
烟草氨基酸代谢及其调控机制
2016-09-13 08:20黎旺姐崔明昆
安徽农业科学 2016年21期
黎旺姐, 李 勇, 崔明昆, 张 刚, 龚 明*
(1.云南师范大学生命科学学院,生物能源持续开发利用教育部工程研究中心,云南省生物质能与环境生物技术重点实验室,云南昆明 650500;2.云南省烟草农业科学研究院,云南昆明 650031)
烟草氨基酸代谢及其调控机制
黎旺姐1, 李 勇2, 崔明昆1, 张 刚1, 龚 明1*
(1.云南师范大学生命科学学院,生物能源持续开发利用教育部工程研究中心,云南省生物质能与环境生物技术重点实验室,云南昆明 650500;2.云南省烟草农业科学研究院,云南昆明 650031)
综述了植物氨同化以及烟草中苯丙氨酸代谢、脯氨酸代谢、美拉德反应及其与烟叶风味和品质关系的研究进展,并分析了环境因素对烟草氨基酸代谢的影响,为深入研究氨基酸对烟叶品质和风味影响的机制奠定了基础。
氨基酸;氨同化;苯丙氨酸和脯氨酸代谢;美拉德反应;烟草
氮作为必需的最重要矿质元素之一,参与植物的生长发育、生化合成、能量转换以及信号转导等生理生化途径,是促进植物生长发育和新陈代谢的重要因子。大量的氮被植物体所需要,通过合成氨基酸等途径来形成蛋白质等生物大分子[1]。氮代谢是植物体中重要的生理过程,是植物生长发育所需的基本物质的基础来源。近年来,生物化学、分子生物学以及其他相关学科和各种研究技术的发展使氮代谢研究取得很大发展。氨基酸是植物体内必不可少的化合物,是合成各类蛋白质的前体物质,因此,氨基酸合成及其代谢是植物生长发育中重要的生理生化过程。高等植物中,氨同化可生成氨基酸以及其他含氮化合物,是氨基酸的重要来源,此外,植物体还可直接从土壤中吸收铵盐来合成氨基酸[2]。烟草中的氨基酸不仅是生长发育所必需的物质,也是重要的与烤烟品质与风味相关的重要物质。笔者综述了植物硝酸盐及铵盐转化为氨基酸的途径、氨基酸与烤烟品质和风味的关系,以及几种与烤烟风味有关的重要氨基酸的代谢及调控机制,旨在为全面深入认识氮素代谢、重要氨基酸的代谢途径以及氨基酸与烟叶品质和风味的关系提供理论依据。
1 植物氨基酸的代谢
1.1植物氮同化土壤中的铵盐可直接转化为氨基酸被植物吸收,而另外一些无机氮却需要转化为有机氮才能被植物吸收利用[3]。铵盐是植物氮的重要来源,另外,氨同化在植物的氮代谢过程中的意义重大[4]。植物体可通过对土壤中的氮素同化过程来吸收并利用土壤中的氮源,但是有些木本植物却不能及时利用最初通过土壤中氮同化作用所吸收来的氮,所以会先合成蛋白并通过储存蛋白的形式将氮源储藏起来[5-6]。Sims等[7]通过假丝酵母试验表明谷氨酸脱氢酶催化氨形成谷氨酸并认为此途径是氨进入氨基酸途径的唯一方式。植物体内的NH4+有的来自NH4+与NO3-这些离子是根通过细胞质膜上的专一性转运子吸收而来的,另外,还有的来自植物根瘤菌的固氮作用、光呼吸作用、氨基酸代谢等[5]。氨同化途径如图1所示[8]。
1.2植物体内硝酸盐和铵盐同化为氨基酸过程中的相关酶及其调控植物体内的氮同化为氨基酸的过程中有众多酶的参与,包括谷氨酰胺合成酶(GS)、谷氨酸合酶(GOGAT)、谷氨酸脱氢酶(GDH)等,这些酶在植物的不同生长发育阶段含量不同,另外,不同的组织器官中含量也有所差异[3,5]。GS是氨同化的关键酶,在ATP存在时,NH4+和谷氨酸在GS的催化下转化成谷氨酰胺将以无机形式存在的氮转化为植物可以吸收利用的有机氮,作为起关键作用的酶参与氨同化的第1步反应,是植物氮素同化中最重要的酶之一。