
不同生态环境中梭梭枝系构型特征分析
2016-06-14 15:46史红娟于秀立庄丽
江苏农业科学 2016年4期
史红娟+于秀立+庄丽
摘要:按降水差异在古尔班通古特沙漠南缘自西向东依次选择克拉玛依、121团、150团、阜康、奇台5个样点,研究不同生态环境中梭梭的构型特征,以求探讨荒漠植物梭梭对环境的适从机理。结果显示:分枝率数值依样地顺序大体呈减小的趋势,阜康和奇台的分枝总数和1级分枝数远远大于其他3个样地。枝径比和分枝率有着相似的发展趋势;各级分枝枝长大体上依所选样地顺序减短;各样地1~4级分枝角度自36°~ 62°逐渐增大,1~3级分枝角度增大幅度不明显,4级分枝角度增幅在10°以上,各样地分枝角度大体依样地顺序减小;克拉玛依、121团和150团的梭梭外部整体轮廓大致呈椭球形,阜康和奇台梭梭外部整体轮廓趋于球形。据野外观察,发现克拉玛依、121团以及150团的梭梭枝系构型大致呈伸展型,阜康和奇台的梭梭枝系构型大致呈发散型。
关键词:生态环境;沙漠植物;梭梭;枝系构型特征
中图分类号: S718.42
文献标志码: A
文章编号:1002-1302(2016)04-0217-04
植物由根、茎、叶、芽和果实组成,因此植物构型是由地上部分枝系构型和地下部分根系构型组成的,表征植株不同构件在空间的不同生长及分配方式[1-3]。植株的各个构件共同组成植物构型[4-5]。
植株生长较为缓慢,受周围环境影响比较大,因此,植物在不同生态环境中存在着构型分异[6]。而这种分异是植物对外界生存环境一种选择适从,也是种内构型差异的原因[7]。研究表明,引起这种差异的原因有植物自身遗传因素[8]和环境因素,而且影响荒漠植物构型最主要的环境因子为水分、温湿度、土壤理化性质[9-10],反映到植物构型指标具体表现为分枝率、分枝长度、分枝角度和枝径比等指标数值的差异。但是,大量的证据表明,这4个构型指标不能单一表征植物构型特征,需协同参考来衡量植物的构形特征[11-13]。
1 材料与方法
1.1 研究区概况
试验依据降水差异在古尔班通古特沙漠南缘自西向东依次选择克拉玛依、石河子(121团、150团)、阜康、奇台5个样点。古尔班通古特沙漠位于准葛尔盆地的中央地带。地理坐标为44°11′~46°20′N,84°31′~90°00′E,这里夏季燥热干旱,冬季酷寒漫长,是典型的温带大陆性干旱气候。年日温差比较大,植物种类大为耐盐碱和抗干旱的荒漠植物种。除大片建群种梭梭(荒漠植物梭梭在古尔班通古特沙漠的分布达100万hm2)、白梭梭外,还有柽柳、白皮沙拐枣、苦艾蒿、白蒿、琵琶柴、碱蓬、角果藜、猪毛菜、囊果苔草和一些短命植物。研究区生态环境概况汇总见表1。
