
Targeting neuronal nitric oxide synthase as a valuable strategy for the therapy of neurological disorders
2016-02-09 05:17CristinaMaccalliniRosaAmorosoDepartmentofPharmacyUniversityAnnunzioofChietiPescaraChietiItaly
中国神经再生研究(英文版) 2016年11期
Cristina Maccallini, Rosa AmorosoDepartment of Pharmacy, University “G. d’Annunzio” of Chieti-Pescara, Chieti, Italy
Targeting neuronal nitric oxide synthase as a valuable strategy for the therapy of neurological disorders
Cristina Maccallini*, Rosa Amoroso
Department of Pharmacy, University “G. d’Annunzio” of Chieti-Pescara, Chieti, Italy
How to cite this article:Maccallini C, Amoroso R (2016) Targeting neuronal nitric oxide synthase as a valuable strategy for the therapy of neurological disorders. Neural Regen Res 11(11):1731-1734.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
The management of neurological disorders have huge and increasing human and economic costs. Despite this, there is a scarcity of effective therapeutics, and there is an extreme urgency for new and real treatments. In this short review we analyze some promising advancements in the search of new bioactive molecules targeting neuronal nitric oxide synthase (nNOS), an enzyme deputed to the biosynthesis of nitric oxide (NO). In different conditions of neuronal damages, this molecule is overproduced, contributing to the pathogenesis and progression of neuronal diseases. Two main approaches to modulate nNOS are discussed: a first one consisting in the direct inhibition of the enzyme by means of small organic molecules, which can be also active against other different targets involved in such diseases. A second section is dedicated to molecules able to prevent the formation of the ternary complex N-methyl-D-aspartate (NMDA)-type glutamate receptors, postsynaptic density-95 (PSD95) protein-nNOS, which is necessary to activate the latter for the biosynthesis of NO.
amidines; carbonic anhydrase; inhibitors; neurological diseases; neuronal nitric oxide synthase; NMDAR; PSD95
Introduction
A growing number of studies demonstrate that an abnormal nitrergic signaling in the central nervous system (CNS) is a crucial event in the development of neurological disorders, such as ischemic stroke, migraine, acute brain and spinal cord injury,etc. (Yuste et al., 2015; Antonaci et al., 2016; Wang et al., 2016). The free radical nitric oxide (NO) is a signaling molecule ubiquitously biosynthesized by the nitric oxide synthases (NOS) family, in a two step reaction involving oxygen and many cofactors, such as nicotinamide adenine dinucleotide phosphate (NADPH), flavins and tetrahydrobiopterin. Three NOS isoforms are currently known: the neuronal and endothelial ones are constitutive (nNOS and eNOS, respectively), while the inducible isoform (iNOS) is expressed in response to invading pathogens, or pro-inflammatory stimuli. All of the NOSs have essential physiological roles in the CNS, and their activity is compromised in different neurological states. However, among NOS isoforms, the nNOS is the most implicated in a wide range of functions and pathologies with pleiotropic effects. In the last years, a growing number of molecules able to modulate nNOS activity have appeared, and in this review we focus on nNOS as a possible therapeutic target for neuroregeneration and recovery of neuronal functions.
In the CNS, the nNOS is localized inside the postsynaptic cell membrane and is physically tethered to N-methyl-D-aspartate (NMDA)-type glutamate receptors, through postsynaptic density-95 (PSD95) protein. Under physiological conditions, mild activation of ‘synaptic’ NMDARs allows the infux of Ca2+, which, binding to calmodulin, leads to nNOS catalytic activation. The produced NO exerts different cellular effects, mainly by nitrosylation of the ferrous heme of the metalloenzyme soluble guanylate cyclase, activating the cyclic guanosine monophosphate (cGMP) signaling. Moreover, the reaction of NO and its derived species (N2O3, NO2–, NO+,etc.) with protein cysteine and tyrosine residues, generates S-nitrosothiols and 3-nitrotyrosines, and activates the so called nitrosative signaling (Figure 1,panel A) (Heinrich et al., 2013). These events trigger downstream signals, maintaining normal synaptic plasticity and promoting neuronal differentiation or survival. For example, synaptic NO activates the cyclic adenosine monophosphate (cAMP) response element-binding protein (CREB) pathway, regulating long-term potentiation (LTP, an electrical correlate of learning and memory) and mediating neuronal survival (Lu et al., 1999).