GS由GS基因家族来调控,GS基因的调控不仅存在于启动子结构的特异性方面,同时还存在于转录水平和翻译水平上[9-10]。GOGAT通过催化α-酮戊二酸和谷氨酰胺形成谷氨酸这个反应来影响植物氮代谢过程,且该反应是和前一个GS催化的反应偶联成GS/GOGAT循环来调控植物氮代谢,是植物氮素代谢的主要途径[11]。GOGAT有以还原型辅酶Ι(NADH)为辅酶定位于植物非光合组织中的NADH-GOGAT和以铁氧还蛋白(Fd)为辅酶定位于叶绿体中的Fd-GOGAT 这2种形式,它们分别通过固定根瘤菌中的氮(豆类植物中)、氮的初吸收或者重吸收氨基酸代谢所生成的氮(NADH-GOGAT)和植物呼吸作用所生成的氨(Fd-GOGAT)来调控植物氮代谢,它们分别由GLT、GLU1,2来调控,有组织特异性和器官特异性[11-12]。GDH是植物氮循环中催化铵盐和α-酮戊二酸生成谷氨酸的关键酶,它普遍存在于植物体中,各组织器官中该酶的含量有差异,特别是随生长发育年龄的增长,作为氨的重吸收利用关键调控因子的GDH含量会增加,除了参与铵盐和α-酮戊二酸反应合成谷氨酸的反应外,GDH还参与谷氨酸的氧化反应,该反应又可生成铵[13]。

图1 植物体内硝酸盐和铵盐同化为氨基酸的主要途径[9]Fig.1 Main pathways in which nitrate and ammonium are assimilated to amino acids in plants
2 烟草中脯氨酸和苯丙氨酸的代谢及其与烟叶风味的关系
氨基酸是烟草重要化合物之一,很大程度上影响着烟叶的品质与风味[14-16],是烟草致香物质的间接前体,某些氨基酸(如苯丙氨酸)可通过反应来自身直接分解为一些诸如苯甲醇、苯乙醇等致香前体物质[17-19]。氨基酸通过脱羧或醛转氨产生某些不参与合成蛋白质的氨基酸、芥子油、生物碱、胺类化合物以及氰苷等,其中占生物碱总含量超过90%的烟碱是烟草重要活性物质[20-21]。鸟氨酸以及精氨酸脱羧反应是烟碱合成的起始步骤,在鸟氨酸脱羧酶或者精氨酸酶的参与下,鸟氨酸以及精氨酸脱羧并合成腐胺,再经过一系列的催化反应最终生成烟碱,烟碱可刺激大脑产生兴奋,烤烟中的烟碱含量是评价烟草品质好坏的重要指标之一[20-22]。
2.1烟草中苯丙氨酸代谢
2.1.1苯丙氨酸代谢途径。苯丙氨酸属于芳香族氨基酸,是烟草致香化合物苯甲醇、苯乙醇的前体[19]。作为植物代谢中最重要的次生代谢之一,在细胞中通过苯丙氨酸代谢途径来直接或间接生成含苯丙烷骨架的物质,超过20%的代谢均会通过该途径[23]。苯丙氨酸的合成首先是通过莽草酸途径,由磷酸烯醇式丙酮酸和赤藓糖-4-磷酸经3-脱氧-D-阿拉伯-庚酮糖酸7-磷酸合酶催化下形成分支酸,又经分支酸变位酶催化先生成预苯酸,再经脱水、脱羧反应进一步生成苯丙酮酸,经转氨酶的作用,苯丙酮酸与谷氨酸通过转氨作用最终生成苯丙氨酸,苯丙氨酸合成及去路如图2所示[24]。苯丙氨酸代谢途径是多酚类物质、生物碱等次生代谢物的通用途径,是这些次生代谢物合成都需要经过的途径。在苯丙氨酸解氨酶(PAL)以及肉桂酸-4-羟化酶(C4H)、多酚氧化酶(PPO)等酶的催化下经过一系列反应形成某些酚类化合物的中间产物(如咖啡酸、阿魏酸、芥子酸等),这些酚类中间产物在4-香豆酸辅酶A连接酶(4CL)以及其他酶的共同作用下进一步反应生成CoA酯或者香豆素、绿原酸、莨菪亭等物质。CoA酯在肉桂醇脱氢酶(CAD)以及香豆醇辅酶A降解酶(CCR)等酶的作用下生成对香豆醇、芥子醇、松柏醇等木质素单体,这些木质素单体再经过氧化物酶(POD)或漆酶氧化最终形成木质素,此外,CoA酯还可经一系列反应形成各种黄酮类物质[25-26]。
