
以“科学思维逻辑”组织“噬菌体侵染细菌实验”的教学建议
2016-08-21 02:38黄建华
生物学教学 2016年9期
黄建华
(江苏省南通大学附属中学 226019)
1 考证“噬菌体侵染细菌实验”的科学思维逻辑
1952年,赫尔希(A.D.Hershey)和蔡斯(M.Chase)所做“噬菌体侵染细菌的实验”是在前人研究的基础上完成的。
1.1 噬菌体的早期研究为同位素标记噬菌体提供了依据 英国人Twort和法国人D′Herelle分别于1915年和1917年发现一种能使细菌裂解的过滤因子,命名为噬菌体。进一步的研究发现,噬菌体是一种病毒,感染细菌后合成子代噬菌体的原料全部来自宿主细胞。另外,已知硫元素只存在于蛋白中,磷元素几乎都存在于核酸中,这为赫尔希和和蔡斯用35S和32P分别标记噬菌体的蛋白质和核酸提供了依据。
1.2 T2噬菌体一步生长曲线的研究为噬菌体侵染细菌实验的时间控制提供了理论依据 德国人M·Delbrück(噬菌体学派创始人)测定了烈性噬菌体T2侵染细菌和成熟噬菌体释放的时间间隔,绘制出每个被侵染细胞释放噬菌体的生长曲线(称为T2噬菌体一步生长曲线),并于1939年发表“噬菌体的生长”一文。T2噬菌体的一步生长曲线表明:T2噬菌体侵染细菌的潜伏期为20 min(潜伏期是指噬菌体感染细菌至细菌释放子代噬菌体的一段时间)。这为赫尔希和蔡斯在实验中将噬菌体感染细菌、搅拌、离心的实验时间控制在20 min之内提供了理论依据。
1.3 静止噬菌体化学形态的研究推测噬菌体的增殖首先要除去保护性的外壳 美国人Anderson于1949年发现用水快速稀释悬浮T2噬菌体的高浓度氯化钠溶液,利用渗透压的骤变可使噬菌体失活,在电子显微镜下可见这种失活的噬菌体呈蝌蚪形的“空壳”。1951年Herriott研究发现渗透压骤变能使噬菌体的DNA释放到溶液中,而“空壳”仍可以吸附到细菌上。这些研究推测噬菌体的增殖首先要除去保护性的外壳。
1.4 噬菌体吸附细菌后DNA对DNA酶变敏感,推测噬菌体吸附后DNA从外壳中排出 Graham和他的同事于1951年发现噬菌体吸附到加热杀死的细菌上后,其DNA对DNA酶变敏感了;Benzer Dulbecco于1952年也发现在不引起生长的缓冲液中,噬菌体吸附到细菌上后,其DNA变得对DNA酶敏感了。据此推测噬菌体吸附到细菌上以后,其DNA就从其保护性外壳中排出,由于失去外壳的保护,故其DNA对DNA酶变敏感。
