
肿瘤的氨基酸代谢变化及其意义
2016-07-10 10:27苏兰鸿孙林冲高平
中国生化药物杂志 2016年9期
苏兰鸿,孙林冲,高平
(中国科学技术大学 生命学院,安徽 合肥 230027)
肿瘤的氨基酸代谢变化及其意义
苏兰鸿,孙林冲,高平Δ
(中国科学技术大学 生命学院,安徽 合肥 230027)
近年来,肿瘤代谢重编程不断被更新,其中氨基酸代谢重编程越来越受到关注,尤其是谷氨酰胺代谢。谷氨酰胺是人体内最为丰富的非必需氨基酸之一,但对肿瘤而言,却是特定情况下的必需氨基酸。因此,通过靶向谷氨酰胺代谢有望发展新的抗癌药物。此外,丝氨酸/甘氨酸代谢对肿瘤细胞的作用也在不断地被阐明。本文旨在讨论肿瘤细胞的谷氨酰胺代谢重编程及其在肿瘤发生发展过程中发挥的作用,并简单介绍丝氨酸/甘氨酸代谢对肿瘤的影响。
肿瘤;代谢;谷氨酰胺
随着现代医学的发展,人类已经可以战胜许多病痛,然而肿瘤却始终威胁着人类的健康。自上个世纪开始,科学界就对肿瘤发生发展的机制展开了广泛的研究。2011年,Hanahan和Weinberg总结的肿瘤细胞新十大特征中新增肿瘤细胞能量代谢重编程[1],表明细胞能量代谢重编程已然成为肿瘤研究的新热点。其中广为人知的是“Warburg effect”概念的提出,即上个世纪20年代德国生化学家Warburg发现在常氧条件下,肿瘤细胞仍然通过糖酵解途径来获取能源物质以满足快速增殖的一种现象[2-3]。虽然糖代谢的重编程在肿瘤中起到重要的生物学意义,但某些肿瘤细胞却存在不依赖葡萄糖的现象,后续研究表明谷氨酰胺在其中发挥着重要作用。
直到1955年,谷氨酰胺对细胞增殖的重要性才被Eagle首次强调。谷氨酰胺不仅是人体内最丰富的非必需氨基酸,还是肿瘤细胞培养基中必不可少的关键成分[4]。在肿瘤细胞中,谷氨酰胺主要被谷氨酰胺酶(Glutaminase,GLS)催化转变为谷氨酸,进一步又被谷氨酸脱氢酶(Glutamate dehydrogenase,GLUD)催化形成α-酮戊二酸,最后进入TCA循环,实现对TCA循环的回补作用。其中,谷氨酰胺来源的苹果酸可以穿梭出线粒体,进入细胞质基质,在苹果酸酶1(malic enzyme 1,ME1)的催化下转变为丙酮酸,然后被乳酸脱氢酶(lactate dehydrogenase A,LDHA)催化产生乳酸并排至胞外,这一代谢过程我们称之为“Glutaminolysis”。另外,谷氨酰胺来源的柠檬酸也能穿梭出线粒体进入细胞质基质,参与脂类物质的从头合成。除了给生物大分子提供碳骨架,谷氨酰胺还是重要的氮源,是合成核苷酸和其他非必需氨基酸的基础。
因此本文将对肿瘤细胞的谷氨酰胺代谢变化及其生物学意义进行重点介绍。另外,除谷氨酰胺外,还有一些氨基酸对肿瘤的作用也越来越受到关注,鉴于此,本文还将简单介绍肿瘤细胞中的丝氨酸/甘氨酸代谢。
1 谷氨酰胺代谢在肿瘤中的变化
对于正常细胞而言,谷氨酰胺是非必需氨基酸,可以通过谷氨酰胺合酶合成满足生长所需。但肿瘤细胞通过自身合成谷氨酰胺无法满足快速增殖的要求,往往需要从外界摄入大量的谷氨酰胺,意味着谷氨酰胺是肿瘤细胞的“必需氨基酸”。肿瘤细胞则主要通过增加膜上的转运体从胞外摄入谷氨酰胺或增强谷氨酰胺代谢通路中关键代谢酶的表达与活性,来满足自身生存与增殖所需的营养物质和能量。