2.1.2苯丙氨酸代谢的调控。芳香族氨基酸(包括苯丙氨酸、酪氨酸和色氨酸)不能在动物中合成,但在植物或者微生物中却可以合成,其中DAHP合成酶作为第1个限速酶催化PEP和E4P生成DAHP并由此进入芳香氨基酸合成途径,再经转氨酶作用进入苯丙氨酸途径[24]。在苯丙氨酸代谢途径中,苯丙氨酸解氨酶(PAL)是苯丙氨酸代谢途径中的限速酶,它催化苯丙氨酸代谢途径中首步反应,且广泛存在于植物中[23,27]。PAL可将苯丙氨酸催化成反式肉桂酸,还可促进香豆素、木质素、生物碱及黄酮等次生物质的生成[28]。PAL含量在不同的植物以及物种中不同,另外,不同植物器官组织中所含的PAL也不同[29]。水分、温度等生长环境的不同PAL的含量也会有所变化,PAL是一个四聚体,含4个亚基,没有一个特定的米氏常数[29]。不同组织中都有PAL的同工酶,是一个多基因家族,且不同成员的表达都有特异性,各不相同[30-31]。PAL表达受复杂的调控,包括生长发育的环境因子、产物的反馈调节、组织的特异性等,除此之外,PAL基因表达还会受病原菌、乙烯、机械伤害等因素的影响,在应对这些刺激时,PAL分别使用不同的转录起始位点来应对不同的刺激[32-33]。
苯丙氨酸代谢途径中,C4H是继PAL后的最重要的关键酶,它催化该途径中的第1个氧化反应,在氧以及还原型辅酶Ⅱ(即烟酰胺腺嘌呤二核苷酸,NADPH)的存在下行驶功能并将反式肉桂酸催化生成对-香豆酸,从此进入下游代谢途径[27]。不同植物中所含有的C4H基因拷贝数是有差异的,C4H酶可通过不同的催化活性中心残基(I109、K113、V118等)、不同的肽链的化学修饰、不同的空间构象、不同的磷酸化位点来调控C4H酶的活性[34]。
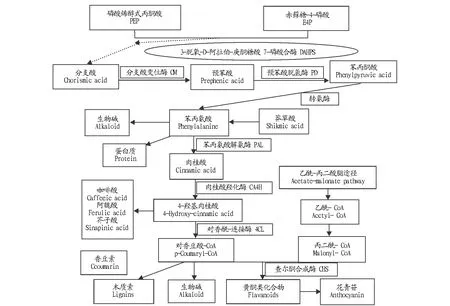
图2 苯丙氨酸合成途径和去路Fig.2 Phenylalanine metabolic pathways in plants
2.1.3苯丙氨酸代谢与烟叶风味的关系。苯丙氨酸是烟叶致香前提物质,通过苯丙氨酸途径合成的多酚类化合物在多酚氧化酶的催化下可发生氧化反应并生成醌类物质,也可通过多酚及其氧化物发生棕色化反应从而得到类黑素等物质,这些物质对烟叶颜色有重要影响[35]。除了影响烟叶色泽,多酚类物质还会影响烟草制品的香气和吃味并呈正相关,多酚也可通过降解来产生一些可使烟草制品拥有优雅的香气的化合物,并且可增加烤烟制品中香气量的物质,从而影响烟叶品质[35]。生物碱中的烟碱(即尼古丁)含量越高,烟叶的香吃味评价也越高,烟碱含量和香气浓度呈正相关,但如果含量太高则会使杂气升高,进而余味就会变差,所以在烟草制品中烟碱含量不能太高[36]。
诸多环境因子会影响苯丙氨酸代谢,温度是调节植物代谢的重要因素,对苯丙氨酸代谢也有很大影响。云南师范大学研究表明,温度对苯丙氨酸代谢下游的多酚代谢有一定影响。特别是对低温的响应很明显[25,37]。另外,光照(包括光质、光强以及日照时间)、水分、氮素水平等因素都对苯丙氨酸代谢有重要影响。
2.2脯氨酸非酶棕化反应及对烟草品质的影响
