
有氧运动对饮食诱导的肥胖鼠下丘脑和褐色脂肪BMP7表达的影响
2016-03-13 08:07龚文辉张素梅储珏
中国康复 2016年2期
龚文辉,张素梅,储珏
骨形成蛋白(bone morphogenetic proteins,BMPs)属于转化生长因子β蛋白超家族,共由14种蛋白组成,分别对于不同组织的生理功能发挥着迥异的调节作用[1-2]。BMP7促进脂肪细胞分化和产热功能,增加褐色脂肪的产热功能和总热量平衡[3-5]。褐色脂肪细胞能够利用脂质进行氧化代谢,增加能量的消耗,减少机体脂肪堆积[6-7]。研究显示,褐色脂肪组织能够增加啮齿动物能量消耗,减少肥胖,缓解饮食诱导的肥胖发生[8-9]。成年人颈-锁骨上部存在褐色脂肪库,部分人胸廓和脊柱旁肌也发现褐色脂肪,褐色脂肪产热作用能够对抗肥胖的发生[10-13]。本文拟建立饮食诱导的肥胖(diet induced obesity,DIO)鼠模型,观察下丘脑、肩胛褐色脂肪组织中BMP7表达,下丘脑产热功能调节酶腺苷酸(adenosine monophosphate,AMP)依赖的腺苷酸活化蛋白激酶(Amp-activated protein kinase,AMPK)表达,线粒体产热标志物环氧化酶2(Cycloxygenase 2,COX 2)表达,以及机体产热功能的变化情况,探索运动影响DIO大鼠产热功能的机制。
1 材料与方法
1.1 材料 雄性8周龄SD大鼠30只,体质量180~200g。
1.2 方法 30只大鼠随机分为基础组10只和DIO组20只,基础组予标准基础饲料喂养。①DIO模型制备。采用高脂肪饮食喂养6周制备饮食诱导DIO模型,高脂饲料由87.8%基础饲料,2%胆固醇,10%猪油,0.2%甲基硫氧嘧啶混合配置[14],DIO大鼠造模成功标准为体质量≥基础组体质量均值加1.4倍标准差[15]。②运动模型制备。DIO组高脂饲料喂养6周后,随机分为安静对照组(CO组)和有氧运动组(EO组),每组10只;基础组大鼠作为正常安静对照组(C组)。EO组运动处方:DIO大鼠进行8周强度约为70%~75%最大摄氧量的跑台训练[16],适应性训练3d后,跑台速度15 m/min[16],每次60 min,每日1次,每周训练5d。4只实验鼠未能完成运动干预,中途退出,最终纳入分析的各组实验鼠数量均为6只。基础组大鼠保持原生活方式不变。
1.3 指标检测 ①体质量、脂体比检测:第8周(实验第14周)训练后断头处死大鼠,心脏取血,取双侧下丘脑组织、肩胛部褐色脂肪、肾周脂肪、附睾脂肪,称重后液氮保存待检。脂体比=(附睾脂肪组织质量+肾周脂肪组织质量)/体质量。②静止代谢率(resting metabolic rate,RMR)、非颤抖性产热(nonshivering thermogenesis,NST)测定[17]: RMR测定以实验鼠单位时间内单位体重耗氧量表示,采用封闭式流体压力呼吸计检测;NST测定以代谢率结束后30min最大呼吸量为非颤抖性产热,检测前先予去甲肾上腺素溶液(0.2mg/ml)皮下注射,有效剂量与体质量相当。③褐色脂肪BMP 7、COX 2和下丘脑BMP 7、AMPK蛋白表达测定:免疫印记法检测蛋白表达。分别取褐色脂肪提取的100ugBMP 7和COX 2总蛋白,以及取下丘脑提取的100ugBMP 7和AMPK总蛋白,细胞裂解上清溶解物SDS-PAGE蛋白分离、转膜3h,分别加入1:2000相应抗体,洗膜,加1:5000辣根过氧化物酶-抗,曝光、洗片,图像分析系统定量分析。
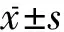
2 结果
2.1 体质量、脂体比变化 实验第6周,EO组、CO组大鼠体质量及脂体比均显著高于C组(P<0.01);实验第14周,EO组大鼠体质量及脂体比明显低于CO组(P<0.01)。见表1。

表1 3组大鼠实验第6及14周体质量及脂体比比较
与C组比较,aP<0.01;与CO组比较,bP<0.01
2.2 RMR、NST变化 实验第6周,EO组及 CO组大鼠RMR及NST均明显低于C组(P<0.01);实验第14周,EO组大鼠RMR及NST明显高于CO组及C组(P<0.01)。见表2。
表2 3组大鼠实验第6及14周RMR及NST变化比较 mlO2/(g.h) ,
