团的鸟类数量下降或消失,另一个能够利用这一空间或食物的集团的鸟类数量就会明显上升来填补鸟类数量下降所带来的生态位空缺,这也是城市化导致鸟类多样性下降但优势鸟种密度升高的原因之一。Clergeau[30]和Sandstrom[5]的研究表明,城市化导致大山雀(Parusmajor),戴菊(Regulusregulus),虎斑地鸫(Zootheradauma)和白喉矶鸫(Monticolagularis)的数量显著下降而树麻雀(Passermontanus),乌鸫(Turdusmerula)和灰喜鹊(Cyanopicacyana)的数量显著升高。通过对比分析发现,数量下降的多为食虫拾取鸟类而数量上升的多为杂食拾取鸟类,这一结果与本文结论一致。
食虫探取集团和食虫飞取集团互为栖息空间生态位重叠度最高集团,而其食性又同为食虫,因此两者避免激烈竞争的原因之一可能源于取食方式的不同。食虫探取集团以啄破或爪穿破树干,枯枝等搜寻方式取食,因此其取食位置具有随机性;食虫飞取集团以在树枝间来回跳跃或飞翔的方式取食[24]。两者避免激烈竞争的另一个原因可能是昆虫资源非常充足,食虫飞取集团中5种鸟类均为候鸟,在滨江森林公园均为5—10 月观察到的,这个时期恰好是昆虫的爆发期,食物资源相对丰富,相关研究结果见王勇等[25]。
3.3 城市公园作为鸟类栖息地生态服务功能的建议
上海市位于澳大利亚西部-东西伯利亚鸟类迁徙路线的中点,对上海鸟类资源的保护,不仅对维护城市生态系统起着重要作用,而且保护了整个迁徙路线上的鸟类资源。城市公园作为城市陆生鸟类重要的栖息地,其生态服务功能不言而喻。从植被角度来看,公园非常重视乔木植物的种植与搭配,5/8的鸟类集团以其作为主要的栖息场所,还有部分鸟类以其作为运动和取食的场所。而灌木植物和草本植物的种植和搭配往往不能满足多数鸟类集团的要求。从城市公园作为陆生鸟类栖息地的生态服务功能考虑,应增加灌木植物的密度和盖度,增加草本植物的多样性,减小人工草坪的种植面积或者将人工大草坪分割为若干小块。但是从公园作为游客休闲场所的角度考虑,显然高密度的灌木容易使游客产生恐惧感[37],因此建议通过植物分区的方式来实现公园功能区域的划分[5,39],从而为野生鸟类提供较理想的栖息地。从鸟类集团的角度出发,不同鸟类集团对植被层次和结构的要求存在差异。要提高鸟类集团的多样性水平,必须配置不同结构的林型。例如,对于食虫拾取集团,植被3 个层次均不可或缺,应配置较为分层结构复杂的林型;对于食虫探取集团应当根据其取食方式配置易于探取的树种;对于食虫飞取集团,应配置盖度较大的灌木层为其提供栖息和觅食的场所。
致谢:滨江森林公园全体工作人员给予大力支持,李宏庆老师,毕玉科,周青梅,何娅,杨会涛,刘旭,钟宇,刘晶,路珊帮助写作,特此致谢。
[1] Grimm N B, Faeth S H, Golubiewski N E, Redman C L, Wu J G, Bai X. M, Briggs J M. Global change and the ecology of cities. Science, 2008, 319(5864):756-760.
[2] Padoa-Schioppa E, Baietto M, Massa, R, Bottoni. L. Bird communities as bioindicators: The focal species concept in agricultural landscapes. Ecological Indicators, 2006, 6:83-93.
[3] Goode, D. A. Urban nature conservation in Britain. Journal of Applied Ecology, 1989: 859-873.
[4] 陈水华, 丁平, 郑光美, 诸葛阳. 城市鸟类群落生态学研究展望. 动物学研究, 2000, 21(2): 165-169.
[5] Sandstrom U G, Angelstam P, Mikusinski G. Ecological diversity of birds in relation to the structure of urban green space. Landscape and Urban Planning, 2006, 77(1/2):39-53.
[6] Root R B. The niche exploitation pattern of the blue-gray gnatcatcher. Ecological Monographs, 1967, 37(4):317-350.
[7] Landres P B, MacMahon J A. Guilds and community organization: analysis of an oak woodland avifauna in Sonora, Mexico. The Auk, 1980: 351-365.
[8] 蔡音亭, 唐仕, 袁晓, 王吉衣,马志军. 上海市鸟类记录及变化. 复旦学报(自然科学版), 2011, 50 (3): 334-343.
[9] 栾晓峰. 上海鸟类群落特征及其保护规划研究[D]. 上海: 华东师范大学, 2003.