2.2.1非酶棕化反应对烟草品质的影响。20世纪初法国著名科学家C.C.Maillard把甘氨酸和葡萄糖混在一起,然后进行加热,则形成了褐色物质(类黑精),从此发现了美拉德反应,也称非酶棕化反应,该反应是指还原糖与氨基酸经过缩合、脱水、降解、聚合等一系列反应生成类黑素的过程[38-40]。后来随着在食品中的应用,美拉德反应被定义为羰基化合物和氨基化合物(氨基酸、肽类化合物和蛋白质)所发生的复杂反应[41]。在日常食品贮藏和加工过程中,美拉德反应被广泛应用,用于烟草可使烟草香气更加优美、口味醇和宜人[39]。在烟草上,美拉德反应一般发生在烘烤及加工过程中,美拉德反应可生成优美的天然香气,随着人民生活水平的提高,对香烟的口感、香气等舒适度的要求也越来越高,同时,在烟草行业的降焦减害的要求也越来越高,烟草工业中通过美拉德反应产生多种重要的烟草风味前提物质,卷烟中0.5%~2.0%是美拉德反应的重量,其产物与烟的香味相互协调在一起,可修饰并圆润烟气,因此,美拉德反应产物在烟草制品的增香领域中的研究也越来越受重视[40,42-44]。同时,美拉德反应产物通过热裂解可产生诸如烟气中使尼古丁的含量增加的嘧啶、氨基类化合物等碱性化合物,修饰并增加烟香,增加烟民的劲头和满足感,进而烟民会减少吸烟量,符合降焦减害的要求[45]。
反应温度和pH是影响美拉德反应的重要外界条件,一般来说,温度升高可加快美拉德反应速率,在适宜pH条件下pH升高会加快美拉德反应速率,pH太高则会降低美拉德反应速率[46]。有研究表明,满足反应条件温度105 ℃、时间4 h、pH10、乙醇体积分数为40%时,美拉德反应可有效增加烟草香气[47]。
2.2.2烟草脯氨酸代谢及其抗渗透胁迫的重要作用和对烟叶品质的影响。脯氨酸是蛋白质的一个结构组成部分,是贮存氮的化合物, 是植物正常发育过程中重要的物质[48]。另外,脯氨酸是烤烟中含量最多的氨基酸之一,在烟草根、茎、叶、花等器官中都含有脯氨酸,同时它是极易溶于水的氨基酸,也是一种小分子渗透调节物质,脯氨酸可稳定细胞的亚细胞结构并清除自由基,是渗透调节物质,所以通过积累它可有效应对渗透胁迫的恶劣环境,在受干旱、高盐等渗透逆境胁迫下会在植物体内大量积累,可稳定蛋白质的结构、调节细胞的渗透势,在解除渗透胁迫后,脯氨酸可以为植物提供碳氮源[49-51]。另外,通过与糖类(果糖、还原糖、葡萄糖等)发生美拉德反应而生成的产物在烤烟风味中有重要贡献[47,52-53]。
脯氨酸的生物合成途径最早在大肠杆菌中被发现,植物中,以谷氨酸(Glu)为底物以及以鸟氨酸(Orn)为底物来合成脯氨酸是得到脯氨酸的2个途径,其中吡咯啉-5-羧酸催化合成酶(P5CS)、△1-二氢吡咯-5-羧酸还原酶(P5CR)是谷氨酸合成途径中的限速和调节酶,ATP存在时,在谷氨酸激酶的催化下,首先得到γ-谷氨酰磷酸,接着由L-谷氨酸-γ-半醛(GSA)脱氢酶催化反应并生成L-谷氨酸-γ-半醛,后在△1-二氢吡咯-5-羧酸合成酶(P5CS)作用下发生自环化并形成△1-二氢吡咯-5-羧酸,接着经△1-二氢吡咯-5-羧酸还原酶(P5CR)作用最终生成脯氨酸,是高等植物中合成脯氨酸的主要途径;δ-氨基转移酶(δ-OAT)是调节鸟氨酸代谢途径的限速酶,而ProDH主要负责脯氨酸的降解,是该途径的限速酶,主要负责脯氨酸的降解[49-50,54-55]。植物中脯氨酸通过以谷氨酸为起始底物的谷氨酸途径和以鸟氨酸为底物的鸟氨酸途径来合成,氮源不足时主要通过谷氨酸途径来合成脯氨酸,在具有有足够的氮源时,合成主要通过鸟氨酸途径来完成[56-57]。植物脯氨酸合成及去路如图3所示[58]。
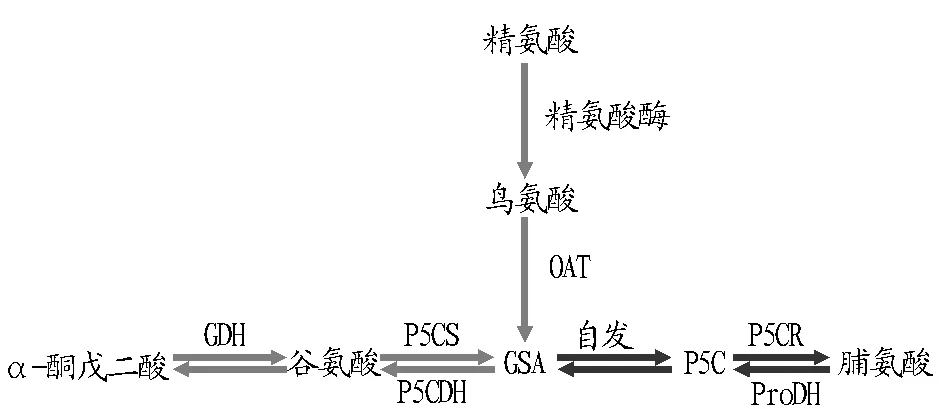