By contrast, accumulating evidences suggest that hyperactivation of ‘extrasynaptic’ NMDARs can lead to an abnormal Ca2+influx into the postsynaptic neuron, with a subsequent overstimulation of nNOS, through the ternary NMDAR/ PSD95/nNOS complex formation. Thus, nNOS results permanently activated, and the overproduced NO is responsible for excessive protein nitrosation and nitrosylation, determiningnitrosative stress. Additionally, the excessive NO by reaction with superoxide anion (O2–), forms the highly reactive oxidant and cytotoxic molecule peroxynitrite (ONOO–), which triggers lipid peroxidation and damage (Heinrich et al., 2013). As a consequence of these events, neurotramission is impaired, due to mithocondrial dysfunction and synaptic damage (Figure 1,panelB). In this condition, nitric oxide activates the MAPK p38 and other downstream signaling proteins responsible for neuronal death, inducing apoptosis (Cao et al., 2005).
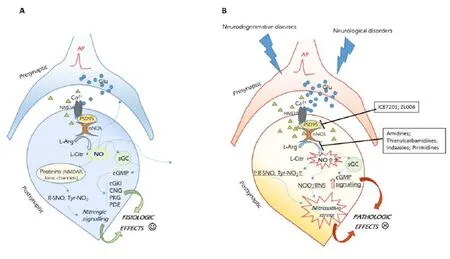
Figure 1 Schematic representation of NO synthetic and activity pathways in physiologic conditions (panel A), and pathologic conditions (panel B).
Therefore, blocking overstimulated neuronal nitric oxide synthase can be considered as a target to inhibit neurodegeneration and restore neurological functions. Actually, there are two distinct ways to inhibit NO overproduction from nNOS: 1) blocking the enzyme with inhibitors; 2) inhibiting the NMDAR-PSD95-nNOS complex (Figure 1, panel B).
Inhibitors of the neuronal NOS
The very first challenge in the discovery of nNOS inhibitors is to obtain selective molecules, which do not interfere with iNOS and eNOS. The difficulty in reaching this goal lies in the fact that the three isoforms have a very similar architecture, and it is necessary to exploit the subtle differences between the catalytic sites of the enzymes, to have selective ligands. Indeed, the early nNOS inhibitors, which were substrate analogues or arginine based dipeptides, were abandoned, since they showed unacceptable cardiovascular side effects (increase of blood pressure, heart failure,etc.), mainly due to their very low isoform selectivity. In the research of non-aminoacidic compounds able to selectively inhibit nNOS, sterically hindered amidines (1, 2,Table 1) gave interesting results (Maccallini et al., 2013). The phenyl and isopropyl groups of these molecules are the key fragments conferring them the desired selectivity, because they are responsible for the favorable interactions within the nNOS catalytic pocket. Better results have been obtained with thienylcarbamidines, which proved to be effective against human nNOS and showed neuroprotection in ischemia models and neurophatic pain (3, 4, Table 1) (Vaughan et al., 2010; Annedi et al., 2012a, b). Molecule4is currently in Phase III clinical trial for the treatment of acute migraine with aura. Recently, indazole based compounds have been reported as dual agents able to inhibit nNOS and to activate human carbonic anhydrase, another emerging target for the therapy of neurodegeneration. Compound5(Table 1) was the most interesting one of this series, showing high potency against nNOS (IC50= 0.174 μM), excellentin vitroselectivity with respect to eNOS (327 folds), together with a potent and selective activation of the human Carbonic Anhydrase isoform I (KA= 15 nM) (Maccallini et al., 2016). Another important chemical scaffold which generated potent and selective nNOS inhibitors was the pyrrolidinomethyl-2-aminopyridineone. Several molecules have been published endowed with very interesting biological profiles, but the lack of selectivity at tissue level, together with their poor bioavailability and physicochemical properties, limit theirin vivoapplication (Pigott et al., 2013). These unfavorable aspects have been overcome by the 2,4-disubstitued pyrimidine derivatives (6, Table 1), which are promising agents for the treatment of NO mediated neurological diseases (Annedi, 2015).
