
新疆小麦 TaLox-B1、 TaLox-B2、 TaLox-B3位点的等位变异及其分布规律
2016-12-29 01:44禹飞雄茹仙古丽牙生吕文娟耿洪伟
麦类作物学报 2016年11期
白 璐,禹飞雄,茹仙古丽·牙生,吕文娟,任 毅,谢 磊,耿洪伟
(1.新疆农业大学科学技术学院,新疆乌鲁木齐 830052; 2.新疆农业大学农学院,新疆农业大学生物技术重点实验室,新疆乌鲁木齐 830052; 3.新疆巴州农业局,新疆库尔勒 841000)
新疆小麦 TaLox-B1、 TaLox-B2、 TaLox-B3位点的等位变异及其分布规律
白 璐1,禹飞雄2,茹仙古丽·牙生3,吕文娟2,任 毅2,谢 磊2,耿洪伟2
(1.新疆农业大学科学技术学院,新疆乌鲁木齐 830052; 2.新疆农业大学农学院,新疆农业大学生物技术重点实验室,新疆乌鲁木齐 830052; 3.新疆巴州农业局,新疆库尔勒 841000)
面粉颜色是小麦品质分级的重要指标。为了给新疆小麦面粉白度改良提供依据,本研究利用位于4BS染色体上的功能标记LOX16、LOX18和Lox-B23,对123份新疆小麦品种(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3 等位变异进行分子标记检测。结果表明,在123份新疆小麦品种(系)中,具有高LOX活性等位基因 TaLox-B1a和低LOX活性等位基因 TaLox-B1b的材料分别有33份(26.8%)和90份(73.2%);具有高LOX活性等位基因 TaLox-B2a和低LOX活性等位基因 TaLox-B2b的材料分别有122份(99.2%)和1份(0.8%);具有高LOX活性等位基因 TaLox-B3a和低LOX活性等位基因 TaLox-B3b的材料分别有95份(77.2%)和28份(22.8%)。在不同类型冬小麦品种(系)中, TaLox-B1a的分布频率表现为引进品种(系)>自育品种(系)>地方品种外, TaLox-B3a的分布频率表现为地方品种>引进品种(系)>自育品种(系);在不同类型春小麦中 TaLox-B1a只存在于晚期品种, TaLox-B2a和 TaLox-B3a的分布频率均表现为早期品种(100.0%)>晚期品种。所检测的新疆小麦品种(系)共有5种基因型组合,具有最高LOX活性的基因型组合 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a,具有中等LOX活性的基因型组合 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b以及具有低LOX活性的基因型组合 TaLox-B1b/ TaLox-B2b/ TaLox-B3b的分布频率分别为23.6%、53.7%、19.5%、2.4%和0.8%。在新疆小麦资源中具有高LOX活性基因型组合的材料的频率高于全国平均水平。本研究将3个位点功能标记组合起来使用,能更有效地评价检测材料的LOX活性水平。
新疆小麦;脂肪氧化酶;功能标记;变异组合
小麦面粉颜色是小麦品质分级的重要指标。小麦籽粒中脂肪氧化酶(lipoxygenase,LOX)活性是影响面粉色泽的重要因素之一,高LOX活性可以提高面粉及其相关产品的白度从而提高其商品价值[1-3]。LOX能够与小麦面粉中的类胡萝卜素发生偶联氧化作用降低黄色素含量,进而使小麦面粉变白[4]。还有研究认为,低 LOX 活性更有助于小麦的贮藏[1],高LOX活性则有助于增强面筋强度[5]。
LOX活性受基因型和环境的影响,但主要受遗传因素的影响[6]。前人研究表明,小麦籽粒LOX活性主要受第4和第5同源群上的基因调控[7-10]。Geng等[11]研究表明,在1AL上的 QLpx.caas-1AL位点和4BS上的 QLpx.caas-4B位点上存在决定LOX活性的2个主效基因,分别与SSR标记 Xwmc312和 Xgwm251紧密连锁。基于4BS上的 TaLox-B1位点基因不同等位变异的序列差异,Geng等[12]开发了互补显性标记LOX16和LOX18,其中,LOX16可在具有等位基因 TaLox-B1a(a指与高LOX活性相关,下同)的材料中扩增出489 bp的片段,而在具有等位基因 TaLox-B1b(b指与低LOX活性相关,下同)的材料中无扩增产物;LOX18在具有等位基因 TaLox-B1b的材料中可扩增出791 bp的片段,而在具有等位基因 TaLox-B1a的材料中无扩增产物。