1.2 试验方法
1.2.1 标准珠选择及构型指标测定方法 在每个样地拉100 m×100 m样方,在样方内选择基径>6.5 cm(成熟期)[14]标准珠进行取样测量。标准珠的选择标准为:(1)植株生命力旺盛,无人为破坏,无动物及病虫迫害;(2)生长环境良好;(3)种内种间保持间距,防止资源竞争对植株构型影响。最终共选择了标准珠107棵。
枝序的确定有很多种方法,本试验选择向心式法[15]测量各个地上部分构型指标。向心式法选择最外层第1小枝作为1级分枝,2个1级分枝相遇定义为2级分枝,以此类推,2个相同级别分枝相遇,取下级分枝作为枝级。
用精度0.1 cm的卷尺测取标准珠的株高、冠幅和分枝长度,用游标卡尺测取植株基径和各级分的枝径,植株各级分枝角度用量角器测量。
1.2.2 构型指标计算方法
1.2.3 环境因子测定 环境因子分别在克拉玛依、121团、150团、阜康、奇台5个样地测定,温度、湿度测定利用干湿度计。土壤在标准珠中选3棵树下取样,各取3个0~60 cm土壤样品,作为测定土壤水分、pH的样本。土壤含水量采用烘干(105 ℃)法测定,用ZD-2型电位滴定计测定土壤pH值。
1.3 数据分析
试验所得数据用Excel进行整理和作图,梭梭枝系构型临摹图根据野外观察及数码照片临摹而来,再采用Photoshop CC转化处理成JPG格式。
2 结果与分析
植物枝系构型由植株的分枝率、各级分枝长度、各级分枝角度和枝径比等构型参数共同影响决定。比如:分枝率反映的是植株的分枝能力,分枝长度表征植株的伸展能力,分枝角度代表枝条的承载能力强弱,枝径比能够说明上级枝条对下级枝条的承载力度[16]
2.1 不同生态环境中梭梭分枝率特征
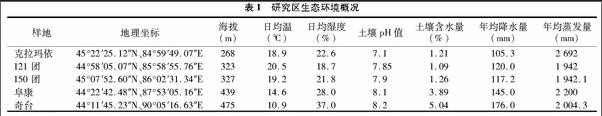
如图1所示,研究区梭梭分枝率数值OBR和SBR自121团至奇台皆依次减小。OBR及SBR2 ∶1数值远远小于 SBR3 ∶2 和SBR4 ∶3;研究区分枝总数和第一级分枝数自克拉玛依至奇台呈增长趋势,最低值与最高值相差悬殊,平均相差112个总分枝数和103个1级平均分枝数。
OBR数值越小,植株当年生植株越多,即第1级分枝数会越多。SBR数值越小,说明植株该级枝级枝条分枝能力越强。很明显,奇台研究区的梭梭拥有更多的分枝总数和同化枝数量。这与奇台及其他样地的环境条件是密不可分的,奇台的降水条件是所选样地中最充足的,同化枝由足够的水分支持,因此滋生较多枝条。