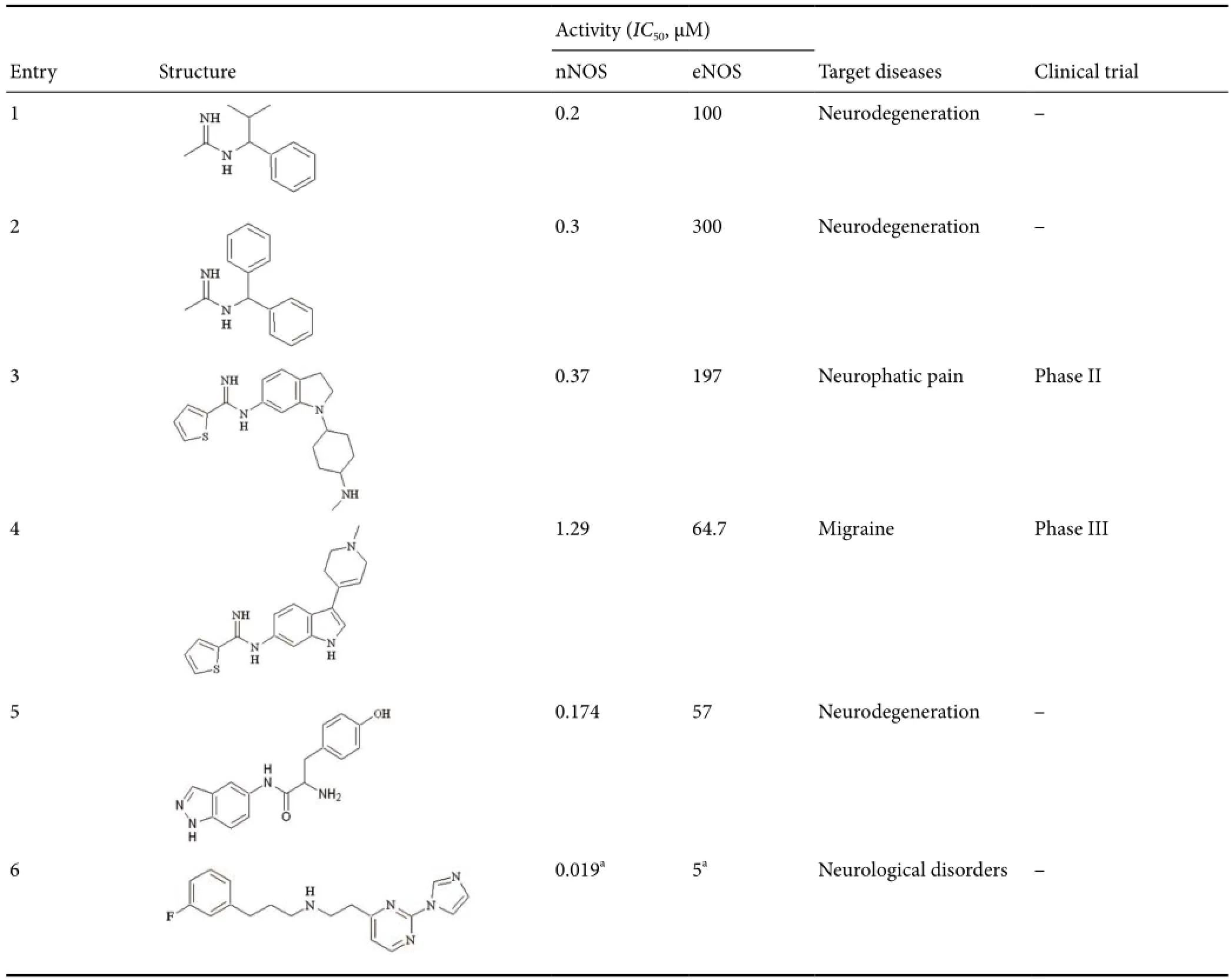
Table 1 Inhibitors of neuronal nitric oxide synthase.