1.5 噬菌体吸附到细菌碎片上后,其DNA可从外壳中释放出来 在前人实验和推测的基础之上,为了更好地验证噬菌体吸附细菌后DNA从外壳中排出,赫尔希和蔡斯用35S和32P标记的噬菌体吸附细菌碎片(选择吸附细菌碎片能更好的观察到DNA释放的结果),在37℃保温30 min,离心15 min,然后对沉淀和上清液分别进行分析,其结果如表1[1]所示(用同位素的百分比来表示)。
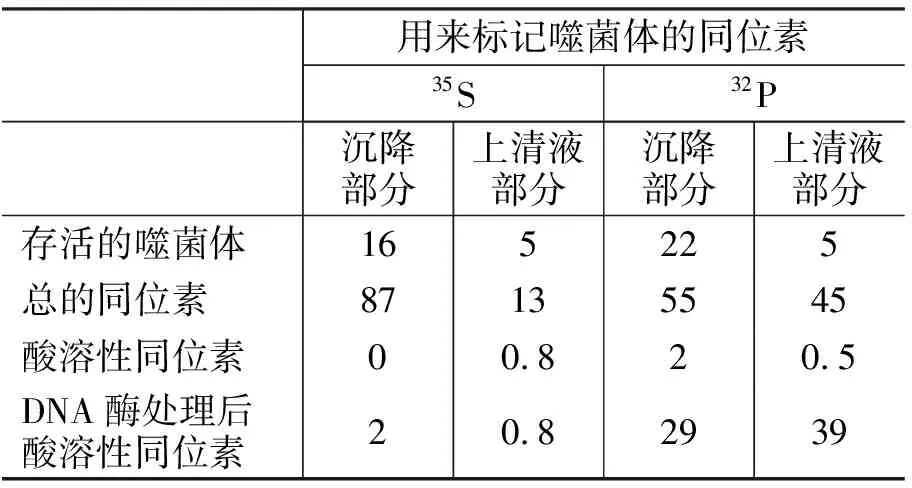
表1 赫尔希和蔡斯同位素标记实验的结果
从35S标记的噬菌体吸附细菌碎片的结果可知:①有87%的噬菌体分布在沉降部分中,说明噬菌体的蛋白尾部吸附在细菌碎片上(因没有捣碎搅拌),其中16%是存活的噬菌体(表明噬菌体吸附细菌后有少部分仍保持活性);②有13%的噬菌体(分布在上清液中)没有吸附到细菌碎片上,其中5%是存活的噬菌体。
从32P标记的噬菌体吸附细菌碎片的结果可知:①有55%的总噬菌体DNA随细菌碎片沉降到底部,其中22%是存活的噬菌体;29%对DNA酶敏感(能被DNA酶分解),说明这部分噬菌体吸附细菌后释放出DNA并失活;酸溶性同位素占2%,说明其未释放DNA;②有45%的总噬菌体DNA分布在上清液中,其中5%存在于存活的噬菌体中;剩下的约40% DNA基本都对DNA酶敏感(39%能被DNA酶分解)。
上述实验数据说明,噬菌体的蛋白尾部起吸附作用;噬菌体吸附细菌后,大部分释放DNA并失活;小部分不失活;还有些失活,但不释放DNA。
1.6 搅拌可从受侵染的细菌上去掉噬菌体外壳 1951年,Anderson通过电子显微镜发现T2噬菌体是靠它的尾部吸附到细菌上去的。假定这种吸附不稳定,搅拌应该能将吸附在菌体上的噬菌体脱落下来。赫尔希和蔡斯用35S和32P标记的噬菌体在吸附培养基中侵染细菌,离心除去未吸附的噬菌体,然后将菌体放在一定的溶液(MgSO41 mM;CaCl20.1 mM;明胶 0.1 g;水 1000 mL)中,在waring捣碎器中搅拌,每隔60 s取样,用抗噬菌体血清进行滴定,测出细菌产生的噬菌体数目,并离心测定同位素的比例及受侵染细菌的存活曲线[1](图1)。
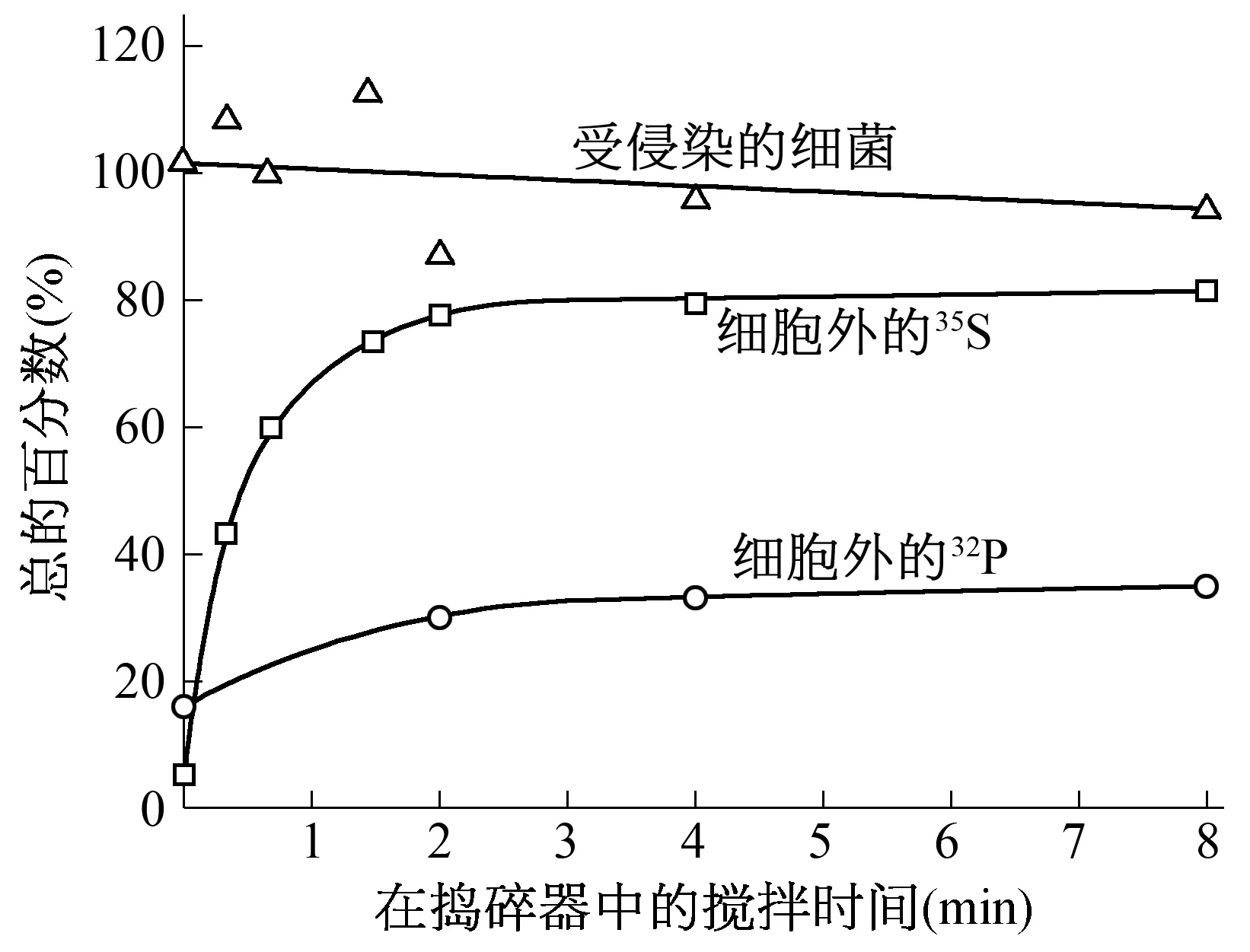
图1 搅拌中测定的同位素比例及受侵染细菌的存活曲线