1.1 谷氨酰胺转运体 谷氨酰胺是亲水性的氨基酸,无法直接透过细胞膜入胞,需要转运体协助,故转运体是决定谷氨酰胺能否被肿瘤细胞利用的第一道门槛。至今,已有不少研究表明氨基酸相关转运体在肿瘤细胞中存在表达上调的现象,包括与谷氨酰胺代谢密切相关的转运体SLC1A5(ASCT2)、SLC6A14、SLC7A5(LAT1)等[5],其中SLC1A5通过把胞内Na+转运出胞,将谷氨酰胺转运入胞;SLC6A14将谷氨酰胺以及Na+和Cl-共同转运入胞;而SLC7A5负责将谷氨酰胺转运出胞的同时把亮氨酸等必需氨基酸转运入胞。
作为转运谷氨酰胺入胞的关键转运体,SLC1A5和SLC6A14在肿瘤细胞中发挥重要作用,其中 SLC6A14还能为SLC1A5和其他转运体提供物质基础(如Na+和谷氨酰胺等),决定他们功能的发挥。已有报道,阻断SLC6A14会影响肿瘤细胞内蛋白质、脂类、核苷酸的合成等,最后导致细胞死亡[6]。SLC1A5也会影响其他转运体,尤其是SLC7A5。Nicklin等[7]发现SLC1A5缺失使得胞内谷氨酰胺含量降低,导致SLC7A5无法正常转运亮氨酸入胞,mTOR信号通路不能被激活,最后影响肿瘤细胞的生长。
1.2 谷氨酰胺酶 谷氨酰胺酶(GLS)有2种亚型,分别是肾型谷氨酰胺酶(GLS1)和肝型谷氨酰胺酶(GLS2)[8]。迄今为止,已有不少研究表明肿瘤细胞中存在GLS1明显上调的现象。近些年有不少对谷氨酰胺代谢中谷氨酰胺酶上调具体机制的研究。Gao等[9]发现在B淋巴瘤细胞和前列腺癌细胞中,高表达的癌基因c-Myc能够通过抑制miR-23a/b来上调GLS1,增强谷氨酰胺代谢,促进肿瘤细胞的增殖。在此基础上,Wang 等[10]在乳腺癌和B淋巴瘤细胞中发现GLS1的上调依赖于Rho GTPases和NF-κB信号通路的作用,并证明通过敲低GLS1能够有效地抑制肿瘤细胞的增殖。
但是,GLS2对肿瘤细胞发挥的作用存在矛盾性。Suzuki 等[11]在结肠癌细胞中发现,GLS2是抑癌基因P53的靶基因,能够抑制肿瘤。在某些肝癌细胞中发现GLS2的表达是降低甚至缺失的,过表达GLS2会减少肝癌细胞的克隆形成[12]。但在MYCN扩增的神经母细胞瘤中观察到,GLS2活性的丧失使肿瘤细胞的谷氨酰胺代谢被抑制,进而导致细胞增殖减慢[13]。
2 增强谷氨酰胺代谢对肿瘤细胞的生物学意义
在肿瘤细胞代谢重编程中,葡萄糖通过糖酵解产生许多中间代谢产物,进入其他代谢旁路,如磷酸戊糖途径、己糖合成通路、丝氨酸/甘氨酸合成通路等,合成增殖所需的生物大分子。但是,葡萄糖来源的丙酮酸仅少部分进入TCA循环,绝大多数通过乳酸脱氢酶(LDHA)以乳酸形式排出。在这种情况下,谷氨酰胺可以很好地回补TCA循环,用于合成其他氨基酸和脂类物质等,甚至通过苹果酸或草酰乙酸重新进入糖酵解途径,维持在葡萄糖缺乏情况下相关代谢通路的进行。更重要的是,谷氨酰胺提供葡萄糖所不具备的氮源,是合成嘌呤环和嘧啶环的关键,也是合成许多非必需氨基酸的基础。另外,谷氨酰胺代谢还可以参与细胞信号转导。
2.1 为合成生物大分子提供碳骨架 谷氨酰胺是除葡萄糖外另一重要的能源物质,能通过TCA循环过程中的中间代谢物逆向进入糖酵解途径,给葡萄糖缺乏或葡萄糖不依赖的肿瘤细胞补充物质与能量。在葡萄糖饥饿的情况下,Vincent等[14]发现谷氨酰胺来源的草酰乙酸能被线粒体型磷酸烯醇式丙酮酸羧化酶(PCK2)催化变为磷酸烯醇式丙酮酸,逆向进入糖酵解,参与磷酸戊糖途径,或丝氨酸合成途径等。