[10] Davids A M, Glick T F. Urban ecosystems and island biogeography. Environmental Conservation, 1978, 5: 299-304.
[11] Jokimäki J. Occurrence of breeding bird species in urban parks: effects of park structure and broad-scale variables. Urban Ecosystems, 1999, 3(1): 21-34.
[12] Gavareski G A. Relation of park size and vegetation to urban bird populations in Seattle, Washinton. The Condor, 1976, 78: 375-382.
[13] Robinson S K, Wilcove D S. Forest fragmentation in the temperate zone and its effects on migratory songbirds. Bird Conservation International, 1994, 4: 233-249.
[14] Mehlman D W, Mabey S E, Ewert D N, Duncan C, Abel B, Cimprich D, Sutter R D. Woodrey M. Conserving stopover sites for forest-dewelling migratory landbirds. The Auk, 2005, 122: 1281-1290.
[15] Fontana S, Sattler T, Bontadina F, Moretti M. How to manage the urban green to improve bird diversity and community structure. Landscape and Urban Planning, 2011, 101: 278-285.
[16] 葛振鸣, 王天厚, 施文彧, 周立晨, 薛文杰. 环境因子对上海城市园林春季鸟类群落结构特征的影响. 动物学研究, 2005, 26: 17-24.
[17] 徐骁俊, 葛振鸣, 裴恩乐, 施文彧, 王正寰, 王天厚. 上海世界博览会园区内及周边地区鸟类多样性及其影响因子. 生态学杂志, 2007, 26: 1954-1958.
[18] Degraaf R M, Wentworth J M. Avian Guild Structure and Habitat Associations in Suburban Bird Communities. Urban Ecology, 1986, 9(3/.4): 399-412.
[19] Camprodon J, Brotons L. Effects of undergrowth clearing on the bird communities of the Northwestern Mediterranean Coppice Holm oak forests. Forest Ecology and Management, 2006, 221(1/3): 72-82.
[20] Shannahan D F, Miller C, Possingham H P, Fuller R A. The influence of patch area and connectivity on avian communities in urban revegetation. Biological Conservation, 2011, 144: 722-729.
[21] 葛振鸣, 王天厚, 周晓, 赵平, 施文彧. 上海崇明东滩堤内次生人工湿地鸟类冬春季生境选择的因子分析. 动物学研究, 2006, 27(2): 144-150.
[22] 任梦非, 朱祥明. 上海滨江森林公园规划设计研究. 中国园林, 2007, 2: 21-27.
[23] 万福绪, 董波, 陈敏. 上海滨江森林公园景观空间格局分析. 中国园林, 2008, 7: 61-65.
[24] 赵正阶. 中国鸟类志. 长春: 吉林科学技术出版社, 2001.
[25] 王勇, 许洁, 杨刚, 李宏庆, 吴时英, 唐海明, 马波, 王正寰. 城市公共绿地常见木本植物组成对鸟类群落的影响. 生物多样性, 2014, 22(2): 196—207.
[26] 刘菡, 韩联宪. 云南哀牢山徐家坝常绿阔叶林的鸟类取食集团. 动物学研究, 2008, 29 (5): 561-568.
[27] 刘彬, 周立志, 汪文革, 沈三宝, 韩德民. 大别山山地次生林鸟类群落集团结构的季节变化. 动物学研究, 2009, 30 (3): 277-287.
[28] Diaz L. Influences of forest type and forest structure on bird communities in oak and pine woodlands in Spain. Forest Ecology and Management, 2006, 223(1/3): 54-65.
[29] Horn H S. Measurement of “overlap” in comparative ecological studies. American Naturalist, 1966: 419-424.
[30] Clergeau P, Savard, J P L, Mennechez G, Falardeau G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor, 1998, 100: 413-425.
[31] Palomino D, Carrascal L M. Urban influence on birds at a regional scale: A case study with the avifauna of northern Madrid province. Landscape and Urban Planning, 2006, 77(3): 276-290.
[32] Evans K L, Newson S E, Gaston K J. Habitat influences on urban avian assemblages. Ibis, 2009, 151: 19-39.
[33] Pearman P B. The scale of community structure: Habitat variation and avian guilds in tropical forest understory. Ecological Monographs, 2002, 72(1):19-39.
[34] 吴时英. 城市森林病虫害图鉴. 上海:上海科学技术出版社, 2005.
[35] 刘继亮, 曹靖, 李世杰, 潘春林, 潘成臣. 秦岭西部山地次生林和人工林大型土壤动物群落结构特征. 应用生态学报, 2012, 23(9): 2459-2466.
[36] Hansson L. Traditional management of forests: plant and bird community responses to alternative restoration of oak-hazel woodland in Sweden. Biodiversity and Conservation, 2001, 10(11):1865-1873.