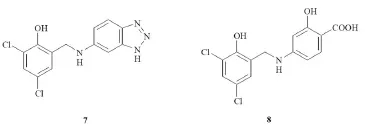
Figure 2 Small molecules able to inhibit neuronal nitric oxide synthase (nNOS) by dysrupting the PSD95–nNOS interaction.
Inhibitors of the NMDAR-PSD95-nNOS Complex
NMDARs excitotoxicity is mediated by nNOS, which is activated not only by the increased calcium concentration, but also by the binding with the PSD95 protein. Therefore, this protein interaction is a viable druggable target, providing an alternative and selective approach to the direct inhibition of nNOS or NMDAR.
From a medicinal chemistry viewpoint, this is a less investigated area and few compounds have been reported as PSD95/ nNOS inhibitors. NR2B9c, a small interference peptide that is supposed to disrupt with high potency both the interactions between the GluN2A subunit of the NMDAR and the PSD95, and between the latter and nNOS, is giving the most encouraging clinical results. It is currently under evaluation in patients with acute cerebral ischemia, in a phase III clinical trial (Aarts et al., 2002). Due to the poor pharmacokinetic profile of NR2B9c, investigations on different compounds able to actwith the same mechanism have been performed. IC87201 is a small molecule identified from a virtual screening of a 150,000-compound library, showing anIC50of 31 μM (7, Figure 2) (Loh et al., 2005). Notably, IC87201 disrupts the PSD95–nNOS interaction without affecting the interaction of PSD95 with other proteins. Based on the presumable structural requirements of the PSD95–nNOS interaction, it was synthetized ZL006 (8, Figure 2), another interesting small molecule endowed with high specificity and potency for disrupting the PSD95–nNOS interaction. ZL006 is neuroprotective against NMDAR-mediated excitotoxicityin vitroand ischemic neuronal injuryin vivo, and readily crosses the blood-brain barrier (Zhou et al., 2012).
Conclusions
In the last decade, the excessive production of nitric oxide by nNOS has been recognized as a major player in the induction and progression of neurological diseases. Medicinal chemists disclosed several molecules able to inhibit this enzyme, although there is still much to do to ameliorate the safety profile of these compounds, as well as their pharmacokinetics. Indeed, only few of the developed nNOS inhibitors have reached clinical trials. Future structure–activity relationship studies are also necessary to identify a pharmacophoric model to develop potent compounds targeting the NMDAR-PSD95-nNOS complex, as a promising strategy to inhibit the downstream death signaling factors responsible of neuronal death.
Author contributions:All authors wrote and revised the manuscript.
Conflicts of interest:None declared.
Aarts M, Liu Y, Liu L, Besshoh S, Arundine M, Gurd JW, Wang YT, Salter MW, Tymianski M (2002) Treatment of ischemic brain damage by perturbing NMDA receptor-PSD-95 protein interactions. Science 298:846-850.
Annedi SC, Ramnauth J, Maddaford SP, Renton P, Rakhit S, Mladenova G, Dove P, Silverman S, Andrews JS, Felice MD, Porreca F (2012) Discovery of cis-N-(1-(4-(methylamino)cyclohexyl) indolin-6-yl)thiophene-2-carboximidamide: a 1,6-disubstituted indoline derivative as a highly selective inhibitor of human neuronal nitric oxide synthase (nNOS) without any cardiovascular liabilities. J Med Chem 55:943-955.