谷氨酰胺还是肿瘤细胞脂类物质合成的重要物质基础。在低氧条件下,低氧诱导因子HIF能够通过激活丙酮酸脱氢酶激酶(PDK)来抑制丙酮酸脱氢酶(PDH)的活性,使得葡萄糖来源的丙酮酸几乎不进入TCA循环,从而不能为脂类的从头合成提供葡萄糖来源的乙酰辅酶A。这时,谷氨酰胺起到非常关键的作用。Metallo等[15]观察到人非小细胞肺癌细胞可以通过还原性谷氨酰胺代谢合成脂类物质,其中细胞质基质中的异柠檬酸脱氢酶1(IDH1)发挥着重要作用,其能将α-酮戊二酸催化成异柠檬酸,最后以乙酰辅酶A的形式参与脂类从头合成。与此同时,在线粒体缺陷的骨肉瘤细胞中,Mullen等[16]发现谷氨酰胺也是通过还原路径合成柠檬酸,进入脂类从头合成路径。
2.2 为合成核苷酸和非必需氨基酸提供氮源 2016年,Pavlova和Thompson总结了肿瘤代谢六大特征,指出肿瘤细胞需要许多氮源用于合成核苷酸和非必需氨基酸,而作为丰富的非必需氨基酸,谷氨酰胺能为肿瘤细胞提供氮源[17]。谷氨酰胺可以分别通过三步独立的酶促反应合成嘌呤和两步反应合成嘧啶,将氨基(γ氮)贡献出来,自己则转变为谷氨酸。Gaglio等[18]发现在谷氨酰胺缺乏的情况下,细胞周期阻滞在G1/S期,但是回补核苷酸能减缓这种阻滞作用,从而促进肿瘤细胞生长。
作为谷氨酰胺代谢的中间代谢物,谷氨酸是合成非必需氨基酸的主要氮源供体,能将谷氨酰胺来源的氨基(α氮)贡献给非必需氨基酸,同时产生的α-酮戊二酸可以进入TCA循环为氨基酸提供碳骨架。例如,在天冬氨酸氨基转移酶的催化下,草酰乙酸接受谷氨酸提供的氨基,转变为天冬氨酸。而天冬氨酸又能为合成甲硫氨酸、苏氨酸和赖氨酸等氨基酸提供重要的前体物质。总而言之,这些氨基酸的氨基源自谷氨酰胺。
2.3 为维持胞内氧化还原平衡提供还原力 肿瘤细胞内普遍存在活性氧(reactive oxygen species,ROS)升高的现象,主要是由代谢活动加快、线粒体功能障碍、细胞受体信号增强、癌基因、氧化酶和环加氧酶活性增强等原因引起的[19]。低水平的ROS通过可逆地氧化蛋白巯基,改变蛋白质结构和功能参与到细胞信号通路的转导;也可通过引起DNA的损伤和突变,导致基因组的不稳定,最后使细胞发生癌变[20]。但同时细胞必须抵抗过多的ROS造成的毒害作用。
谷氨酰胺代谢能通过2种方式来抵抗过多的ROS,维持胞内氧化还原平衡。首先,谷胱甘肽是细胞内主要的抗氧化剂,由谷氨酰胺提供碳源和氮源。谷氨酰胺饥饿或敲低谷氨酰胺酶(GLS)会抑制GSH的产生,导致胞内ROS水平增加[9]。再者,谷氨酰胺来源的苹果酸被苹果酸酶(ME)催化可产生NADPH,为细胞提供还原力,维持谷胱甘肽的还原状态,还可为脂类合成提供还原力。
2.4 参与细胞信号转导调节 谷氨酰胺除了提供能源物质外,还参与细胞信号转导。近些年,不少研究揭示谷氨酰胺在激活必需氨基酸依赖的mTOR信号通路中发挥重要作用。此外,谷氨酰胺代谢的中间产物可通过自分泌或旁分泌方式参与信号转导,最终影响肿瘤的生长。谷氨酸是中枢神经系统中已知的神经递质,能通过离子型或代谢型的受体来传递信号。其中,代谢型受体mGluRs是G蛋白偶联受体家族的一员,在肿瘤发展过程中发挥重要作用[21]。mGluRs的活化能激活ERK(extracellular signal-regulated protein kinase)信号通路和PI3K(phosphatidy-linositol 3’-kinase)信号通路,促进细胞增殖。Pollock等[22]在黑色素瘤细胞膜上观察到mGluR1的表达,但在正常的黑色素细胞膜上并未检测到;在小鼠的黑色素细胞中人为过表达mGluR1,发现黑色素细胞增殖速度加快,可转化成黑色素瘤细胞。Zhang等[23]发现抑制mGluR1后,影响PI3K/Akt/mTOR信号通路,导致细胞自噬,最终干扰U87MG胶质细胞瘤的生长。