[37] Heyman E. Clearance of understory in urban woodlands: Assessing impact on bird abundance and diversity. Forest Ecology and Management, 2010, 260(1):125-131.
[38] Holmes R, Bonney J R, Pacala S. Guild structure of the Hubbard Brook bird community: a multivariate approach. Ecology, 1979: 512-520.
[39] Gordon A, Simondson D, White M, Moilanen A, Bekessy S A. Integrating conservation planning and landuse planning in urban landscapes. Landscape and Urban Planning, 2009, 91(4):183-194.
The influence of vegetation structure on bird guilds in an urban park
YANG Gang1, 2, XU Jie1, 2, WANG Yong1, 2, DING Youzhong1,2, YUAN Xiao3, PEI Enle3, MA Bo3, WANG Xiaoming4, WANG Zhenghuan1,2,*
1SchoolofLifeSciences,EastChinaNormalUniversity,Shanghai200062,China2ShanghaiKeyLaboratoryofUrbanizationandEcologicalRestoration,EastChinaNormalUniversity,Shanghai200241,China3DepartmentofWildlifeProtectionandManagementAdministration,Shanghai200023,China4ShanghaiScienceandTechnologyMuseum,Shanghai200127,China
The acceleration of urbanization has led to a loss of biodiversity, which is a global ecological problem of great concern. Meanwhile, birds are useful indicators in monitoring environmental changes in urban ecosystem. Urban green areas have always been paid great attention because of their functions as natural habitats for many wildlife species. Bird guilds are groups of species that use environmental resources in similar ways. Therefore, we analyze the function of vegetation structure to bird community and bird guilds so that we make suggestions related to the design of urban green spaces in Shanghai. From October 2009 to October 2011, we set the line transect to survey the bird community in Shanghai Binjing Forest Park. According to the height of plant, we divided the habitat vegetation into three vertical levels including tree layer, shrub layer and ground layer. Chi-square test showed that there were significant differences among behaviors in three vegetation layers. The frequency of perching and moving in tree layer was more significant than that in other two layers and the frequency of feeding in ground layer was more significant than that in other two layers. According to the variables based on the location, feeding habits and foraging behavior pattern of birds, we classified 8 bird guilds including herbivorous collecting, herbivorous pecking, insectivorous collecting, insectivorous flying catching, insectivorous pecking, omnivorous collecting, omnivorous pecking and raptorial flying catching. There were differences and overlaps among the frequency of three behaviors of 8 bird guilds in three layers, so we considered there was niche overlap among 8 bird guilds. According to Morisita-Horn Index, the results indicated that insectivorous collecting guild, omnivorous collecting guild, flying catching guild and herbivorous collecting guild had high niche overlap of perching, and moving and feeding so these guilds reached niche separation by their diets. Insectivorous pecking guild and insectivorous flying catching guild had the most perching niche overlap with each other, and they reached niche separation by their foraging behavior patterns. Based on the results above, we concluded that trees were one of the most important vegetation components that enhance the bird species richness and diversity in urban green spaces because they provide roosting, nesting, hiding, and foraging sites. The shield function of shrub cover was confirmed to be an important habitat component and the ground layer provided food resources to bird community. The suggestion and recommendation in construction of urban parks was presented based on this study. The density and coverage of shrub layer should improve and large area of artificial lawn should be divided into several small pieces. We also discuss the conflicts between habitat for biodiversity maintenance and recreational function in urban parks.
vegetation structure; bird guilds; niche overlap; principal component analysis; bird behavior
上海市绿化和市容管理局专项基金; 浦东新区科技发展基金(PKJ2011-N020); 上海市城市化生态过程与生态恢复重点实验室开放课题基金
2013-11-04;
2014-09-09
10.5846/stxb201311042667
*通讯作者Corresponding author.E-mail: zhwang@bio.ecnu.edu.cn
杨刚, 许洁, 王勇, 丁由中, 袁晓, 裴恩乐, 马波, 王小明, 王正寰.城市公园植被特征对陆生鸟类集团的影响.生态学报,2015,35(14):4824-4835.
Yang G, Xu J, Wang Y, Ding Y Z, Yuan X, Pei E L, Ma B, Wang X M, Wang Z H.The influence of vegetation structure on bird guilds in an urban park.Acta Ecologica Sinica,2015,35(14):4824-4835.
猜你喜欢
知识窗(2022年8期)2022-09-08
幼儿园(2021年2期)2021-05-25
红领巾·探索(2020年9期)2020-10-26
小学生学习指导(低年级)(2019年9期)2019-09-25
乡村科技(2018年8期)2018-02-13
小学生学习指导(低年级)(2018年3期)2018-01-31
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
台声(2016年16期)2016-11-25
甘肃林业(2016年4期)2016-11-07