注:GSA.谷氨酰半缩醛; P5C.Δ1-吡咯啉-5-羧酸; P5CS.Δ1-吡咯啉-5-羧酸合成酶; OAT.鸟氨酸转氨酶; ProDH.脯氨酸脱氢酶; P5CR. Δ1-吡咯啉-5-羧酸还原酶; GDH.谷氨酸脱氢酶; P5CDH.吡咯琳-5-羧酸脱氢酶。Note: GSA.glutamic-γ-semialdehyde; P5C.Δ1-pyrroline-5-carboxylate; P5CS.Δ1-pyrroline-5-carboxylate synthetase; OAT.ornithine aminotransferase; ProDH.proline dehydrogenase; P5CR.Δ1-pyrroline-5-carboxylate reductase; GDH.glutamate dehydrogenase; P5CDH.pyrroline-5-carboxylate dehydragenase.图3 植物脯氨酸合成及去路Fig.3 Proline metabolic pathways in plants
脯氨酸的含量占烤烟和香料烟中氨基酸含量的比例很大,是含量最多的氨基酸之一,对烟叶品质的贡献主要也是通过与糖类物质的美拉德反应。有研究表明,果糖和脯氨酸通过非酶棕化反应可使烟草香气增加,同时使杂气和刺激性降低,在生产中,可通过改变反应条件来满足所需的增香效果,要达到对烤烟制品的增香效果,可使果糖与脯氨酸比例为2∶1(重量比),100 ℃下反应2 h,该反应条件是最适反应条件[52];另外,当山梨糖与脯氨酸为1∶3(糖氨摩尔比),pH为6的条件下,于160 ℃下反应5 h所得到的非酶棕化产物对卷烟增加香吃味有很好的效果[40]。
3 环境因素对烟草氨基酸代谢的影响
在烟草中,氨基酸是重要的物质,它不仅是烟株中蛋白质的基本单位,还可以在烤烟调制过程中生成具有挥发性的羰基化合物,有些氨基酸还是烟草中重要的致香物质的前体[19,21]。烟草中的游离氨基酸在不同烤烟品种中含量并不相同,另外,各种环境因子的影响也会导致烟叶中氨基酸含量有差异[59]。烟株在不同的生长发育过程中,氮代谢、烟株衰老过程等均可影响氨基酸的含量[60-61]。有研究表明,在香料烟中,旺长部位含有较高的总游离氨基酸和谷氨酸,这对烟株的生长、抵抗不良环境和抗病都有积极作用[62]。在受到水分缺乏或盐分过高或者其他的渗透胁迫时,脯氨酸在烟叶中的含量会明显增加[63]。在严格控制施肥等因素时,脯氨酸主要受后期加工中调制方式的影响[59]。另外,打顶方式和砍收时期也会影响氨基酸的含量,其中丝氨酸含量随着砍收时期增长而增加,而苏氨酸则相反[59]。肥料作为重要因素,可影响烟株生长发育,其中氮肥是影响烟株生长发育的关键肥料,合理施用氮肥可以得到优质的烟叶,还可提高产量,不同施肥量对红花大金元的各生长指标有重要影响,肥量过低或过高都会影响烟草生长发育,使烟株发育不良[64]。
综上所述,氨基酸是构成蛋白质的基本单位,也是生物体内重要生物活性分子。游离氨基酸是烟叶中的重要化合物,很大程度上影响着烟叶的品质和风味,其含量和种类受到烟草生长发育阶段及外界环境条件的调控,其机制有待进一步深入研究。
[1] YANG X Y,WANG X F,WE M,et al.Response of ammonia assimilation in cucumber seedlingsto nitrate stress[J].Plant Biol,2010,53:173-179.