与C组比较,aP<0.01;与CO组比较,bP<0.01
2.3 大鼠褐色脂肪BMP 7、COX 2蛋白表达变化 实验第6周, EO组及CO组大鼠褐色脂肪BMP 7及COX 2蛋白表达均明显低于C组(P<0.01);实验第14周,EO组及C组大鼠褐色脂肪BMP 7及COX 2蛋白表达明显高于CO组(P<0.01)。见表3。
表3 3组大鼠实验第6及14周褐色脂肪BMP 7及COX 2蛋白表达比较 ug,
与C组比较,aP<0.01;与CO组比较,bP<0.01
2.4 大鼠下丘脑BMP7、AMPK蛋白变化 实验第6周,EO组及CO组大鼠下丘脑BMP 7蛋白表达明显低于C组(P<0.01),AMPK蛋白表达明显高于C组(P<0.01);实验第14周,EO组及C组大鼠下丘脑BMP 7蛋白表达明显高于CO组(P<0.01),AMPK蛋白表达明显低于CO组(P<0.01)。见表4。
表4 3组大鼠实验第6及14周下丘脑BMP 7及COX 2蛋白表达比较 ug,
与C组比较,aP<0.01;与CO组比较,bP<0.01
3 讨论
为了探讨运动影响DIO大鼠能量代谢的机制,本研究进一步分析了中枢和外周产热标志物的变化情况。在下丘脑中,运动能够显著增加BMP 7蛋白表达,降低AMPK蛋白表达;褐色脂肪BMP 7蛋白表达,褐色脂肪线粒体功能蛋白COX 2蛋白表达也显著增加;总体产热功能提高,去甲肾上腺素刺激的RMR、NST均显著增加,体质量、脂体比减少。提示运动影响DIO大鼠能量代谢机制涉及中枢和外周的BMP7变化,分别在中枢通过改变交感神经系统的兴奋程度,在外周通过增加褐色脂肪的产热,调节机体总热量平衡,最终可能与全身脂肪分布、脂代谢改善及体质量下降存在一定联系。 研究显示,BMP 7是褐色脂肪中一种具有调节机体产热和能量平衡的功能蛋白,随着褐色脂肪的增加和减少,褐色脂肪中BMP 7的表达也出现了相同的变化趋势,证实BMP 7表达可以作为评价褐色脂肪功能变化的指标[3-4]。以往的研究均认为BMP家族在祖细胞特定的细胞分化过程中发挥调节作用[18]。 针对实验鼠的研究[19]显示:BMP 7 能够增加机体能量消耗,治疗饮食诱导的肥胖,降低体重。提示褐色脂肪功能变化与全身能量代谢平衡存在直接关系,并影响体质量的增减,为治疗肥胖和代谢综合症提供了一种新的选项。
本研究中DIO大鼠在第6周体质量、脂体比均比基础对照组显著增加。分析原因涉及到了中枢产热调节部位与外周产热组织功能的改变,证据来自本研究观察到的下丘脑和褐色脂肪BMP 7蛋白表达均显著下降;褐色脂肪线粒体功能减弱,大鼠去甲肾上腺素刺激的RMR、NST也显著下降。显示机体产热作用已经显著下降,从而导致脂肪在体内堆积,合理地解释了体质量、脂体比的增加机制。
研究显示,BMPs不仅仅增加外周褐色脂肪对肾上腺素刺激的反应,还在中枢激活交感神经对褐色脂肪的调节作用,在调节褐色脂肪产热作用的中枢部位丘脑下部的腹内侧核也发现了BMPs的表达[20-21]。长期以来丘脑下部的腹内侧核被认为是作为摄食中枢,调节摄食行为,新的研究又不断揭示了下丘脑在能量平衡调节中的重要作用。通过对下丘脑能量平衡调节发挥重要作用的AMPK研究,发现AMPK涉及到外周各种信号(如胃饥饿素)与下丘脑信号网络系统的整合,以利感知急性的血糖变化,以便增加机体葡萄糖应答反应的调节能力,发挥了一种“总能量计量器”的作用,既控制摄食,也调节产热作用[22- 23]。选择性的下调下丘脑AMPK,特别是丘脑下部腹内侧核内的AMPK,能够增加交感神经系统活性,上调褐色脂肪的产热能力(产热标志物),能够有效地控制诸如如肥胖、甲状腺功能亢进和其它能量代谢紊乱而引起的能量平衡失恒状态[20]。
研究发现,自愿转轮运动使雌性长爪沙鼠能量消耗水平提高, 运动开始后运动组RMR逐渐高于对照组,至第7周和第8周分别高于对照组32%和27%[24]。本研究中DIO大鼠给予8周有氧运动干预后,与安静对照组相比,去甲肾上腺素刺激的RMR、NST均显著增加,与Selman等[25-27]的研究结果一致,并观察到了体质量、脂体比的显著减少。运动调节能量摄入和热量支出的机制十分复杂,褐色脂肪的产热功能变化研究也有助于解释运动影响能量代谢的机制。褐色脂肪产热能力的增强常伴随着线粒体蛋白和线粒体功能基因COX 2活性的增加[28]。在正常人体急性运动实验中也发现了类似的结果,Tete[29]等给予16名研究对象(男、女各8名)4h 和24h的抗阻训练后COX 2 mRNA表达显著增加。作为褐色细胞产热参数,COX 2可以作为评价机体能量代谢的重要指标,与解偶联蛋白、鸟苷二磷酸、甲状腺素5,脱碘酶等能够间接反映褐色脂肪整体功能状态[29]。