氨作为谷氨酰胺的另一代谢产物,发挥着与谷氨酰胺不同的功能,能通过mTORC1非依赖的途径促进肿瘤细胞的自噬。2者之间看似矛盾却存在其合理性,因为在营养压力或其他压力条件下,通过自噬,肿瘤细胞能循环利用细胞蛋白质和脂类物质,转换成其他生存所需的物质,这反而会有利于肿瘤细胞度过恶劣的肿瘤细胞微环境[24-25]。
3 干预谷氨酰胺代谢在抗肿瘤治疗中的意义
目前已知有不少肿瘤细胞对谷氨酰胺饥饿非常敏感,如胰腺癌、急性淋巴癌和小细胞肺癌等。针对这么这一类对“谷氨酰胺成瘾”的肿瘤,通过干预胞内谷氨酰胺代谢,有利于治疗肿瘤。
3.1 抑制谷氨酰胺的摄入 在肿瘤细胞中,与谷氨酰胺转运相关的蛋白存在上调的现象,利于摄入大量的谷氨酰胺。SLC1A5是负责转运谷氨酰胺入胞的关键转运体之一,可以被L-γ-glutamyl-p-nitroanilide(GPAN)抑制。GPAN使谷氨酰胺依赖的肿瘤细胞缺乏能源物质,最终被饿死[26]。此外,抑制SLC1A5还会影响亮氨酸等必需氨基酸的摄入,使mTOR信号不能激活。
3.2 抑制关键酶阻断谷氨酰胺代谢 谷氨酰胺酶(GLS)是谷氨酰胺代谢中的关键酶,受c-Myc调控,过表达的GLS促进肿瘤细胞增殖。目前已有不少针对GLS的抑制剂,早期用的L-DON(6-diazo-5-oxo-L-norleucine)、Azaserine等,虽然体外使用这类抑制剂会导致细胞增殖速率降低,但特异性较差,有较大的不良反应[27]。最近报道的GLS抑制剂如化合物C-968、BPTES(bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl)ethyl sulfide)等有较好的效果[10,28],正在不同阶段的临床实验得到验证。
3.3 抑制谷氨酰胺代谢参与的信号通路中关键靶点 代谢型谷氨酸受体(mGluRs)的活化有利于ERK/PI3K/Akt信号的激活,从而促进肿瘤的增殖。近来发现一种用于治疗肌萎缩性侧索硬化症(amyotrophic lateral sclerosis,ALS)的药物,即利鲁唑能抑制mGluR1,通过抑制谷氨酸的释放,阻断信号的活化,有望用于治疗肿瘤[29]。
3.4 谷氨酰胺类似物用于诊断 许多肿瘤存在糖代谢异常的现象,于是通过18F-脱氧葡萄糖(18F-FDG)和PET(positron emission tomography)技术的联用可以成像肿瘤患者体内特定部位异常积累的葡萄糖,从而达到诊断的目的[30]。但人体某些部位本身就存在很高的糖利用现象,如脑,使用这种方法会导致成像背景强,无法准确进行诊断[31]。另外,针对某些倾向利用谷氨酰胺的肿瘤细胞,更不适合利用这种成像方法。根据这类肿瘤的特点,发展出了谷氨酰胺类似物4-18F-(2S,2R)-fluoroglutamine(18F-FGln),替代18F-FDG,与PET联用,从而能更准确诊断这类肿瘤[31]。
4 肿瘤的丝氨酸/甘氨酸代谢
