
苗期不同抗性大豆荧光参数及叶绿素含量对大豆花叶病毒胁迫的响应
2016-11-24 07:30齐玉鑫王燕平宗春美孙晓环白艳凤任海祥杜维广侯国强徐德海王晓梅
安徽农业科学 2016年28期
齐玉鑫, 王燕平, 宗春美, 孙晓环, 白艳凤, 任海祥, 杜维广, 侯国强, 徐德海,李 文,王晓梅,张 帅
(黑龙江省农业科学院牡丹江分院/国家大豆改良中心牡丹江试验站,黑龙江牡丹江 157041)
苗期不同抗性大豆荧光参数及叶绿素含量对大豆花叶病毒胁迫的响应
齐玉鑫, 王燕平*, 宗春美, 孙晓环, 白艳凤, 任海祥, 杜维广, 侯国强, 徐德海,李 文,王晓梅,张 帅
(黑龙江省农业科学院牡丹江分院/国家大豆改良中心牡丹江试验站,黑龙江牡丹江 157041)
[目的]研究苗期不同抗性大豆荧光参数和叶绿素含量对大豆花叶病毒(SMV)胁迫的响应。[方法]以感SMV品种合丰25和抗SMV品系牡304为材料,以东北地区SMVⅠ号株系为毒源,测定接种SMV后苗期不同抗性大豆荧光参数及叶绿素含量的变化。[结果]与对照相比,SMV胁迫下,除Fo和qN升高外,其他荧光参数均有所降低,抗SMV品种(系)的变化幅度较小。随着胁迫时间的推移,Fm、Fm’、ΦPSII和Fv/Fm逐渐降低,Fo和qN逐渐升高,qP先降低后升高,且抗SMV品系牡304和感SMV品种合丰25分别在6 d和12 d出现最低值。受SMV胁迫后,叶绿素a和叶绿素b含量的变化趋势相同,抗病品种(系)叶绿素含量的上升幅度大于感病品种(系),感病品种(系)叶绿素含量的降低率大于抗病品种(系)。[结论]苗期荧光参数以及叶绿素含量可作为鉴定大豆对SMV抗性的重要依据。
大豆;SMV胁迫;荧光动力学参数;叶绿素含量
大豆花叶病毒(Soybean mosaic virus,SMV)病是影响世界大豆生产的主要病害之一,也是目前我国东北、黄淮海、长江流域和南方大豆产区最重要的大豆病害之一。大豆花叶病毒隶属马铃薯Y病毒科(Potyviridae)Y病毒属(Potyvirus)大豆花叶病毒种。SMV在大豆植株上表现的症状非常复杂,有多种类型,主要与大豆品种、毒株类型、环境条件有关,也受播期和传播介体的影响。SMV侵染大豆后会导致大豆叶片中叶绿素含量和叶质下降,叶面积减小,影响光合面积和光合能力,植株矮小,生长量下降,单株荚数减少,降低种子百粒重、萌发率、蛋白质含量和含油量,并影响脂肪酸、蛋白质、微量元素及游离氨基酸组分等,最终严重影响大豆产量[1]。感染SMV后的大豆病株籽粒会出现褐斑,严重时病粒率达50%以上[2],当与菜豆荚斑驳病毒(BPMV)复合侵染大豆时出现斑驳的种子比例高达96%[3],严重影响籽粒的外观品质和商品价值。SMV在大豆分枝期至开花期为发病高峰,鼓粒期叶部症状开始潜隐,因此前人研究发现感染SMV后大豆的生理指标变化多从始花期到鼓粒期。在发病初期,大豆植株并无明显症状,对这一时期大豆在SMV胁迫下的生理指标变化鲜见报道。笔者研究了大豆真叶期接种SMV后15 d内叶绿素含量和荧光参数的变化规律,并比较了抗病品种和感病品种在此期间叶绿素含量及荧光参数的差异,旨在为揭示SMV胁迫对不同抗性大豆苗期光合作用的影响以及大豆对SMV的抗性生理机制提供理论依据。
1 材料与方法
1.1 试验材料 选用黑龙江省农业科学院牡丹江分院选育的大豆新品系“牡304”作为抗病材料,该品种以黑农48为母本、以合丰47为父本通过有性杂交获得,2011—2012年的病毒病抗病性鉴定中均表现为抗病;以合丰25为感病材料。供试毒源采用东北地区SMV 1号株系,由黑龙江省农业科学院大豆研究所提供。