[2] 徐春英.冬小麦氨基酸代谢与抗旱性关系的研究[D].北京:中国农业科学院,2008.
[3] 李常健,林清华,张楚富.高等植物中氨同化酶及其同工酶研究[J].零陵师范高等专科学校学报,2000(3):20-22.
[4] 阎桂萍,马凤鸣,李文华.甜菜氨同化途径的研究[J].东北农业大学学报,1999(4):318-323.
[5] 彭方仁,郭红彦,杨玉珍,等.木本植物氨同化作用研究进展[J].南京林业大学学报,2006(6):117-122.
[6] 田维敏,闫兴富,胡正海.杨树新梢积累营养贮藏蛋白质的细胞学研究[J].西北植物学报,2003(7):1143-1147.
[7] SIMS A P,FOLES B F.A kinetic study of the assimilation of (15N)-ammonia and the synthesis of amino acids in an exponentially growing culture ofCadidautilis[J].Proc R Soc Lond,1964,159:479-502.
[8] LEAL P J,AZEVEDO R A.Nitrogen use efficiency.2.Amino acid metabolism[J].Annals applied biology,2007,151:269-275.
[9] HOFF T,TRUONG H N,CABOCHE M.The use of mutants and transgenic plants to study nitrate assimilation[J].Plant,cell & environment,1994,17(5):489-506.
[10] 黄冰艳,高伟,苗利娟,等.谷氨酰胺合成酶基因研究进展及其在植物氮代谢调控中的应用[J].中国农学通报,2010(23):53-57.
[11] 卢永恩,罗风,杨猛,等.抑制表达谷氨酸合酶基因对水稻碳氮代谢的影响[J].中国科学生命科学:中文版,2011(6): 481-493.
[12] LAM H M,COSCHIGANO K T,OLIVERIA I C,et al.The molecular-genetics of nitrogen assimilation into amino acids in higher plants[J].Annual review of plant biology,1996,1:569-593.
[13] 黄国存,田波.高等植物中的谷氨酸脱氢酶及其生理作用[J].植物学通报,2001(4):396-401.
[14] 韩锦峰.烟草栽培生理[M].北京:农业出版社,1986:200-250.
[15] 张逸宾,编译.烟草生物化学[M].轻工业部包装科学研究所,1991:25-70.
[16] 杨德廉,王树声,王宝华,等.烟草中氨基酸变化规律及其与烟叶品质关系研究进展[J].中国烟草科学,1998(3):11-13.
[17] HECHT S S.Nitrogenous compounds in cigarette smoke and their possible precursors[J].Rec Adv Tob Sci,1977,3:59-93.
[18] BURTON H R.Effect of curing on the chemical composition of burley tobacco[J].Rec Adv Tob Sci,1983,9:91-153.
[19] KOSUGE T,GILCHRIST D G.Aromatic amino acid biosynthesis and its regulation[J].Biocem Plant,1980,5:507-531.
[20] 李进平,谢志坚,涂书新,等.烤烟烟碱合成及其氮素来源与移栽期和氮肥的关系研究[J].植物营养与肥料学报,2010(3):714-719.
[21] 段胜智,李军营,杨利云,等.烟叶致香物质及其环境影响因子的研究进展[J].贵州农业科学,2015(1):45-52.
[22] FACCHINI P J.Alkaloid biosynthesis in plant:Biochemistry,cell biology,molecular regulation,and metabolic engineering applications[J].Annual review of plant physiology and plant molecular biology,2001,52:29-66.
[23] 李莉,赵越,马君兰.苯丙氨酸代谢途径关键酶:PAL、C4H、4CL研究进展[J].生物信息学,2006(4):187-189.
[24] 李永辉.苯丙氨酸生物合成的研究进展[J].生物技术通讯,2002(4):296-300.
