
中华水韭孢子囊发育研究
2015-07-05 07:02连永权刘保东
西北植物学报 2015年5期
李 婧,连永权,李 范,刘保东*
(1 哈尔滨师范大学 黑龙江省普通高等学校植物生物学重点实验室,哈尔滨150025;2 黑龙江职业学院,哈尔滨150111)
水韭科(Isoetaceae)是古老的孑遗拟蕨类水生植物,全球零散分布约250种,多为珍稀濒危保护物种[1],中国仅5种水韭,且均为特产种[2]。由于生存环境不断恶化,野生数量减少,全科均被列为国家一级保护物种[3]。中华水韭(Isoetes sinensis)曾是长江中下游湿地常见蕨类,对自然环境变化较为敏感[4],随着近几十年该地区经济活动的快速发展,数量急剧下降,已列为国家极危级(CR)保护物种[5]。He通过细胞学研究指出减数分裂过程中染色体的行为变化可能是有性生殖的限制因素,但不是种群数量下降的主要原因;陈进明等[7]就中华水韭的遗传多样性进行分析,认为遗传结构是限制种群发展的主要成因之一;中华水韭对温度这一古老因素具有抗性[8],对现代常用除草剂的抗性较低[9];邢建娇等[10]通过人工繁殖和栽培指出培养基质是其濒危的重要原因;李范[11]提出中华水韭在发育幼期是有缘膜存在的,随着植株生长而逐渐消失。迄今为止,尚未有中华水韭孢子囊发育过程以及发育特征与濒危关系等方面的相关报道。
孢子囊作为蕨类植物无性生殖的唯一场所,蕴涵着大量保守而稳定的系统学信息[12],分子生物学也认为孢子囊是蕨类植物重要的溯祖形态[13]。真蕨亚门的经典分类包括厚囊蕨纲、薄囊蕨纲和原始薄囊蕨纲,孢子囊是分纲的主要依据;不少目或科的分类也主要依据孢子囊的特征,如着生方式、环带的有无、开裂方式等[14]。就卷柏属(Selaginella)而言,孢子囊也有重要的系统学和生殖生物学参考价值,刘保东等在研究24种卷柏的孢子囊排布规律后指出,大小孢子囊的着生位置和数量既能反映出物种间的亲缘关系,又与有性生殖能力有关[15]。
水韭科(Isoetaceae)与卷柏科(Selaginellaceae)同属石松亚门(Lycophytina)。两者大孢子囊和小孢子囊分别着生在不同叶片的基部[13],水韭偶尔还会出现混生型孢子囊[16],叶舌的结构也远比卷柏类复杂[11],孢子囊的发育特征应具有较高的系统学研究价值[17];国产水韭中,中华水韭濒危程度最高,尽管濒危原因尚不明确,但孢子囊的发育直接决定着孢子的数量和质量[18],是中华水韭无性生殖的关键环节。本实验用半薄切片法,连续观察了中华水韭大小孢子囊的发育过程,从无性生殖的角度为探讨其濒危原因提供直观可靠的理论证据,也为研究水韭类的亲缘关系提供保守而稳定的溯祖形态。
1 材料和方法
实验材料中华水韭(Isoetes sinensis)原产浙江省华顶山,由浙江大学丁炳扬教授采集并赠送活体植株,经刘保东[19]人工繁殖并栽培在哈尔滨师范大学蕨类培养室(126°42′E,45°24′N)。取个体发育不同阶段的人工繁殖栽培苗,在解剖镜下检查孢子囊的发育情况以及与叶片的关系。切取不同发育程度的孢子囊,用FAA 固定并经过常规半薄切片,在Olympus BH-2型光学显微镜下观察并拍照。
将成熟且完好的小孢子囊,放在10 mL 量筒内,滴加约5 mL 蒸馏水后,在水中将小孢子囊挑破,取出孢子囊碎片,定容至10 mL 并摇动成均匀悬浊液,吸取1mL悬浊液再稀释至10mL,以此连续稀释数次后,用移液器吸取0.01mL 滴加在载玻片上,置显微镜记录全部小孢子的数量后,换算出小孢子囊中小孢子的总数。将成熟且完好的大孢子囊直接置于载玻片的一滴蒸馏水中,挑破孢子囊壁,直接记录大孢子的数量。大小孢子囊的孢子产量均取随机检测10个数值的平均值。