1.2 试验设计 试验设置4个处理:Ⅰ为接种SMV的合丰25,Ⅱ为未接种SMV的合丰25,Ⅲ为接种SMV的牡304,Ⅳ为未接种SMV的牡304,3次重复。将供试大豆幼苗栽于营养钵中,接种病毒时期为大豆对生真叶期,接种病毒方法采用人工汁液摩擦法。幼苗接种后在智能温室中培养,设定温度25 ℃,光照18 h;20 ℃,黑暗6 h;从接种第3天开始,每隔6 d对叶绿素含量进行测定,每隔3 d对第1片复叶进行叶绿素荧光参数的测定。
1.3 测定指标与方法 使用调制式荧光仪mini-PAM(由德国WALZ公司生产)测定荧光动力学参数。叶片经暗适应20 min后,首先用弱测定光测定初始荧光Fo,随后给1个强闪光测定最大荧光Fm。然后,在智能温室恒定光源照射下适应20 min,当荧光基本稳定时测定荧光产量Ft,此后加1次强闪测量光适应的样品的最大荧光产量Fm’。
按照下式计算光系统Ⅱ(PSⅡ)最大光化学量子产量:
Fv/Fm=(Fm-Fo)/Fm
(1)
按照以下公式计算PSⅡ实际光化学量子产量:
ΦPSⅡ=(Fm’-Ft)/Fm’
(2)
按照以下公式计算光化学猝灭系数:
qP=(Fm’-Ft)/(Fm’-Fo)
(3)
按照以下公式计算非光化学猝灭系数:
qN=(Fm-Fm’)/(Fm-Fo)
(4)
叶绿素含量的测定采用牛俊义等[4]的乙醇丙酮混合法。
2 结果与分析
2.1 SMV胁迫对大豆苗期荧光参数的影响Fo是在暗适应状态下当PSⅡ 的所有反应中心处于完全开放状态(qP=1)并且所有的非光化学过程处于最小时(qN=0)的荧光产量,其大小与叶绿素浓度有关。陈建明等[5]研究表明Fo的增加可能是植物叶PSⅡ反应中心出现可逆的失活或出现不易逆转的破坏,也可能是植物叶片类囊体膜受到损伤,而且Fo增加量越多,类囊体膜的受损程度就越严重。从表1可以看出,处理 Ⅱ与处理ⅣFo无显著差异,都呈现出逐渐减小的趋势;处理 ⅠFo从第9天开始与其他处理存在显著差异,且逐渐增加;处理 Ⅲ 的Fo变化不大,且与处理Ⅱ、Ⅳ无显著差异。通过对处理Ⅰ与Ⅲ的对比可知,在SMV胁迫下抗病植株Fo增加的幅度要远远小于感病植株,这说明抗病植株受SMV胁迫时PSⅡ 反应中心受到的影响更小。
Fm是在暗适应状态下当PSⅡ的所有反应中心处于完全关闭状态(qP=0)并且所有的非光化学过程处于最小(qN=0)时的荧光产量,Fm大小与QA的氧化还原状态有关;Fm’是在光适应状态下当PSⅡ的所有反应中心处于关闭态(qP=0)并且所有的非光化学过程处于最优态时(qN>0)的荧光产量,二者都可以反映出PSⅡ的电子传递情况。由表1可知,各处理Fm在6 d前无明显差异,9 d后处理 Ⅰ与其他处理开始出现显著差异,处理 Ⅱ、Ⅲ、Ⅳ的Fm无明显变化,处理ⅠFm逐渐下降。处理Ⅰ的Fm’从第3天开始与其他处理存在显著差异,处理 Ⅲ从第12天开始与处理 Ⅰ、Ⅱ存在显著差异,处理 Ⅱ、Ⅳ的Fm’无显著变化,处理 Ⅰ、Ⅲ的Fm’均表现出下降趋势,其中处理Ⅰ下降幅度较大。SMV胁迫下抗病植株和感病植株的PSⅡ反应中心电子传递都受到影响,抗病植株受到的影响小于感病植株。
Ft呈现出不规律变化,但第3天与第15天处理 Ⅰ 的Ft明显降低,而其他3个处理没有明显变化。

表1 SMV胁迫对大豆苗期Fo、Fm、Fm’和Ft的影响
注:相同时间不同处理同一指标标有不同小写字母表示差异显著(P<0.05)。
Note:Different lowercases and capital letters of the same indicators in different treatments at the same time stand for significant difference(P<0.05).