2.2 不同生态环境中梭梭分枝长度特征
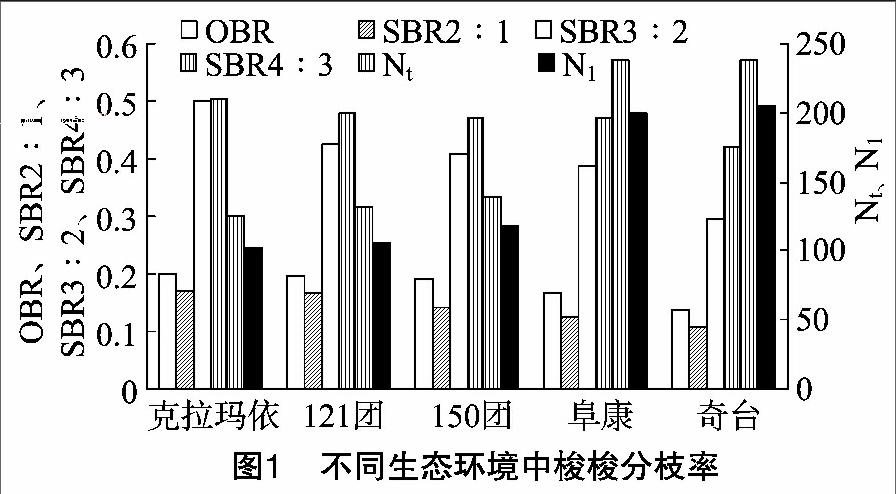
如图2所示,各样地各级分枝长度自121团至奇台大致呈减小的趋势,而且对于各级最长和最短分枝,1级分枝长度平均相差14.41 cm,2级分枝长度平均相差10 cm,3级分枝长度平均相差29.59 cm,4级分枝长度相差42.44 cm。
分枝长度大,说明植株自身伸展能力较强,也说明由于环境条件的限制,植株需要强的伸展能力来获取空间资源,以满足自身生长需求。
2.3 不同生态环境中梭梭分枝角度特征
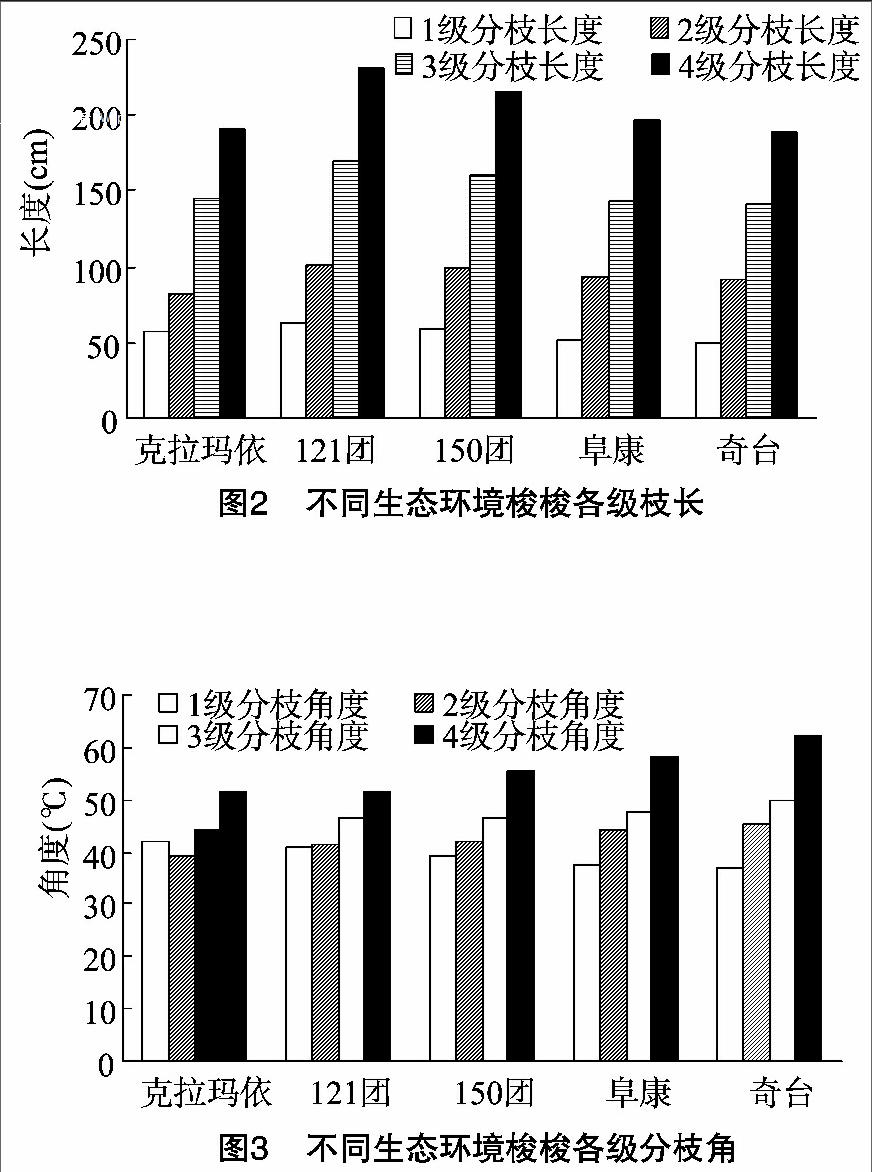
图3显示各个样地1级分枝角度自克拉玛依至奇台呈减小趋势,3~4级分枝角度依样地选择次序逐渐增大;1级分枝角度平均39°,4级分枝角度平均56°,1级至3级各个样地分枝角度数变动不是很明显,但是由3级至4级,度数增加10°以上,幅度较大。
分枝角度大,说明老枝对新枝的承载能力强,4级分枝角度最大,因为要承受1~3级分枝的生物量,奇台1级分枝角度最小,这是由其较多的1级分枝数决定的,分枝数越多,同化枝对空间的竞争越大,单位枝条获取的生存空间越小,反映到分枝角度则数值较小,并且由短的分枝长度来平衡生长竞争。
2.4 不同生态环境中梭梭枝径比特征
比较图1和图4,发现枝径比和分枝率有着相似的数值增减趋势,枝径比表征的是老枝对新枝的承载能力。数值越大,表明老枝对新枝的承载能力越强。图中显示,各级逐步分枝率自克拉玛依至奇台数值是减小的,RBD1 ∶2最大值与最小值差异明显;RBD2 ∶3和RBD3 ∶4数值差异不明显。
2.5 不同生态环境中梭梭整体枝系构型特征
2.5.1 不同生态环境中梭梭枝系构型图 图5至图9是根据实地观察各样地的梭梭枝系生长构型及对标准株数码照的临摹而得到的图片,据观察,克拉玛依、121和150团的梭梭趋于伸展型生长,最高级分枝角度大,分枝少,同化枝枝条也较少,但是枝条长度要比阜康和奇台的梭梭枝条长度长,并且枝节较长;而阜康和奇台的梭梭趋于发散型生长,最高级分枝角较大,1级分枝角度由于同化枝枝条繁多而呈现较小的数值,同化枝虽多,但是枝条长度小,枝节也较短。这是由各样地的自然生态环境决定的,相比较而言,由克拉玛依越往东至奇台,降水越充足,土壤条件越适宜梭梭生长,野外观察到奇台的梭梭枝条相较其他样地明显更为嫩绿和饱满,这与数据分析结果是吻合的。