本研究发现,DIO大鼠产热能力显著下降,体质量和脂体比增加,机制与下丘脑、褐色脂肪BMP 7以及褐色脂肪COX 2蛋白表达减少有关。8周有氧运动能够增加DIO大鼠产热能力,降低体质量和脂体比,机制与增加下丘脑、褐色脂肪BMP 7以及褐色脂肪COX 2蛋白表达有关。
[1] Reddi AH. Role of morphogenetic proteins in skeletal tissue engineering and regeneration[J]. Nat Biotechnol,1998,16(3): 247-252.
[2] Miyazono K. Signal transduction by bone morphogenetic protein receptors: functional roles of Smad proteins[J]. Bone,1999,25(1): 91-93.
[3] Schleinitz D, Kloting N, Bottcher Y, et al. Genetic and evolutionary analyses of the human bone morphogenetic protein receptor 2 (BMPR2) in the pathophysiology of obesity[J]. PLoS One, 2011,6(2): 155-161.
[4] Tseng YH, Kokkotou E, Schulz TJ, et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure[J]. Nature,2008, 454(7207): 1000-1004.
[5] Wang RN, Green J, Wang Z, et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases[J]. Genes Dis,2014,1(1): 87-105.
[6] Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance[J]. Physiol Rev, 2004, 84(1): 277-289.
[7] Porter C, Borsheim E, Sidossis LS. Does adipose tissue thermogenesis play a role in metabolic health[J]. J Obes, 2013,23(2): 204-214.
[8] Ghorbani M, Claus TH, Himms-Hagen J. Hypertrophy of brown adipocytes in brown and white adipose tissues and reversal of diet-induced obesity in rats treated with a beta3-adrenoceptor agonist[J]. Biochem Pharmacol,1997,54(1): 121-131.
[9] Bachman ES, Dhillon H, Zhang CY, et al. betaAR signaling required for diet-induced thermogenesis and obesity resistance[J]. Science,2002,297(5582): 843-845.
[10] Cypess AM, Lehman S, Williams G, et al. Identification and importance of brown adipose tissue in adult humans[J]. N Engl J Med,2009,360(15): 1509-1517.