除了谷氨酰胺,还有不少氨基酸代谢对肿瘤细胞也是必要的,例如丝氨酸/甘氨酸代谢(一碳单位代谢)。
1955年,丝氨酸从头合成途径首次在肿瘤细胞中被观察到[32],它是葡萄糖代谢通向生物大分子合成的关键节点之一,主要是从糖酵解中间代谢产物3-磷酸甘油酸开始,通过三步酶促反应产生;另外谷氨酰胺来源的谷氨酸与3-磷酸脱氢丙酮酸反应也能合成丝氨酸。在肿瘤细胞中,糖酵解过程中产生的约10%的3-磷酸甘油酸能被磷酸甘油酸脱氢酶(PHGDH)催化成丝氨酸的前体物质3-磷酸脱氢丙酮酸[33]。然后,在磷酸丝氨酸氨基转移酶(phosphoserine aminotransferase 1,PSAT1)和磷酸丝氨酸磷酸酶(phosphoserine Phosphatase,PSPH)的作用下,生成丝氨酸。在三阴性乳腺癌和黑色素瘤细胞中检测到PHGDH的表达量显著上调,敲低PHGDH使丝氨酸从头合成阻断,导致细胞增殖速率显著下降[34]。在葡萄糖或谷氨酰胺饥饿下,Sun等[35]观察到肝癌细胞中丝氨酸从头合成通路是增强的,且相关代谢酶的上调与癌基因c-Myc密切相关。
从外界摄入或从头合成的丝氨酸可被丝氨酸转羟甲基转移酶(serinehydromethyl-transferase,SHMT)催化成甘氨酸,并将脱去的羟基加到四氢叶酸上,形成N5,N10-甲叉四氢叶酸,进入叶酸循环。在甘氨酸合酶的催化下,甘氨酸本身也可以与四氢叶酸反应,形成N5,N10-甲叉四氢叶酸,进入叶酸循环将一碳单位传递下去,促进一碳代谢,最后合成其他氨基酸、脂类和核苷酸等生物大分子[36-37]。而且在这个循环过程会伴随着谷胱甘肽的产生,有助于维持线粒体内氧化还原平衡。但是,直到2012年,才意识到甘氨酸代谢与细胞快速增殖相关。通过敲低SHMT或剥夺外源的甘氨酸会使Hela细胞和其他快速增殖的癌细胞的细胞周期停滞在G1期[38],可见甘氨酸对肿瘤细胞是必要的。目前,临床前的研究发现限制丝氨酸/甘氨酸的摄入能够抑制肿瘤的生长。此外,相关人员还在致力于设计特异性抑制丝氨酸/甘氨酸合成代谢的药物,从而实现对肿瘤的抑制。
5 总结与展望
肿瘤代谢重编程一直都是肿瘤研究的焦点,其中肿瘤细胞中的氨基酸代谢渐渐被揭开面纱。本文着重介绍了谷氨酰胺代谢重编程对肿瘤增殖所发挥的作用,除了提供碳源,还提供氮源,甚至参与细胞信号转导,见图1。目前已有不少针对谷氨酰胺代谢设计的药物用于抗肿瘤治疗,另外针对谷氨酰胺代谢发展出的18F-FGln PET成像,在一定程度上弥补了18F-FDG PET成像的缺陷。而丝氨酸/甘氨酸代谢对肿瘤的意义也在不断地被阐明,针对这一代谢设计药物也将会有很好的抗肿瘤前景。此外,还有许多氨基酸在肿瘤发生发展过程中发挥着重要作用,如亮氨酸等支链氨基酸可在一系列代谢酶的催化下代谢成乙酰辅酶A,最后可以进入TCA循环。
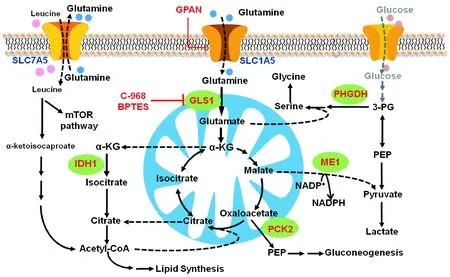
图1 肿瘤的氨基酸代谢Fig.1 Amino acid metabolism in cancer
志谢 本文得到国家科技部、自然科学基金委、中科院以及中国科学技术大学985高校平台的支持。
[1] Hanahan D,Weinberg RA.Hallmarks of cancer:the next generation[J].Cell,2011,144(5):646-674.