从实验结果可知:在不搅拌的情况下,就有部分35S和32P自发脱落下来;而搅拌可以把75%左右的35S剥落下来;在这些硫释放的同时,只有35%左右的噬菌体32P释放出来。赫尔希和蔡斯还做了高侵染倍数实验(增加噬菌体的感染量做重复实验),发现高侵染倍数对32P的自发释放影响不大,却大大提高35S的自发释放。这表明,噬菌体在侵染细菌的过程中,其所含的硫(蛋白质)大部分留在细菌的表面;所含的磷(DNA)大部分在吸附细菌后很快进入细胞。
1.7 硫和磷从亲代噬菌体向子代噬菌体的传递说明含硫蛋白质在噬菌体增殖中不起作用,而DNA起作用 赫尔希和蔡斯用放射性35S标记的噬菌体侵染细菌,产生的子代噬菌体中所含的放射性,还不到亲本的1%(可能是吸附在细菌表面存活的亲代噬菌体中标记的硫);用32P标记的噬菌体侵染细菌,所产生的子代噬菌体所含的放射性达到亲本的30%以上。1952年,Watson和Maalos研究发现:侵染性噬菌体DNA中的磷和腺嘌呤以相当大的量而且同等程度传递给子代噬菌体。所以赫尔希和蔡斯推断:含硫蛋白质在噬菌体增殖中不起作用,起作用的是DNA。
1.8 赫尔希和蔡斯当时留下的讨论问题 赫尔希和蔡斯在发表的论文“噬菌体生长过程中蛋白质和核酸各自的功能”中,讨论部分明确提出未弄清楚的问题:除了DNA之外,噬菌体还有没有其他种类的非硫的物质进入细胞?假如有,那么它能传递给子代噬菌体吗?磷传给子代是直接的还是间接的?[2]
从上可知,赫尔希和蔡斯在前人研究的基础之上,设计了“噬菌体侵染细菌实验”,逐步深入研究噬菌体生长过程中蛋白质和核酸各自的功能,其结论是蛋白质担负着使噬菌体吸附到细菌上并使DNA注入细胞的责任,对胞内噬菌体的增殖没有作用,而是DNA在起作用。
2 合理处理“噬菌体侵染细菌实验”的教学建议
2.1 完善教材中的实验步骤,尊重科学探究史实 高中生物学教材中介绍“噬菌体侵染细菌实验”的步骤为:标记→侵染→搅拌→离心→检测。建议根据科学史实将步骤完善为:标记→侵染→离心→培养→搅拌→离心→检测,侵染后离心的目的是去除未吸附的噬菌体,排除未吸附的噬菌体对实验结果的干扰。完善实验步骤一方面尊重科学探究史实;另一方面利于实验结果的准确分析。
2.2 准确分析实验结果,校正现行分析偏差35S标记噬菌体侵染细菌实验中,离心后沉淀物中也有较弱放射性原因的分析,一些资料、试题认为是搅拌不彻底,部分吸附的噬菌体外壳未脱落下来导致的。其实不是搅拌不彻底,而是在保证细菌结构稳定的前提下,搅拌不可能将吸附在细菌表面的噬菌体全部剥落下来(大约剥落75%),导致部分吸附在细菌表面的噬菌体外壳随细菌沉降,使沉淀物中也有较弱的放射性。