2 观察结果
2.1 大小孢子囊的发育节律及其孢子产量
中华水韭在个体发育初期的1~20枚叶片上,均无孢子囊着生,约20枚叶片后,每枚叶片的基部近轴面均着生1个孢子囊,每个孢子囊内只发育出大孢子或小孢子,大小孢子异囊异叶,大小孢子叶相间排列(图版Ⅰ,1)。在人工培养条件下,孢子囊随孢子叶持续发生,不受自然条件的季节限制,在相应的叶片伸长生长过程中完成减数分裂,并未发现同时发育有大小孢子的混生孢子囊。
中华水韭的两种孢子囊在形态大小上都基本相同(图版Ⅰ,2),小孢子囊幼嫩时为浅黄色,成熟后暗灰色(图版Ⅰ,3),小孢子的平均产量是5.36×105个;大孢子囊幼嫩时为浅黄绿色(图版Ⅰ,4),成熟后灰色(图版Ⅰ,3),大孢子的平均产量是408个。
2.2 大小孢子囊的早期分化
当中华水韭的个体从合子发育到约20枚叶片的幼植物体时,以后分化的每个叶片,其基部都发育出一个孢子囊,孢子囊的分化稍迟于叶原基的分化。当每个叶原基的高度发育到2mm 左右时,孢子囊几乎与叶片的维管束同步分化,孢子囊位于维管束的近轴面(图版Ⅰ,5),稍呈截球形隆起,孢子囊的分裂能力较为旺盛,着色略深于叶片组织,此时的大小孢子囊在形态结构上都很简单,没有明显的差别。当叶片发育至1cm 长短时,孢子囊随着叶片的伸长也向上下延长,呈椭球形与叶片组织紧密相连,且向外明显突起,但内部的细胞还没有明显的分化(图版Ⅰ,6)。其后孢子囊的突起速度显著慢于上下延长的速度,而且孢子囊隆起部分向周围扩展,孢子囊与叶片相连的面积显著小于其表面积(图版Ⅰ,7)。当叶片长度达到3cm 以上后,孢子囊在快速隆起的同时,内部细胞也发生着明显的分化,横隔丝原始细胞及孢子母细胞的细胞质浓度不断加深(图版Ⅰ,8),但大小孢子囊的区别仍不明显。
2.3 小孢子囊的发育
当叶片长度达到10cm 以上时,孢子囊内染色较深的区域进一步加深,大小孢子囊的特征逐渐显现,首先会出现十多条横隔丝及小孢子分生组织(图版Ⅰ,9),最初的横隔丝较宽、着色较深,细胞排列密集,其后,一些横隔丝出现岛屿化溶解(图版Ⅰ,10),小孢子母细胞的减数分裂几乎是同步进行,半数以上的小孢子囊败育,所以产生的小孢子颜色较浅,胞质透明(图版Ⅰ,11),近半数的小孢子囊内虽然产生了不少颜色较深、胞质浓密的小孢子,但隔丝较粗壮(图版Ⅰ,12)。在正常发育的小孢子囊内,小孢子母细胞分裂旺盛,能迅速发育成四分体,而且远轴侧的孢子囊壁较厚且不规则(图版Ⅰ,13),近轴侧的孢子囊壁与叶片维管束融为一体(图版Ⅰ,14)。四分体密集,细胞核较大,着色较深(图版Ⅰ,15),当叶片长度达到20cm 以上且不再伸长后,小孢子囊内四分体的中壁和外壁(周壁)才不断形成(图版Ⅰ,16)。小孢子囊无柄,也无明显的开裂结构。
2.4 大孢子囊的发育
在叶片长度达到10cm 以上时,大孢子囊内先出现少数横隔丝(图版Ⅰ,17)。大孢子母细胞为不整齐发育,即一些大孢子四分体已经形成,一些大孢子母细胞还在减数分裂之中(图版Ⅰ,18)。四分体各具有1个细胞核,胞质染色较深(图版Ⅰ,19)。有近半数的大孢子囊败育,整个孢子囊内除少数分化不完全的隔丝外,其余区域均近乎透明(图版Ⅰ,20)。在近半数正常发育的大孢子囊内,多数大孢子的四分体呈圆形(图版Ⅰ,21),随着大孢子的发育,内含物不断充实,三层孢子壁分化明显(图版Ⅰ,22)。也有少数大孢子囊外观上与正常发育的基本相似(图版Ⅰ,23),但其内部的大孢子多数败育,即孢子壁内陷,内含物较少(图版Ⅰ,24)。大孢子囊的减数分裂发生在叶片伸长生长过程中,当叶片停止生长后,孢子壁才逐渐分化,败育的大孢子囊要么萎缩干瘪,要么基本类似于正常大孢子囊。大孢子囊无柄,也无明显的开裂结构,从切片上观察,在可育的大孢子囊中,隔丝较粗,所占体积比例要明显大于小孢子囊。