[2]Warburg O.On the origin of cancer cells[J].Science,1956,123(3191):309-314.
[3]Warburg O,Wind F,Negelein E.The metabolism of tumors in the body[J].J Gen Physiol,1927,8(6):519-530.
[4]Lu W,Pelicano H,Huang P.Cancer metabolism:is glutamine sweeter than glucose?[J].Cancer Cell,2010,18(3):199-200.
[5]Bhutia YD,Babu E,Ramachandran S,et al.Amino acid transporters in cancer and their relevance to “glutamine addiction”:novel targets for the design of a new class of anticancer drugs[J].Cancer Res,2015,75(9):1782-1788.
[6]Bhutia YD,Babu E,Prasad PD,et al.The amino acid transporter SLC6A14 in cancer and its potential use in chemotherapy[J].Asian J Pharm Sci,2014,9(6):293-303.
[7]Nicklin P,Bergman P,Zhang B,et al.Bidirectional transport of amino acids regulates mTOR and autophagy[J].Cell,2009,136(3):521-534.
[8]Perez-Gomez C,Campos-Sandoval JA,Alonso FJ,et al.Co-expression of glutaminase K and L isoenzymes in human tumour cells[J].Biochem J,2005,386(Pt3):535-542.
[9]Gao P,Tchernyshyov I,Chang TC,et al.c-Myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism[J].Nature,2009,458(7239):762-766.
[10]Wang JB,Erickson JW,Fuji R,et al.Targeting mitochondrial glutaminase activity inhibits oncogenic transformation[J].Cancer Cell,2010,18(4):397-397.
[11]Suzuki S,Tanaka T,Suyama K,et al.Phosphateactivated glutaminase(GLS2),a novel p53-inducible regulator of glutamine metabolism and reactive oxygen species[J].Endocr Rev,2010,107(16):7461-7466.