2.5.2 不同生态环境中梭梭冠高比特征 从表2可以看出,所选各个样地的标准株平均基经大小较为接近,植株高度最大值在121团,植株冠幅最大值在150团。相比较而言,阜康和奇台的梭梭冠幅和高度数值较为接近,呈现出近球形的外部轮廓,而克拉玛依、121团和150团的梭梭则呈现出椭球形的外部轮廓。这与梭梭自身对环境选择适从密切相关。
3 结论与讨论
植物构型是植物自身遗传因子和环境相互作用的最终外观表达。植物个体是有生命的个体,有生命就代表其会为了自身生存而做出最有利于其发展的生长策略,环境与植物构型之间息息相关。
试验所选5个样地,奇台的总体分枝率OBR数值最小,逐步分枝率SBR数值大体依所选样地顺序依次减小,阜康和奇台的分枝总数远远大于其他3个样地。枝径比和分枝率有着正比的发展趋势。各级分枝枝长也是依所选样地顺序减小。各样地1级分枝角度依所选样地顺序逐渐减小,3~4级分枝角度则逐渐增大。各样地1~4级分枝角度自36°~ 62°逐渐增大。1~3级分枝角度增减幅度不明显,4级分枝角度增幅在10°以上。
这与各个样地梭梭的生长策略是相关的,121团和150团的环境条件恶劣,光照水分等条件都次于阜康和奇台,所以新生枝条较少,表现为较大的总体分枝率OBR数值。枝条少,则分摊到每一根枝条的营养物质相对较多,枝条对空间资源的竞争相对较弱,因此会加长生长。而奇台样地的梭梭生存环境相对要好一些,降水明显比其他几个样地充足,土壤含水量也明显较大,所以植株会萌发多的枝条,而且下级分枝比上级分枝明显多出很多,会表现出较小的逐步分枝率SBR值。同化枝枝条繁多会导致同化枝对空间资源的竞争力度加大,单位枝条所获得的空间也会减小,表现出较小的1级分枝角度和较短的同化枝枝长。
通过冠高比数值,发现克拉玛依和阜康的梭梭冠幅大于其生长高度,121、150和奇台梭梭冠幅小于其生长高度;相比较,阜康和奇台的梭梭冠幅和高度数值比较接近,外部轮廓趋于球形外部轮廓,克拉玛依、121团和150团的梭梭或者冠幅大于株高,或株高大于冠幅,外部轮廓呈椭球形。据野外观察及数据分析,发现克拉玛依、121团以及150团的梭梭枝系构型大致呈伸展型,枝条长度长,但是下级枝条数量少;阜康和奇台的梭梭枝系构型大致呈发散型,枝条长度相对较短,但是下级枝条数量多。各样地的梭梭通过各自的构型变异来适应外部生态环境。
参考文献:
[1] Harper J L Population biology of plant[M]. London:Academic Press,1977.