32P标记噬菌体侵染细菌实验中,离心后上清液中也有较弱放射性原因的分析,一些资料、试题认为是培养时间控制不当导致的,培养时间过短导致一些噬菌体还没有吸附细菌,就离心到上清液中;或培养时间过长导致细菌裂解释放的子代噬菌体处于上清液中,这样的解释是不科学的。赫尔希和蔡斯实验的实际步骤是:标记→侵染→离心→培养→搅拌→离心→检测。侵染后离心的目的就是去除未吸附的噬菌体,所以上清液中的放射性不是未吸附细菌的噬菌体携带的。在噬菌体一步生长曲线的基础上,赫尔希和蔡斯严格控制实验时间在20 min内。离心前细菌处于潜伏期,尚未裂解。
所以,上清液中的放射性也不是细菌裂解后释放的子代噬菌体携带的,而是吸附细菌后自发脱落的完整噬菌体(未注入DNA)所携带的。
2.3 牢牢抓住证据力点,理清实验递进关系 1944年艾弗里就已证明DNA是使R型细菌转化为S菌的转化因子,即DNA控制细菌荚膜的形成,证明了DNA能控制生物性状,从而证明DNA是S菌的遗传物质。“噬菌体侵染细菌实验”证明亲代DNA能传给子代,在“格里菲斯体外转化实验”的基础上,证明了DNA在亲子间的传递性,成为DNA是遗传物质的更有力证据。赫尔希和蔡斯的实验结果中,细菌裂解释放的子代噬菌体显示,32P标记的一组实验中可以检测到32P标记的DNA,35S标记的一组实验中却不能检测到35S标记的蛋白质,正说明噬菌体DNA在亲子代间的可传递性,是证明DNA是遗传物质的更有力证据的力点,也是“噬菌体侵染细菌实验”和“格里菲斯体外转化实验”呈递进关系的承接点,分析时一定要牢牢抓住。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
中国动物传染病学报(2021年3期)2021-07-21
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
广西林业科学(2016年3期)2016-03-16
广东海洋大学学报(2015年3期)2015-12-22
爱你(2015年16期)2015-04-24
沈阳医学院学报(2014年4期)2014-12-27
渭南师范学院学报(2014年11期)2014-03-20