2.2 SMV胁迫下ΦPSⅡ、Fv/Fm、qP与qN的变化ΦPSⅡ表示实际光化学量子产量,可反映PSⅡ反应中心部分关闭情况下实际光能捕获的效率。从图1可以看出,处理 Ⅰ 与处理Ⅲ的ΦPSⅡ显著低于健康植株,处理 Ⅰ 与处理 Ⅲ 的ΦPSⅡ均呈下降趋势,说明在SMV胁迫下抗病植株与感病植株捕获光能的能力都受到了影响。研究表明,受SMV侵染的大豆叶片叶绿素含量下降,这可能是受SMV胁迫时大豆植株捕获光能能力下降的主要因素。
叶绿素荧光参数Fv/Fm表示最大光化学量子产量,可反映PSⅡ反应中心内部光能的转换效率。非胁迫条件下,该参数的变化极小,不受物种和生长条件的影响;胁迫条件下,Fv/Fm明显下降。该研究结果表明,SMV对大豆叶片Fv/Fm有显著影响,SMV胁迫下的大豆植株Fv/Fm比未受胁迫的大豆植株低,处理Ⅰ大豆植株的Fv/Fm呈显著下降趋势,且其下降幅度很大,而处理 Ⅲ 大豆植株的Fv/Fm基本保持稳定(图1)。这说明抗病品种SMV胁迫后,抗病植株PSⅡ 反应中心仍然能够保持较高的活性。

图1 SMV胁迫下Fv/Fm、ΦPSⅡ、qP和qN的变化Fig.1 Changes of Fv/Fm,ΦPSⅡ, qP and qN under SMV stress
光化学淬灭系数反映PSⅡ天线色素吸收的光能用于光化学电子传递的份额,若要维持高的光化学淬灭就要使PSⅡ反应中心处于开放状态,因此光化学淬灭系数在一定程度上反映PSⅡ反应中心的开放程度。光化学淬灭系数反映PSⅡ原初电子受体QA的还原状态,它由QA重新氧化形成[6]。光化学淬灭系数qP愈大,PSⅡ的电子传递活性愈大[7]。处理 Ⅰ、Ⅲ的qP均呈现先下降后上升趋势,处理Ⅲ的qP最低值出现在第6天,处理 Ⅰ 的qP最低值出现在第12天(图1)。这表明大豆叶片在SMV胁迫下受到破坏的PSⅡ电子传递是可以逐渐修复的,抗病植株自我修复的能力要优于感病植株。
qN反映PSⅡ线色素吸收的不能用于光合电子传递而以热形式耗散掉的光能部分,它是一种自我保护机制,对光合机构起到一定的保护作用,qN的升高说明PSⅡ正受到破坏。未受SMV胁迫处理 Ⅱ、Ⅳ的qN变化不大,处理Ⅰ的qN出现较明显上升,处理ⅢqN上升幅度小于处理Ⅰ(图1)。这表明随着SMV胁迫时间的增加,PSⅡ天线色素吸收的光能用于光合作用所占的份额在逐渐减少,抗病植株的减小幅度小于感病植株。
2.3 SMV胁迫对大豆苗期叶绿素含量的影响 叶绿素a是光合反应中心复合体的主要组成成分,其中处于特殊状态的反应中心叶绿素a分子是执行能量转化的光合色素,而叶绿素a 是捕光色素蛋白复合体的重要组成部分,其主要作用是捕获和传递光能,它们在活体中大概都是与蛋白质结合在一起,存在于类囊体膜上[8]。由表2可知,4个处理的叶绿素a含量均随着胁迫时间的推移而增加,相同品种(系)受胁迫的处理叶绿素a含量的增加速度要低于未受胁迫的处理;同一品种同一时间不同处理叶绿素a含量差异不大,只有合丰25在15 d受胁迫的处理 Ⅰ 与未受胁迫的处理 Ⅱ 存在显著差异;合丰25叶绿素a含量的降低率随着胁迫时间的推移而增加,牡304的叶绿素a含量则呈现出先下降后上升的趋势,其降低率都在15 d出现明显上升,合丰25叶绿素a含量的上升幅度大于牡304。