[25] 杨慧芹.温度和水分对烟草多酚代谢的影响及种子引发对提高烟草种子和幼苗抗逆性的效应[D].昆明:云南师范大学,2015.
[26] WEISSHAAR B,JENKINS G I.Phenylpropanoid biosynthesis and its regulation[J].Curr Opin Plant Biology,1998,1:251-257.
[27] 崔建东,李艳,牟德华.苯丙氨酸解氨酶(PAL)的研究进展[J].食品工业科技,2008(7):306-308.
[28] 唐寅,张威威,许锋,等.植物苯丙氨酸代谢相关酶基因启动子研究进展[J].长江大学学报(自然科学版:农学卷),2010(2):68-71.
[29] 欧阳光察,薛应龙.植物苯丙烷类代谢的生理意义及其调控[J].植物生理学通讯,1988(3):9-16.
[30] 程水源,陈昆松,刘卫红,等.植物苯丙氨酸解氨酶基因的表达调控与研究展望[J].果树学报,2003(5): 351-357.
[31] BOLWELL G P,BEII J N,CRAME C L,et al.L-Phenylalanine ammonia-lyase fromPhaseolusvukaris[J].Eur J Biochem,1985,149:419.
[32] 董艳珍.植物苯丙氨酸解氨酶基因的研究进展[J].生物技术通报,2006(S1):31-33.
[33] CHANG A,LIM M H,LEE S W,et al.Tomato phenylalanine ammonia-lyase gene family,highly redundant but strongly underutilized[J].Journal of biological chemistry,2008,48: 33591-33601.
[34] 陈安和.甘蓝型油菜及其亲本物种C4H基因家族克隆及比较基因组学研究[D].重庆:西南大学,2006.
[35] 徐晓燕,孙五三.烟草多酚类化合物的合成与烟叶品质的关系[J].中国烟草科学,2003(1):3-5.
[36] 邓小华,陈冬林,周冀衡,等.湖南烤烟烟碱含量空间分布特征及与香吃味的关系[J].中国烟草科学,2009(5):34-40.
[37] 段胜智.不同生长温度对烟草叶片生长发育及光合特性质体色素和多酚代谢的影响[D].昆明:云南师范大学,2014.
[38] 郭俊成,程晓蕾.美拉德增香调味料研究及其应用[J].中国调味品,1995(6):3-8.
[39] 毕莉莎,李璠,刘志华.美拉德反应产物的抗氧化性及在烟草中应用研究进展[J].云南化工,2010(1):58-61.
[40] 程传玲,杨艳勤,宋辉,等.果糖与羟脯氨酸美拉德反应条件的优化[J].河南农业科学,2014(4):147-151.
[41] 蔡妙颜,肖凯军,袁向华.美拉德反应与食品工业[J].食品工业科技,2003(7):90-93.
[42] LEFFINGWELL J C.Leaf chemistry:Basic chemical constituents of tobacco leaf and differences among tobacco types[M]//DAVIS D L,NIELSEN M T.Tobacco: Production chemistry and technology.Oxford:Blackwell Science,1999:265-284.
[43] BRITT P F,BUCHANAN A C,OWENS C V,et al.Does glucoseenhance the formation of nitrogen containing polycyclic aromatic compounds and polycyclic aromatic hydrocarbons in the pyrolysis of proline[J].Fuel,2004,83:1417-1432.
[44] 李元实,韩民杰,朱良华.梅拉德反应产物在白肋烟处理中的应用[J].烟草科技,2001(4):5-7.
[45] 苏强,冯涛.美拉德反应提高烟草舒适度的研究[D].上海:上海应用技术学院,2014.
[46] 袁媛,刘洁,胡小松,等.温度和pH 值对美拉德模拟体系中丙烯酰胺形成的影响[J].食品与发酵工业,2008(8):32-36.
[47] 彭洁.基于烟草加香的葡萄糖/脯氨酸Maillard反应模型体系研究[D].郑州:河南农业大学,2014.
[48] TROVATO M,MATTIOLI R,COSTANTINO P.Multiple roles of proline in plant stress tolerance and development[J].Rend Lincei-Sci Fis,2008,19:325-346.
[49] 王康,刘艳香,董洁,等.盐胁迫对菊苣幼苗脯氨酸积累及其代谢途径的影响[J].草地学报,2011(1):102-106.