3 讨 论
孢子囊既是现代蕨类植物学重要的分类原则之一,也是蕨类植物进行无性生殖的唯一场所[12],孢子囊的孢子产量一直是蕨类植物生殖方式的重要参考依据 。叶创兴等 提出,水韭大、小孢子囊内孢子的个数相差1 000倍,刘星[22]的统计显示中华水韭大小孢子的直径基本相差10倍,我们观察到中华水韭两种孢子囊的体积基本相等,这些似乎都支持叶创兴的观点。然而事实上,隔丝在可育大孢子囊内所占的体积较大,在小孢子囊内则相反,因此,小孢子的数量一定大于大孢子1 000倍,这一结论得到了本研究统计结果的支持。
幼期的小孢子囊有较粗的隔丝,以保证造孢组织的有丝分裂[17]。但小孢子囊内的十多万个小孢子母细胞同时进行减数分裂,此时若环境稍有改变,就很容易破坏小孢子的营养供给从而造成败育。中华水韭的孢子囊都是在叶片伸展时期即产地的初春季节进行减数分裂,而该季节寒潮和干旱多发[23]。笔者认为,环境因素引起的小孢子囊败育应该是导致中华水韭濒危的一个重要原因。尽管大孢子母细胞的不整齐发育避开了偶然环境因素所导致的败育环节,但近半数的隔丝一开始就发育不良,从根本上限制了大孢子母细胞的减数分裂,其必然性败育的致危程度要大于小孢子囊的偶然性败育。
笔者认为多数植物学教科书[17]关于水韭属植物最外围是不育叶,向内依次为大孢子叶和小孢子叶以及尚未分化的幼叶,这一描述存在不妥。据笔者观察,中华水韭为多年生草本植物,既然存在大小孢子叶,“最外轮的叶片”也一定是可育叶,因为幼孢苗时期的前几十枚叶片很早就已脱落[10]。外轮叶片不断随其孢子囊的过熟而不断脱落,内部的孢子叶则逐渐由内轮变成外轮,“外轮是大孢子叶”的结论只能是特殊时期的静态描述。从生殖生态学的角度看,如果外一轮多枚叶片都具有相同的大孢子囊,那么这些大孢子囊间必然存在分化时间和成熟程度的不同,而导致内一轮叶片的小孢子囊远不能及时分化和成熟,以致产生严重的雌雄不遇后果,进而影响到有性生殖。观察得出,中华水韭的大小孢子叶不存在外轮内轮的单一孢子囊现象,大小孢子叶基本是随机的相间排列。尽管单个小孢子囊同时成熟,单个大孢子囊不同时成熟,而每株中华水韭在一个生长季可以成熟多枚大小孢子囊,因此,中华水韭不存在生殖演化上的缺陷,只是小孢子囊对环境的依赖性较强,且大孢子囊的败育比例过高。
水韭的近缘植物石松属(Lycopodium)和卷柏属(Selaginella),多为旱生,孢子能借风力进行远距离传播,孢子囊都有固定的开裂结构和方式[18];卷柏属的孢子囊还能通过基部细胞失水收缩而帮助孢子释放 ,多数卷柏的大孢子囊都位于孢子叶穗的基部,易于被动受精[15],具有较高的繁殖能力。然而,水韭属的大小孢子囊都不存在开裂结构,也没有固定的开裂方式,始终生长在泥水里,只能通过孢子叶基部破裂或腐烂而释放孢子[17],这也应该是导致中华水韭濒危的原因之一。
[1] ZHANG L B,W C TAYLOR.Isoetaceae[M]//Flora of China Editorial Committee.Flora of China.St.Louis,Missouri,USA:Missouri Botanical Garden Press,2013,2-3:35-36.