由表2可知,叶绿素b含量的变化规律与叶绿素a基本相同。3 d和9 d相同品种的2个处理叶绿素含量并没有明显的变化,15 d相同品种的2个处理叶绿素含量的降低率都明显升高,合丰25在15 d受胁迫的处理 Ⅰ 与未受胁迫的处理 Ⅱ 叶绿素b含量存在显著差异。
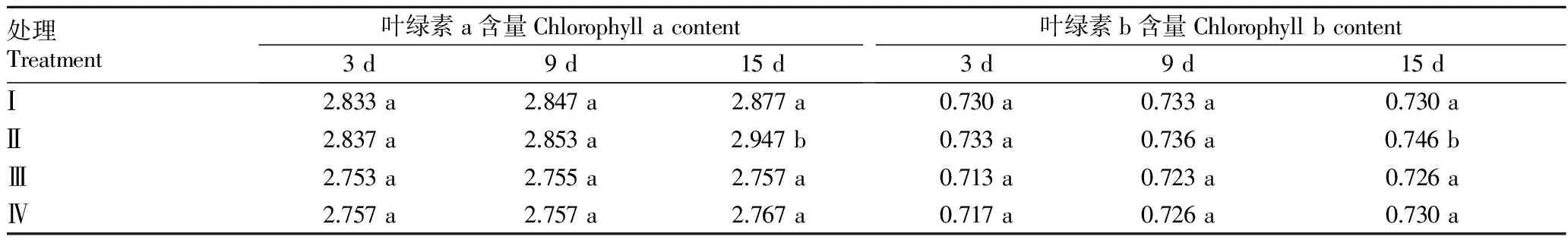
表2 SMV胁迫对大豆苗期叶绿素含量的影响
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercases in the same column stand for significant difference(P<0.05).
3 讨论
陈兵等[9]研究表明,棉花在黄萎病胁迫下,随着病害严重度的增加,棉叶叶绿素 a、叶绿素 b、叶绿素 a+b、叶片含水率,病叶叶绿素荧光参数可变荧光、最大荧光、光系统 Ⅱ 最大光化学效率、光系统 Ⅱ 的潜在活性、最大荧光与最小荧光比、光系统 Ⅱ 的量子产额均减小,而初始荧光增加。柯玉琴等[10]研究表明青枯病侵染的烟草叶片Fv、Fv/Fm、Fv/Fo受到抑制,感病品种抑制程度大于抗病品种。郑国华[11]研究表明感染炭疽病的枇杷叶片Fv、Fv/Fm、Fv/Fo及量子产量受到抑制。杨峰等[12]研究表明受根腐病胁迫后,大豆植株样本叶片荧光参数Fv/Fm、NPQ、ΦPSⅡ均极低于正常植株。穆西玉等[13]研究表明玉米感大斑病后各品种叶片叶绿素含量显著降低,叶绿素含量从高到低依次为感病品种、中抗品种、抗病品种,这与该研究结果相一致。马铃薯Y病毒(PVY)侵染后不同发育期马铃薯叶片的光合参数表明,侵染初期病株的净光合速率(Pn)、Fv/Fm、电子传递速率(ETR) 与健康植株基本相同,随着发育进程的推进,病株的Pn和ETR下降速度显著大于健康株,而Fv/Fm则变化不大[14]。这与该试验结果不同,这可能是因为不同种植物对不同病原菌的抗性差异所造成的。
病害胁迫会造成叶绿体结构的破坏和叶绿素含量的变化,从而影响植物的光合作用。电镜扫描接种SMV Nl株系21 d的细胞超微结构发现,细胞形状异常,少数细胞有质壁分离现象发生,细胞核中核仁融解,叶绿体肿胀变形,淀粉粒减少,叶绿体上嗜锇颗粒增加, 颜色加重,边缘清晰,基粒和基质片层断裂细胞壁附近的线粒体数目增多[15]。Balachandran等[16]研究表明植物感染病毒后除表现为叶绿体遭到破坏、叶绿素含量下降外,一些病程相关蛋白酶的活性得以提高,使正常生长发育受到抑制。植物感染烟草花叶病毒(TMV)和黄瓜花叶病毒(CMV)等病毒后,叶绿体内发生病毒积累,放氧复合体(OCE)中的多肽成分发生变化,光系统 Ⅱ(PSⅡ)电子传递显著降低,同时,病毒外壳蛋白CP积累于叶绿体内光化学反应中心之间,诱导产生光抑制,从而降低植物的光合作用[17]。这些都是病毒侵染植物体后导致其光合生理指标受到影响的因素。