利用这一对互补显性标记,Geng等[12]检测198份中国冬小麦材料,结果表明这一对功能标记能有效应用于分子标记辅助选择。张钰玉等[13]、相吉山等[14]和杨淑萍等[15]利用互补显性标记LOX16和LOX18分别对陕西、新疆和黑龙江小麦材料进行了等位变异检测,结果表明该标记能有效应用于小麦分子育种实践。随后,Zhang等[16]在中国小麦4BS上成功克隆了 TaLox-B2、 TaLox-B3基因,并发现 TaLox-B2基因存在 TaLox-B2a和 TaLox-B2b 两个等位变异,其将 TaLox-B3 基因类型定为 TaLox-B3a变异类型,不含有 TaLox-B3基因的暂定为 TaLox-B3b类型。基于这两个LOX基因,Zhang等[16]开发了共显性标记Lox-B23,能准确区分 TaLox-B2和 TaLox-B3位点的不同等位基因。 Lox-B23在具有 TaLox-B2a/TaLox-B3a基因型的材料中能同时扩增出788 bp和677 bp两种带型,在具有 TaLox-B2a/TaLox-B3b的材料中只能扩增出788 bp的单一带型,而在具有 TaLox-B2b/TaLox-B3b的材料中只能扩增出660 bp的单一带型。对136份中国小麦材料的 TaLox-B1、 TaLox-B2、 TaLox-B3位点进行等位变异检测及LOX活性分析表明,具有最高LOX活性的基因型组合是 TaLox-B1a/TaLox-B2a/TaLox-B3a或 TaLox-B1b/TaLox-B2a/TaLox-B3a,具有中高LOX活性的基因型组合是 TaLox-B1b/TaLox-B2a/TaLox-B3b或 TaLox-B1a/TaLox-B2a/TaLox-B3b,具有最低LOX活性的基因型组合是 TaLox-B1a/TaLox-B2b/TaLox-B3b[16]。
虽然相吉山等[14]利用互补显性标记LOX16和LOX18对新疆小麦种质资源 TaLox-B1位点分布特征做了研究,但目前尚未见有关新疆小麦 TaLox-B2、 TaLox-B3等位变异检测以及三个位点组合基因型的检测及其分布规律的研究报道。因此,本研究利用与LOX 活性相关的LOX16、LOX18和Lox-B23功能标记,对123份新疆麦区具有代表性的品种(系)的三个LOX活性基因 TaLox-B1、 TaLox-B2和 TaLox-B3的等位变异及其分布规律进行研究,以期为改良新疆面粉和面制品白度以及选育高LOX活性品种提供依据。
1 材料与方法
1.1 材料及其来源
参试材料为123份新疆小麦品种(系),其中,冬小麦品种(系)99份(17份为新疆地方品种,39份为引进品种,43份为自育品种),春小麦品种(系)24份(2000年前选育的早期品种4份,2000年后选育的晚期品种20份)。这些材料涵盖新疆主要种植区各个时期种植和使用的冬、春小麦品种,具有较好的代表性。上述小麦材料由新疆农业大学农学院小麦课题组收集保存,其中部分小麦材料由新疆农科院粮食作物研究所冬小麦课题组、新疆农垦科学院作物研究所、新疆农科院核能与生物技术研究所小麦课题组提供。
1.2 基因组DNA的提取
为避免由于单粒种子偶然性的结果错误,每份小麦品种(系)均选取3粒种子,按照Lagudah等[17]的方法提取基因组DNA。DNA提取液包含:200 mmol·L-1Tris-HCl(pH=8.0),25 mmol·L-1EDTA,288 mmol·L-1NaCl 和0.5% SDS。
1.3 PCR扩增与检测
1.3.1 引物
利用Geng等[12]开发的显性标记LOX16和LOX18检测小麦4B染色体上 TaLox-B1基因的等位变异;利用Zhang等[16]开发的共显性标记 Lox-B23检测小麦4B染色体上 TaLox-B2和 TaLox-B3基因的等位变异。引物(表1)均由北京六合华大基因科技股份有限公司合成。
表1 检测目标基因等位变异的引物信息
Table 1 Primers for detecting alleles of target genes