[50] 王丽媛,丁国华,黎莉.脯氨酸代谢的研究进展[J].哈尔滨师范大学自然科学学报,2010(2):84-89.
[51] 朱虹,祖元刚,王文杰,等.逆境胁迫条件TN氨酸对植物生长的影响[J].东北林业大学学报,2009(4):86-89.
[52] 魏明杰,张弘涛,朱智志,等.果糖与脯氨酸的Maillard反应及其在烟草中应用研究[J].安徽农学通报,2009(2):26-28.
[53] 周志磊,徐志强,周顺,等.还原糖与脯氨酸固相美拉德反应模型中焦糖香成分的形成及机理[J].食品与发酵工业,2014(1):30-34.
[54] KAUR G,ASTHIR B.Proline:A key player in plant abiotic stress tolerance[J].Biologia pantarum,2015,4:609-619.
[55] KISHOR P B K,SANGAM S,AMRUTHA R N,et al.Regulation ofprolinebiosynthesis,degradation,up-take and transport inhigher plants:Its implications in plant growth and abioticstress tolerance[J].Current science,2005,88:424-438.
[56] DELAUNEY A J,VERMA D P S.Proline biosynthesis and osmoregulation in plants[J].Plant J,1993,4:215-223.
[57] 许样明,叶和春,李国凤.脯氨酸代谢与植物抗渗透胁迫的研究进展[J].植物学通报,2000(6):536-642.
[58] 邓凤飞,杨双龙,龚明.细胞信号分子对非生物胁迫下植物脯氨酸代谢的调控[J].植物生理学报,2015(10):1573-1582.
[59] 赵田.不同栽培和调制措施对白肋烟和烤烟氨基酸含量的影响[D].郑州:河南农业大学,2012.
[60] 左天觉.烟草的生产、生理和生物化学[M].上海:上海远东出版社,1993.
[61] 史宏志,韩锦峰.不同氮素营养的烟叶氨基酸含量与香吃味品质的关系[J].河南农业大学学报,1997(4):319-322.
[62] 张晨东,殷端,宋玉川,等.香料烟中游离氨基酸含量随施肥量的变化[J].烟草科技,2004(8):38-40.
[63] 汤章城.逆境条件下植物脯氨酸的积累及其可能的意义[J].植物生理学通讯,1984(1):15-21.
[64] 李宏光,赵正雄,杨勇,等.施肥量对烟田土壤氮素供应及烟叶产质量的影响[J].西南师范大学学报(自然科学版),2007(4):37-42.
Amino Acid Metabolism and Its Regulation Mechanism in Tobacco Plants
LI Wang-jie1, LI Yong2, CUI Ming-kun1, GONG Ming1*et al
(1.Engineering Research Center of Sustainable Development and Utilization of Biomass Energyof Ministry of Education,Key Laboratory of Biomass Energy and Environmental Biotechnology of Yunnan Province, School of Life Sciences, Yunnan Normal University,Kunming, Yunnan 650500; 2.Yunnan Academy of Tobacco Agricultural Sciences, Kunming, Yunnan 650031)
The research advances on ammonia assimilation,phenylalanine and proline metabolism, maillard reaction and their relations to quality and flavour of tobacco leave were reviewed, effects of environmental factors on amino acid metabolism were analyzed, which will lay a foundation for deep analysis of effects of amino acids on tobacco quality and flavor.
Amino acids; Ammonia assimilation; Proline and phenylalanine metabolism; Maillard reaction; Tobacco
国家自然科学基金项目(31260064,31460059);国家烟草专卖局科技专项[110201101003(TS-03)]。
黎旺姐(1990- ),女,云南普洱人,硕士研究生,研究方向:植物逆境生物学。*通讯作者,教授,博士,博士生导师,从事植物逆境生物学研究。
2016-06-03
S 572
A
0517-6611(2016)21-015-05
猜你喜欢
世界最新医学信息文摘(2021年24期)2021-04-29
法律史评论(2020年2期)2020-04-13
法律史评论(2020年2期)2020-04-13
学生天地(2019年27期)2019-11-21
赤峰学院学报(蒙文哲学社会科学版)(2018年4期)2018-12-05
三门峡职业技术学院学报(2017年1期)2017-06-05
中国洗涤用品工业(2017年2期)2017-04-16
中国比较医学杂志(2017年5期)2017-01-17
分析测试学报(2015年4期)2016-01-13
医学研究杂志(2015年12期)2015-06-10