除病害外,生理胁迫[18]和环境污染[19]等也会对植物光合生理产生影响。为避免植株长势不同、其他病害和生理胁迫对研究成果造成影响,试验选用的植株长势一致,培养大豆幼苗所用的土壤高温灭菌,营养钵全部经过紫外线照射消毒,避免出现复合侵染,试验过程中每次用注射器向营养钵中加入等量的蒸馏水,避免出现因水分引起的生理胁迫。
叶绿素荧光动力学方法因其快速、灵敏、无损伤等优点,目前已被广泛用于研究和探测各种逆境对植物光合生理的影响。该研究结果表明,不同抗性的大豆幼苗受SMV胁迫后抗病和感病品种(系)的PSⅡ系统捕捉光能的能力、反应中心活性和电子传递都受到影响,Fv/Fm、ΦPSII、qP、qN有显著差异,抗病品种(系)受到的影响较小,综合分析苗期荧光参数以及叶绿素含量可作为鉴定大豆对SMV抗性的重要依据之一。
[1] 智海剑,胡蕴珠,刘天宇.大豆花叶病毒对大豆生长的影响[J].中国油料,1996(1):44-47.
[2] 刘伟.大豆花叶病毒病[J].大豆科技,1998(2):29.
[3] QUINIONES S S,DUNLEVY J M,FISHER J W.Performance of three soybean varieties inoculated with soybean mosaic virus and bean pod mottle virus[J].Crop science,1971,11(5):662-664.
[4] 牛俊义,杨祈峰.作物栽培学研究方法[M].兰州:甘肃民族出版社,1998:85-88.
[5] 陈建明,俞晓平,程家安.叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J].浙江农业学报,2006(1):51-55.
[6] 鞠正春,于振文.追施氮肥时期对冬小麦旗叶叶绿素荧光特性的影响[J].应用生态学报,2006(3):3395-3398.
[7] 张其德,张建华,刘合芹,等.限水灌溉和不同施肥方式对冬小麦旗叶某些光合功能的影响[J].植物营养与肥料学报,2000(1):24-29.
[8] 张亚黎,罗宏海,张旺锋,等.土壤水分亏缺对陆地棉花铃期叶片光化学活性和激发能耗散的影响[J].植物生态学报,2008(3):681-689.
[9] 陈兵,王克如,李少昆,等.病害胁迫对棉叶光谱反射率和叶绿素荧光特性的影响[J].农业工程学报,2011,27(9):86-93.
[10] 柯玉琴,潘廷国,方树民.青枯菌侵染对烟草叶片H2O2代谢、叶绿素荧光参数的影响及其与抗病性的关系[J].中国生态农业学报,2002,10(2):40-43.
[11] 郑国华.炭疽病侵染对枇杷叶片H2O2含量和叶绿素荧光参数的影响[J].福建农业大学学报(自然科学版),2001,30(3):353-356.