[11] Saito M, Ogura Y, Matsushita M, et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity[J]. Diabetes,2009,58(7): 1526-1531.
[12] van MLWD, Vanhommerig JW, Smulders NM, et al. Cold-activated brown adipose tissue in healthy men[J]. N Engl J Med,2009,360(15): 1500-1508.
[13] Chechi K, Nedergaard J, Richard D. Brown adipose tissue as an anti-obesity tissue in humans[J]. Obes Rev,2014, 15(2): 92-106.
[14] 何晓烨. 高脂饮食在诱导大鼠肥胖及肥胖抵抗中的作用[J]. 中国临床康复, 2002,19(1): 60-61.
[15] 徐叔云, 陈修.药理实验方法学[M].第2版. 北京:人民卫生出版社,1992, 19-20.
[16] Thomas DP, Zimmerman SD, Hansen TR, et al. Collagen gene expression in rat left ventricle: interactive effect of age and exercise training[J]. J Appl Physiol,2000, 89(4): 1462-1468.
[17] 许婧. 黑腹绒鼠能量代谢对不同季节和温度的响应[D]. 浙江师范大学, 2007,34-35.
[18] Tseng YH, Kokkotou E, Schulz TJ, et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure[J]. Nature,2008,454(7207): 1000-1004.
[19] Townsend KL, Suzuki R, Huang TL, et al. Bone morphogenetic protein 7 (BMP7) reverses obesity and regulates appetite through a central mTOR pathway[J]. FASEB J, 2012,26(5): 2187-2196.
[20] Lopez M, Varela L, Vazquez MJ, et al. Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance[J]. Nat Med,2010,16(9): 1001-1008.
[21] Morton GJ, Cummings DE, Baskin DG, et al. Central nervous system control of food intake and body weight[J]. Nature,2006,443(7109): 289-295.
[22] Lage R, Vazquez MJ, Varela L, et al. Ghrelin effects on neuropeptides in the rat hypothalamus depend on fatty acid metabolism actions on BSX but not on gender[J]. FASEB J,2010,24(8): 2670-2679.
[23] McCrimmon RJ, Shaw M, Fan X, et al. Key role for AMP-activated protein kinase in the ventromedial hypothalamus in regulating counterregulatory hormone responses to acute hypoglycemia[J]. Diabetes,2008,57(2): 444-450.
[24] 胡振东, 王德华. 自愿转轮运动条件下雌性长爪沙鼠的体重、能量代谢和血清瘦素含量的变化[J]. 中国运动医学杂志,2007, 15(5): 605-608.
[25] Selman C, McLaren JS, Collins AR, et al. Antioxidant enzyme activities, lipid peroxidation, and DNA oxidative damage: the effects of short-term voluntary wheel running[J]. Arch Biochem Biophys,2002,401(2): 255-261.
[26] Levin BE, Meynell AA. Chronic exercise lowers the defended body weight gain and adiposity in diet-induced obese rats[J]. Am J Physiol Regul Integr Comp Physiol,004, 28(4): 771-778.
[27] Gattermann R, Weinandy R, Fritzsche P. Running-wheel activity and body composition in golden hamsters [J]. Physiol Behav,2004,82(2-3): 541-544.
[28] 张志强, 刘全生, 李纪元. 长爪沙鼠褐色脂肪组织和肝脏产热特征的季节性变化[J]. 动物学报,2006,(6): 35-37.
[29] Tate TH, Hyek MF. SERCA2a and mitochondrial cytochrome oxidase expression are increased in hearts of exercise[J]. AJP - Heart, 1996,271(1): H68-H72.
猜你喜欢
航天器环境工程(2022年6期)2023-01-03
森林工程(2020年6期)2020-12-14
北方人(2020年22期)2020-12-02
疯狂英语·新悦读(2020年4期)2020-06-18
江苏农业科学(2020年8期)2020-06-01
阅读(快乐英语中年级)(2019年11期)2019-09-10
中老年健康(2017年9期)2017-12-13
校园英语·下旬(2016年4期)2016-05-09
安徽医科大学学报(2015年9期)2015-12-16
中学科技(2014年11期)2014-12-25