基因Gene标记Marker引物序列(5'-3')Primersequence(5'-3')产物大小Productsize/bp等位变异Allele参考文献ReferenceTaLox-B1LOX16F:CCATGACCTGATCCTTCCCTTR:GCGCGGATAGGGGTGGT489TaLox-B1a[12]LOX18F:GCGCGGATAGGGGTGGTR:GCGCGGATAGGGGTGC791TaLox-B1b[12]TaLox-B23Lox-B23F:TTCCCGCTCCAGCTCATCAAR:AGGGCCTGCATGCAACAAGT788/677788660TaLox-B2a/TaLox-B3aTaLox-B2a/TaLox-B3bTaLox-B2b/TaLox-B3b[16]
1.3.2 PCR的反应体系和扩增程序
PCR反应以基因组DNA为模板,在Eppendorf AG 22331 Hamburg PCR仪上进行。25 μL PCR反应体系:50 ng DNA,TaqDNA Polymerase (2.5 U·μL-1)0.25 μL (北京天根生化科技公司),上、下游引物(10 μmol·L-1)各1 μL,dNTP(25 μmol·L-1) 1 μL(北京天根生化科技公司),10×PCR Buffer 2.5 μL(北京天根生化科技公司),最后用ddH2O补充至25 μL。标记LOX16、LOX18的PCR扩增程序:94 ℃预变性5 min;94 ℃变性45 s,62 ℃退火45 s,72 ℃延伸1 min,共35个循环;最后72 ℃延伸10 min,4 ℃保存。标记Lox-B23的扩增程序:94 ℃预变性4 min;94 ℃变性45 s,退火温度从65 ℃开始每循环降0.3 ℃,退火时间30 s,72 ℃延伸2 min,共35个循环;最后72 ℃延伸10 min,4 ℃保存。
1.3.3 琼脂糖凝胶电泳检测
标记LOX16、LOX18的PCR扩增产物用1.5%的琼脂糖凝胶电泳检测,标记Lox-B23的PCR扩增产物用3.0%的琼脂糖凝胶电泳检测,缓冲体系1×TAE溶液,150 V电压电泳35 min,0.5%溴化乙锭(EB)染色10 min,蒸馏水漂洗后用VILBERLOURMAT凝胶成像系统扫描成像,并存入计算机。
1.4 统计方法
根据每个品种(系)三粒种子DNA检测结果判断该品种的LOX基因 TaLox-B1、 TaLox-B2、 TaLox-B3位点的等位变异类型,对PCR产物条带清晰、单一并符合目标条带大小的结果进行统计,若出现结果不一致的,需重新提取DNA并检测,以3次检测一致的结果为准。
2 结果与分析
2.1 新疆小麦LOX基因 TaLox-B1、 TaLox-B2和 TaLox-B3等位变异类型及分布
用互补显性功能标记LOX16 、LOX18和共显性功能标记 Lox-B23对123份新疆小麦品种(系)进行等位变异检测(图1和图2),结果表明,除 TaLox-B1位点的 TaLox-B1b变异类型的材料有90份(73.2%),为优势等位变异外, TaLox-B2和 TaLox-B3位点分别以 TaLox-B2a和 TaLox-B3a为优势等位变异类型,分别为122份(99.2%)和95份(77.2%)(表2)。99份冬小麦材料在三个位点的变异类型的分布频率也具有上述特点,具有各基因优势等位变异 TaLox-B1b、 TaLox-B2a和 TaLox-B3a的材料分别有69份(69.7%)、99份(100%)和89份(89.9%)(表2)。24份春小麦材料中,除 TaLox-B2位点具有 TaLox-B2a的材料有23份(95.8%),为优势等位变异外, TaLox-B1和 TaLox-B3位点均是以 TaLox-B1b和 TaLox-B3b为优势等位变异,分别为21份(87.5%)和18份(75.0%)(表2)。

M:DL2000;1:冀麦24;2:碧蚂6号;3:济南4号;4:新春27;5:新春29;6:奎花2号;7:新冬17;8:新春17;9:新春35;10:新春37。
M:DL2000;1:Jimai 24;2:Bima 6;3:Jinan 4;4:Xinchun 27;5:Xinchun 29;6:Kuihua 2;7:Xindong 17;8:Xinchun 17;9:Xinchun 35;10:Xinchun 37.