[2] 印 红.中国珍稀濒危植物图鉴[M].北京:中国林业出版社,2013:32-36.
[3] 中华人民共和国国务院.国家重点保护野生植物名录(第一批)[J].植物杂志,1999,(5):3-11.
[4] LIU X,GITURU W R,WANG Q F,et al.Distribution of basic diploid and polyploidy species of Isoetesin East Asia[J].Journal of Biogeography,2004,31:1 239-1 250.
[5] 严岳鸿,张宪春,马克平.中国蕨类植物多样性与地理分布[M].北京:科学出版社,2013:76-218.
[6] HE Z C,WANG H C,LI J Q,et al.Chromosome behavior during meiosis and development of spore mother cells in the Chinese quillwort Isoetes sinensis(Isoetaceae)[J].American Fern Journal,2004,94(4):183-195.
[7] CHEN J M(陈进明),LIU X(刘 星),WANG Q F(王青锋).Genetic diversity in Isoetes yunguiensis,a rare and endangered fern in China[J].Journal of Wuhan University(武汉大学学报),2005,51(6):767-770(in Chinese).
[8] LIU T T(刘婷婷),SUN H(孙 昊),GUAN Y(关 旸),et al.Effects of low temperature stress on physiological characters of critically endangered Isoetes sinensis Palmer[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2013,33(10):2 031-2 036(in Chinese).
[9] SUN H(孙 昊),LIU T T(刘婷婷),GUAN Y(关 旸),et al.Physiological response to herbicides of Isoetes sinensis Palmer critically endangered ferns[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2013,33(9):1 830-1 837(in Chinese).
[10] XING J J(邢建娇),LU J(路 靖),LI F(李 范),et al.Spore seedling cultivation and care of young sporophyte seedling of extreme endangerous wetland plant Isoetes sinensis[J].Wetland Science(湿地科学),2013,11(3):347-351(in Chinese).
[11] LI F(李 范),LIU J H(刘婧宏),XING J J(邢建娇),et al.Research on the development of ligule and velum in Isoetes sinensis Palmer[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2013,33(1):17-21(in Chinese).
[12] 吴兆洪,秦仁昌.中国蕨类植物科属志[M].北京:科学出版社,1991:114-120.
[13] ZHANG X CH(张宪春),WEI R(卫 然),LIU H M(刘红梅),et al.Phylogeny and classification of the extant lycophytes and ferns from China[J].Chinese Bulletin of Botany(植物学报),2013,48(2):119-137(in Chinese).
[14] XIAO X Y,LIN R C,CHEN X D.The comparative studies on microspores of fourteen species in Selaginella[J].Chinese Journal of Pharmaceutical Analysis,2002,22(1):20-24.
[15] 刘保东,包文美,敖志文,等.卷柏科孢子囊排布规律的系统学意义初步研究[C]//中国青年科学技术论文精选.北京:中国科学技术出版社,1994:1 033-1 037.
[16] SINGH R,BOHRA D R,SHARAM B D.Mixed sporangia in Isoetes coromandelina L.structure,systematics[J].Pollen et Spores,1983,25:41-47.
[17] P C VASHISHTA.Botany for degree students:Pteridophyta[M].New Delhi:S.Chand and Company LTD,1997:119-141.
[18] TOM A R,CHRISTOPHER H H.Biology and Evolution of Ferns and Lycophytes[M].The USA:Cambridge University Press,2008:61-74.
[19] LIU B D(刘保东).Artificial batch reproductive success of Isoetes sinensis,a critically endangered species[J].Biodiversity Science(生物多样性),2012,20(6):784(in Chinese).
[20] NAKATO N,KATO M,LIU B D.A cytotaxonomic study of some ferns from Jiangsu and Zhejiang Provinces China[J].The Journal of Japanese Botany,1995,70:194-204.
[21] 叶创兴,朱念德,廖文波,等.植物学[M].北京:高等教育出版社,2007:296.
[22] LIU X(刘 星),LIU H(刘 虹),WANG Q F(王青锋).Spore morphology of Isoetes(Isoetaceae)from China[J].Journal of Systematics and Evolution(植物分类学报),2008,46(4):479-489(in Chinese).