[12]Hu WW,Zhang C,Wu R,et al.Glutaminase 2,a novel p53 target gene regulating energy metabolism and antioxidant function[J].Proc Natl Acad Sci U S A,2010,107(16):7455-7460.
[13]Xiao DB,Ren P,Su HX,et al.Myc promotes glutaminolysis in human neuroblastoma through direct activation of glutaminase 2[J].Oncotarget,2015,6(38) 655-666.
[14]Vincent EE,Sergushichev A,Griss T,et al.Mitochondrial phosphoenolpyruvate carboxykinase regulates metabolic adaptation and enables glucose-independent tumor growth[J].Mol Cell,2015,60(2):195-207.
[15]Metallo CM,Gameiro PA,Bell EL,et al.Reductive glutamine metabolism by IDH1 mediates lipogenesis under hypoxia[J].Nature,2012,481:380-384.
[16]Mullen AR,Wheaton WW,Jin ES,et al.Reductive carboxylation supports growth in tumour cells with defective mitochondria[J].Nature,2012,481:385-388.
[17]Pavlova NN,Thompson CB.The emerging hallmarks of cancer metabolis[J].Cell Metab,2016,23(1):27-47.
[18]Gaglio D,Soldati C,Vanoni M,et al.Glutamine deprivation induces abortive S-phase rescued by deoxyribonucleotides in K-Ras transformed fibroblasts[J].Plos One,2009,4(3):e4715.
[19]Liou GY,Storz P.Reactive oxygen species in cancer[J].Free Radical Res,2010,44(5):479-496.
[20]Schumacker PT.Reactive oxygen species in cancer:a dance with the devil[J].Cancer Cell,2015,27(2):156-157.
[21]Teh J,Chen S.Metabotropic glutamate receptors and cancerous growth[J].WIREs Membr Transp Signal,2012,1(2):211-220.
[22]Pollock PM,Cohen-Solal K,Sood R,et al.Melanoma mouse model implicates metabotropic glutamate signaling in melanocytic neoplasia[J].Nat Genet,2003,34(1):108-112.
[23]Zhang C,Yuan XR,Li HY,et al.Anti-cancer effect of metabotropic glutamate receptor 1 inhibition in human glioma U87 cells:involvement of PI3K/Akt/mTOR pathway[J].Cell Physiol Biochem,2015,35(2):419-432.
[24]Shanware NP,Mullen AR,DeBerardinis RJ,et al.Glutamine:pleiotropic roles in tumor growth and stress resistance[J].J Mol Med-Jmm,2011,89(3):229-236.
[25]Mizushima N,Levine B,Cuervo AM,et al.Autophagy fights disease through cellular self-digestion[J].Nature,2008,451(7182):1069-1075.
[26]Geldermalsen MV,Wang Q,Nagarajah R,et al.ASCT2/SLC1A5 controls glutamine uptake and tumour growth in triple-negative basal-like breast cancer[J].Oncogene,2016,35(24):3201-3208.
[27]Hensley CT,Wasti AT,DeBerardinis RJ.Glutamine and cancer:cellbiology,physiology,and clinical opportunities[J].J Clin Invest,2013,123(9):3678-3684.
[28]Le A,Lane AN,Hamaker M,et al.Glucose-independent glutamine metabolism via TCA cycling for proliferation and survival in B cells[J].Cell Metab,2012,15(1):110-121.
[29]Shanware NP,Mullen AR,DeBerardinis RJ,et al.Glutamine:pleiotropic roles in tumor growth and stress resistance[J].J Mol Med-Jmm,2011,89(3):229-236.
[30]Abouzied MM,Crawford ES,Nabi HA.18F-FDG imaging:pitfalls and artifacts[J].J Nucl Med Technol.2005,33(3):145-155.