图1 LOX16(a)和 LOX18(b)标记检测部分小麦品种 TaLox-B1位点的等位变异结果
Fig.1 Identification of TaLox-B1 alleles by functional markers LOX16 (a) and LOX18 (b)in part of wheat varieties
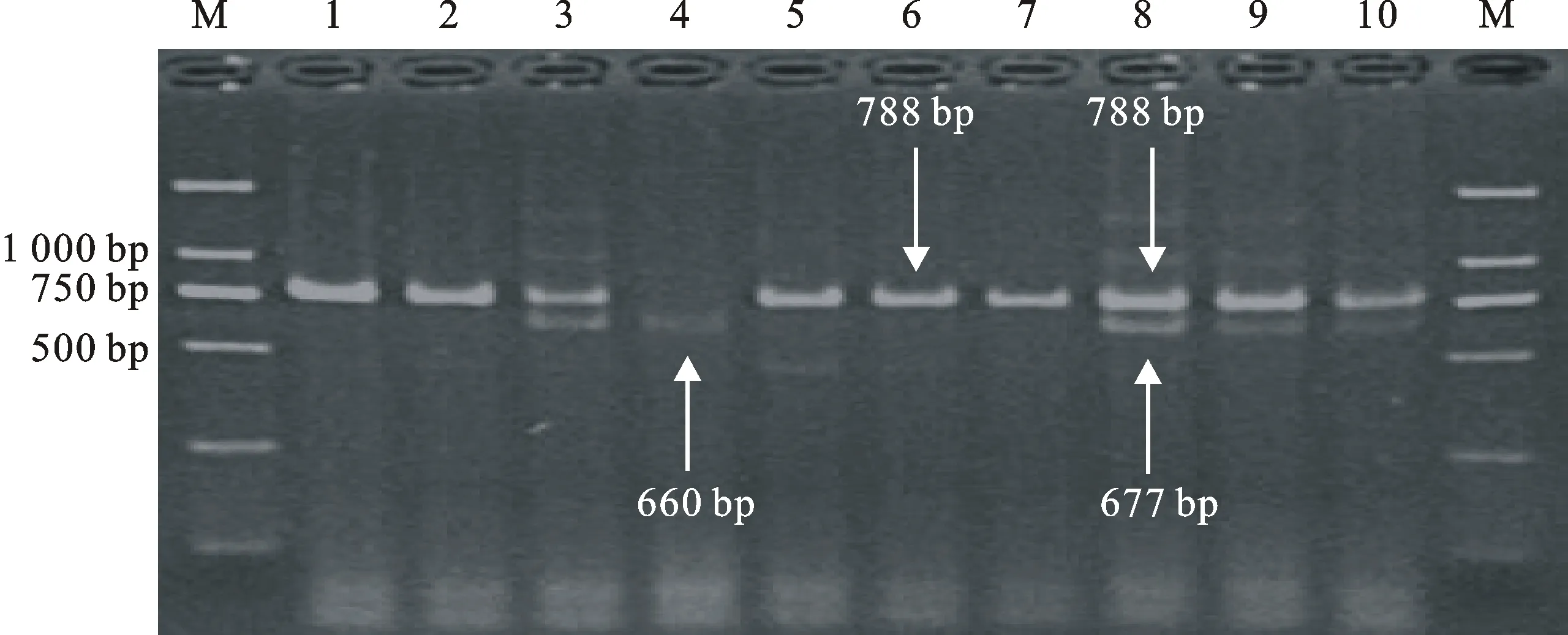
M:DL2000;1:新春14;2:红直头10;3:新春20;4:新春21;5:洛夫林18号;6:新春23;7:新春26;8:小白冬麦;9:新春28;10:新冬2号。
M:DL2000;1:Xinchun 14;2:Hongzhitou 10;3:Xinchun 20;4:Xinchun 21;5:Luofulin 18;6:Xinchun 23;7:Xinchun 26;8:Xiaobaidongmai;9:Xinchun 28;10:Xindong 2.