[2]White J. The plant as a met a population [J]. Ann R Ecol Syst,1979,10:109-145.
[3]屈志强,刘连友,吕艳丽. 沙生植物构型及其与抗风蚀能力关系研究综述[J]. 生态学杂志,2011,30(2):357-362.
[4]Harper J L Plant demography and ecological theory[J]. Oikos,1980,35:244-253
[5]Halle F,Oldeman R,Tomlinson P B,et al. Tropical trees and forests,an architecture analysis[M]. Heidelberg:Springer-Verlag,1978.
[6]张志浩,杨晓东,孙宝伟,等. 浙江天童太白山不同群落植物构型比较[J]. 生态学报,2015,35(3):761-769.
[7]费世民,蔡小虎,何亚平,等. 生物柴油植物麻疯树群体构型对资源性环境因素的反应[J]. 四川林业科技,2009,30(5):1-8.
[8]Sussex L M. The evolution of plant architecture[J]. Current Opinion in Plant Biology,2001,4(1):33-37.
[9]王泽.阿不都克玉木·米吉提,吐尔逊娜依·热依木,等. 北疆荒漠区梭梭水分,盐分和养分状况研究[J]. 新疆农业大学学报,2014,37(4):339-344.
[10]晋 瑜,颜 安,潘存德,等. 准噶尔盆地西北边缘荒漠植物群落分布的主要环境因子影响分析[J]. 内蒙古农业大学学报:自然科学版,2013,34(5):42-46.
[11]何明珠,张景光,王 辉. 荒漠植物枝系构型影响因素分析[J]. 中国沙漠,2006,26(4):625-630.
[12]贾 程,何 飞,樊 华,等. 植物种群构件研究进展及其展望[J]. 四川林业科技,2010,31(3):43-50.
[13]孙栋元,赵成义,王丽娟,等. 荒漠植物构型研究进展[J]. 水土保持研究,2011,18(5):281-287.
[14]贾志清,卢 琦. 梭梭[M]. 北京:中国环境科学出版社,2005:1-14.
[15]宋于洋. 梭梭构件格局的环境变异[J]. 西北林学院学报,2008,23(6):60-65.
[16]王丽娟,孙栋元,赵成义,等. 准噶尔盆地梭梭、白梭梭植物构型特征[J]. 生态学报,2011,31(17):4952-4960.徐 华,刘婉华,姚萍萍,等. 柑橘潜叶蛾危害对脐橙叶片几种生理指标的影响[J]. 江苏农业科学,2016,44(4):221-223.
猜你喜欢
哈哈画报(2022年4期)2022-04-19
科学大众(2020年17期)2020-10-27
商业会计(2016年15期)2016-10-21
商业会计(2016年13期)2016-10-20
科学与财富(2016年28期)2016-10-14
绿色中国(2015年6期)2015-07-03