[31]Venneti S,Dunphy MP,Zhang HW,et al.Glutamine-based PET imaging facilitates enhanced metabolic evaluation of gliomas in vivo[J].Sci Transl Med,2015,7(274):274ra17.
[32]Kit S.The biosynthesis of free glycine and serine by tumors[J].Cancer Res,1955,15(11):715-718.
[33]DeBerardinis RJ.Serine metabolism:sometumors take the road less traveled[J].Cell Metab,2011,14(3):285-286.
[34]Possemato R,Marks KM,Shaul YD,et al.Functional genomics reveal that the serine synthesis pathway is essential in breast cancer[J].Nature,2011,476(7360):346-350.
[35]Sun LC,Song LB,Wan QF,et al.cMyc-mediated activation of serine biosynthesis pathway is critical for cancer progression under nutrient deprivation conditions[J].Cell Res,2015,25(4):429-444.
[36]Locasale JW.Serine,glycine and one-carbon units:cancer metabolism in full circle[J].Nat Rev Cancer,2013,13(8):572-583.
[37]AmelioI,Cutruzzola F,Antonov A,et al.Serine and glycine metabolism in cancer[J].Trends Biochem Sci,2014,39(4):191-198.
[38]Jain M,Nilsson R,Sharma S,et al.Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation[J].Science,2012,336(6084):1040-1044.
(编校:吴茜)
作 者 简 介

高平,博士,中国科学技术大学教授,博士生导师,国家重大科学研究计划项目973首席科学家。2003年于日本大阪大学医学院获博士学位,2003~2005年在美国国立癌症研究所从事博士后研究,曾于2005年获得NIH的杰出研究员奖。2005~2011年,在美国约翰霍普金斯大学医学院任博士后及助理研究员,曾获得约翰霍普金斯大学医学院2008年度的青年研究者奖 (Young Investigator Award)。致力于肿瘤代谢相关的癌症研究,曾在Nature,CancerCell,NatureCommunications,EMBOJ等重要期刊上发表论文50多篇,论文被国内外同行引用3000次以上。
The alterations and significance of amino acid metabolism in cancers
SU Lan-hong, SUN Lin-chong, GAO PingΔ
(School of Life Science,University of Science and Technology of China, Hefei 230027, China)
Recently, the understanding of metabolic reprogramming in cancer is expanding continuously.Besides glycolysis, researchers are now paying more attention to amino acid metabolism, especially glutamine metabolism.Glutamine is one of the most abundant non-essential amino acids in human body, but it is a “conditionally essential” amino acid in various types of cancer cells.Thus, targeting glutamine metabolism is becoming a promising strategy for the development of novel cancer therapeutic agents.In addition, another gradually clarified metabolic pathway is serine and glycine metabolism.In this review, we will focus on the alterations, the underlying mechanisms, and significance of glutamine as well as serine and glycine metabolism.We will also discuss cancer therapy opportunities through targeting amino acid metabolism.
cancer; metabolism; glutamine
10.3969/j.issn.1005-1678.2016.09.002
苏兰鸿,女,博士在读,研究方向:肿瘤微环境与代谢,E-mail:lanhongsu@163.com;高平,通信作者,教授、博士生导师,研究方向:肿瘤代谢相关的癌症,E-mail:pgao2@ustc.edu.cn。
R73
A
猜你喜欢
中国农业科学(2022年12期)2022-06-28
中国畜牧杂志(2022年1期)2022-01-20
南昌大学学报(医学版)(2021年1期)2021-11-29
中国药剂学杂志(网络版)(2021年1期)2021-02-24
食品与生物技术学报(2021年2期)2021-01-16
青岛大学学报(医学版)(2020年6期)2020-11-16
中国人兽共患病学报(2018年10期)2018-12-08
中国当代医药(2018年21期)2018-11-10
江苏农业科学(2016年1期)2017-05-17
家庭医学(2015年6期)2015-07-03