图2 Lox-B23标记检测部分小麦品种 TaLox-B2和 TaLox-B3位点的等位变异结果
Fig.2 Identification of TaLox-B2 and TaLox-B3 alleles by functional markers Lox-B23 in part of wheat varieties
表2 供试材料的 TaLox-B1、 TaLox-B2和 TaLox-B3 基因分布频率
Table 2 Frequency of TaLox-B1, TaLox-B2 and TaLox-B3 in tested materials
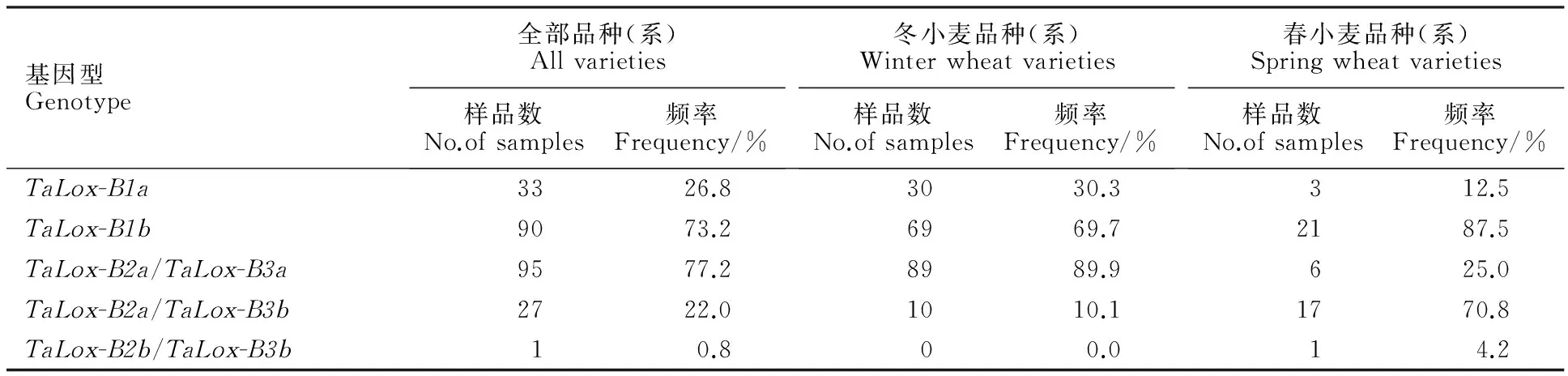
基因型Genotype全部品种(系)Allvarieties样品数No.ofsamples频率Frequency/%冬小麦品种(系)Winterwheatvarieties样品数No.ofsamples频率Frequency/%春小麦品种(系)Springwheatvarieties样品数No.ofsamples频率Frequency/%TaLox-B1a3326.83030.3312.5TaLox-B1b9073.26969.72187.5TaLox-B2a/TaLox-B3a9577.28989.9625.0TaLox-B2a/TaLox-B3b2722.01010.11770.8TaLox-B2b/TaLox-B3b10.800.014.2
2.2 不同类型新疆冬小麦材料的 TaLox-B1、 TaLox-B2和 TaLox-B3 等位变异类型及分布
对99份新疆冬小麦材料 TaLox-B1位点的检测结果表明,17份地方品种中具有 TaLox-B1a的材料有1份(5.9%),具有 TaLox-B1b的材料有16份(94.1%);39份引进品种中具有 TaLox-B1a的材料有19份(48.7%),具有 TaLox-B1b的材料有20份(51.3%);43份自育品种中具有 TaLox-B1a的材料有10份(23.3%),具有 TaLox-B1b的材料有33份(76.7%)。说明新疆冬小麦地方品种中优异等位变异 TaLox-B1a的分布频率远低于 TaLox-B1b,但在地方品种、自育品种和引进品种中逐次升高。 对于 TaLox-B3位点而言,17份地方品种均具有 TaLox-B3a;39份引进品种具有 TaLox-B3a的材料有37份(94.9%),2份(5.1%)具有 TaLox-B3b;43份自育品种中具有 TaLox-B3a的材料有35份(81.4%),具有 TaLox-B3b的材料有8份(18.6%)。总的来看,新疆冬小麦地方品种中优异等位变异 TaLox-B3a的分布频率远高于 TaLox-B3b,并在地方品种、引进品种和自育品种中逐次降低(表3)。