[12] 杨峰,黄山,崔亮,等.玉米/大豆套作下作物叶片氮、磷动态特征及其相关性分析[J].植物营养与肥料学报,2013,19(4):9.
[13] 穆西玉,张海艳,赵延明.粗缩病对不同抗性玉米品种叶片生理特性的影响[J].植物生理学报,2014(6):801-804.
[14] 彭晏辉,雷娟利,黄黎锋,等.马铃薯Y病毒侵染对叶绿体超微结构、光合和荧光参数的影响[J].植物病理学报,2004,34(1):32-36.
[15] 滕卫丽,李文滨,韩英鹏,等.大豆抗感品种(系)接种SMV1叶片细胞超微结构变化的比较[J].作物杂志,2008(1):34-36.
[16] BALACHANDRAN S,OSMOND C B,MAKINO A.Effects of two strains of tobacco mosaic virus on photosynthetic characteristics and nitrogen partitioning in leaves ofNicotianatabacumcv Xanthi during photoacclimation under two nitrogen nutrition regimes[J].Plant Physiol,1994,104(3):1043-1050.
[17] REINERO A,BEACHY R N.Reduced photosystem II activity and accumulation of viral coat protein in chloroplasts of leaves infected with tobacco mosaic virus[J].Plant Physiol,1989,89(1):111-116.
[18] 尹赜鹏,刘雪梅,商志伟,等.不同干旱胁迫下欧李光合及叶绿素荧光参数的响应[J].植物生理学报,2011(5):452-458.
[19] 袁远爽,肖娟,胡艳.模拟酸雨对白簕叶片抗氧化酶活性及叶绿素荧光参数的影响[J].植物生理学报,2014(6):758-764.
Effect of SMV Stress on Chlorophyll Fluorescence Parameters and Chlorophyll Content of Soybean with Different Resistant Levels at Seedling Stage
QI Yu-xin,WANG Yan-ping*,ZONG Chun-mei et al
(Mudanjiang Branch of Heilongjiang Academy of Agricultural Sciences/Mudanjiang Experiment Station of National Soybean Improvement Center,Mudanjiang,Heilongjiang 157041)
[Objective]The aim was to study effect of SMV stress on chlorophyll fluorescence parameters and chlorophyll content of soybean with different resistant levels at seedling stage.[Method] Hefeng25 and Mu304 were used as experiment materials,and virus source was SMVⅠ,effect of SMV stress on chlorophyll fluorescence parameters and chlorophyll content of soybean with different resistant levels at seedling stage was studied.[Result] The result showed that compared with control,under SMV stress,fluorescence parameters declined exceptFoandqNincreasing,and change range of SMV-resistant variety(line) was smaller.Fm,Fm’,ΦPSIIandFv/Fmdeclined,FoandqNincreased andqPfirstly decreased and then increased with the day passing,the minimumqPvalue of Mu304 and Hefeng 25 respectively happened in sixth and twelfth day.Changes of chlorophyll a and b were same under SMV stress,change range of chlorophyll content resistant variety(line)>susceptible variety(line),reduction rate of chlorophyll content resistant variety(line)>susceptible variety(line).[Conclusion] Chlorophyll fluorescence parameters and chlorophyll content at seedling stage can be used as important basis for identifying SMV resistance of soybean.
Soybean; SMV stress; Chlorophyll fluorescence parameters; Chlorophyll content
国家自然科学基金面上项目(31371651);中国科学院大豆分子设计育种重点实验室开放课题(SMDB-02)。
齐玉鑫(1983- ),男,河北沧县人,助理研究员,硕士,从事大豆抗病育种研究。*通讯作者,副研究员,博士,从事大豆抗逆育种研究。
2016-08-03
S 565.1
A
0517-6611(2016)28-0007-04
猜你喜欢
云南化工(2021年9期)2021-12-21
浙江林业科技(2021年3期)2021-07-13
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
中国糖料(2018年4期)2018-07-31
长江大学学报(自科版)(2018年14期)2018-07-17
生物学教学(2017年9期)2017-08-20
食品工业科技(2014年6期)2014-05-10
影像科学与光化学(2014年5期)2014-03-11
东北林业大学学报(2014年11期)2014-03-08