Annedi SC, Maddaford SP, Ramnauth J, Renton P, Speed J, Rakhit S, Andrews JS, Porreca F (2012b) 3,5-Disubstituted indole derivatives as selective human neuronal nitric oxide synthase (nNOS) inhibitors. Bioorg Med Chem Lett 22:1980-1984.
Annedi SC (2015) Cell-permeable inhibitors of neuronal nitric oxide synthase open new prospects for the treatment of neurological disorders. J Med Chem 58:1064-1066.
Antonaci F, Ghiotto N, Wu S, Pucci E, Costa A (2016) Recent advances in migraine therapy. Springer Plus 5:637.
Cao J, Viholainen JI, Dart C, Warwick HK, Leyland ML, Courtney MJ (2005) The PSD95-nNOS interface: a target for inhibition of excitotoxic p38 stress-activated protein kinase activation and cell death. J Cell Biol 168:117-126.
Heinrich TA, da Silva RS, Miranda KM, Switzer CH, Wink DA, Fukuto JM (2013) Biological nitric oxide signalling: chemistry and terminology. Br J Pharmacol 169:1417-1429.
Loh JC, Yee-Wen LY (2005) Agents disrupting the interaction between postsynaptic density protein 95 and neuronal nitric oxide synthase for use as analgesics. WO 2005097090 A2 20051020.
Lu YF, Kandel ER, Hawkins RD (1999) Nitric oxide signaling contributes to late-phase LTP and CREB phosphorylation in the hippocampus. J Neurosci 19:10250-10261.
Maccallini C, Fantacuzzi M, Amoroso R (2013) Amidine-based bioactive compounds for the regulation of arginine metabolism. Mini-Rev Med Chem 13:1305-1310.
Maccallini C, Di Matteo M, Vullo D, Ammazzalorso A, Carradori S, De Filippis B, Fantacuzzi M, Giampietro L, Pandolfi A, Supuran CT, Amoroso R (2016) Indazole, pyrazole, and oxazole derivatives targeting nitric oxide synthases and carbonic anhydrases. ChemMedChem 11:1695-1699.
Pigott B, Bartus K, Garthwaite J (2013) On the selectivity of neuronal NOS inhibitors. Br J Pharmacol 168:1255-1265.
Vaughan D, Speed J, Medve R, Andrews JS (2010) Safety and pharmacokinetics of NXN-188 after single and multiple doses in five phase I, randomized, double-blind, parallel studies in healthy adult volunteers. Clin Ther 32:146-160.
Wang J, Jin L, Zhu Y, Zhou X, Yu R, Gao S (2016) Research progress in NOS1AP in neurological and psychiatric diseases. Brain Res Bull 125:99-105.
Yuste JE, Tarragon E, Campuzano CM, Ros-Bernal F (2015) Implications of glial nitric oxide in neurodegenerative diseases. Frontiers Cell Neurosci 9:332.
Zhou L, Li F, Xu HB, Luo CX, Wu HY, Zhu MM, Lu W, Ji X, Zhou QG, Zhu DY (2010) Treatment of cerebral ischemia by disrupting ischemia-induced interaction of nNOS with PSD-95. Nat Med 16:1439-1443.
*Correspondence to: Cristina Maccallini, Ph.D., cristina.maccallini@unich.it.
orcid: 0000-0003-2957-8650 (Cristina Maccallini)
10.4103/1673-5374.194707
Accepted: 2016-10-13
- 中国神经再生研究(英文版)的其它文章
- The status of Nrf2-based therapeutics: current perspectives and future prospects
- The Epigenome as a therapeutic target for Parkinson’s disease
- Six psychotropics for pre-symptomatic & early Alzheimer’s (MCI), Parkinson’s, and Huntington’s disease modification
- Applicability of tooth derived stem cells in neural regeneration
- Cortical spreading depression-induced preconditioning in the brain
- Neuroinflammation, neurodegeneration and regeneration in multiple sclerosis: intercorrelated manifestations of the immune response