[23] YU L(於 琍),NATIONAL C C.Assessment on ecosystem vulnerability to droucht——The case study of the middle and lower reaches of the Yangtze River[J].Resources and Environment in the Yangtze Basin(长江流域资源与环境),2014,23(7):1 063-1 070(in Chinese).
[24] JAKOB S,HANS G,ALEX Z.Speed and force of spore ejection in Selaginella martensii[J].Alpine Botany,2008,118(1):13-20.
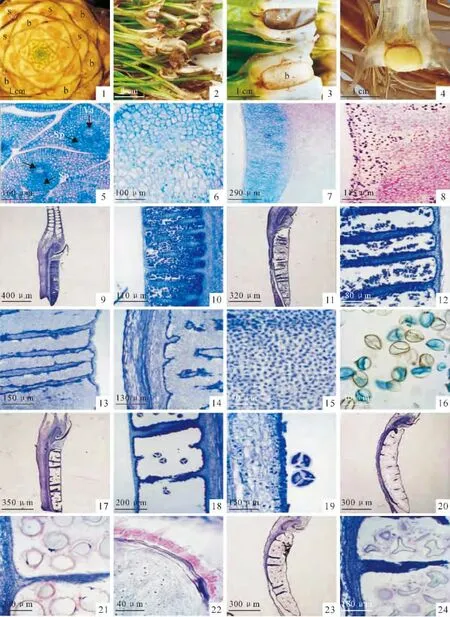
图版Ⅰ 中华水韭大小孢子囊(b.大孢子囊;s.小孢子囊;Sp.孢子囊;Va.维管束)1.大孢子囊与小孢子囊在茎端的排布;2.成熟的孢子囊;3.发育良好的大孢子囊与小孢子囊;4.幼小的大孢子囊;5.孢子囊与叶原基的维管束;6.孢子囊向外隆起;7.孢子囊向上下延展;8.孢子囊内部开始分化;9.小孢子囊的横隔丝及分生组织;10.横隔丝岛屿化溶解;11.败育的小孢子囊;12.具粗壮隔丝的小孢子囊;13.正常发育的小孢子囊及其远轴侧囊壁;14.正常发育的小孢子囊及其近轴侧囊壁;15.小孢子囊内的四分体;16.成熟中的小孢子;17.正常发育的大孢子囊;18.四分体形成中;19.大孢子四分体及造孢组织;20.败育初期的大孢子囊;21.可育的大孢子囊;22.正在成熟的大孢子;23.部分败育的大孢子囊;24.败育的大孢子囊。PlateⅠ Sporangium of I.sinensis(b.Macrosporangium;s.Microsporangium;Sp.Sporangium;Va.Leaf primordium)Fig.1.Distribution of macrosporangium and microsporangium on the stem end;Fig.2.Mature sporangium;Fig.3.Well developed macrosporangium and microsporangium;Fig.4.Young macrosporangium;Fig.5.Sporangium and vascular bundle of leaf primordium;Fig.6.Sporangium hunch out;Fig.7.Sporangium is expanding;Fig.8.Internal structure differentiation in sporangium;Fig.9.Trabecula and meristem of microsporangium;Fig.10.Trabecula is dissolving;Fig.11.Abortive microsporangium;Fig.12.Microsporangium with thick trabecula;Fig.13.Normal developed microsporangium and its abaxial wall;Fig.14.Normal developed microsporangium and its adaxial wall;Fig.15.Tetrad in microsporangium;Fig.16.Mature microsporangium;Fig.17.Normal developed megasporangium;Fig.18.Tetrad forming;Fig.19.Tetrad of macrospore and sporogenous tissue;Fig.20.Megasporangium in early abortion;Fig.21.Fertile megasporangium;Fig.22.Developing megaspore;Fig.23.Partly abortive megasporangium;Fig.24.Abortive megasporangium.
猜你喜欢
西北植物学报(2022年4期)2022-05-24
科学大众·教师版(2022年6期)2022-05-23
北京航空航天大学学报(2021年9期)2021-11-02
现代农业科技(2020年21期)2020-12-14
生物学通报(2019年7期)2019-07-08
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年9期)2018-10-08
江苏农业科学(2016年4期)2016-06-14
农业研究与应用(2014年1期)2014-08-15
作文与考试·高中版(2008年2期)2008-03-03