表3 不同类型冬小麦品种(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3 基因分布频率
Table 3 Frequency of TaLox-B1, TaLox-B1 and TaLox-B1 in different types of winter wheat varieties

基因型Genotype全部品种(系)Allvarieties样品数No.ofsamples频率Frequency/%地方品种(系)Localvarietieies样品数No.ofsamples频率Frequency/%引进品种(系)Introducedvarieties样品数No.ofsamples频率Frequency/%自育品种(系)Bredvarieties样品数No.ofsamples频率Frequency/%TaLox-B1a3030.315.91948.71023.3TaLox-B1b6969.71694.12051.33376.7TaLox-B2a/TaLox-B3a8989.917100.03794.93581.4TaLox-B2a/TaLox-B3b1010.1000.025.1818.6
2.3 不同时期新疆春小麦材料的 TaLox-B1、 TaLox-B2和 TaLox-B3等位变异类型及分布
24份新疆春小麦材料 TaLox-B1位点的检测结果表明,4份早期品种都具有 TaLox-B1b,20份晚期品种中有3份(15.0%)具有 TaLox-B1a,17份(85.0%)具有 TaLox-B1b;新疆春小麦早期品种未发现优异等位变异 TaLox-B1a,晚期品种虽然检测到 TaLox-B1a,但其分布频率也不高。对于 TaLox-B2位点而言,4份早期品种均具有 TaLox-B2a,20份晚期品种中19份(95.0%)材料具有 TaLox-B2a变异类型,只有1份(5.0%)材料具有 TaLox-B2b,表明新疆春小麦早期品种优异等位变异 TaLox-B2a的分布频率高于晚期品种。对于 TaLox-B3位点而言,4份早期品种均具有 TaLox-B3a,20份晚期品种中具有 TaLox-B3a和 TaLox-B3b的材料分别有6份(30.0%)和14份(70.0%);新疆春小麦材料早期品种均为优异等位变异 TaLox-B3a,且其分布频率远高于晚期品种(表4)。
2.4 新疆小麦材料的 TaLox-B1、 TaLox-B2和 TaLox-B3等位变异基因组合及分布频率
对123份新疆小麦材料 TaLox-B1、 TaLox-B2、 TaLox-B3等位变异类型的统计表明,具有最高LOX活性的基因型 TaLox-B1a/ TaLox-B2a/ TaLox-B3a或 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及具有中高LOX活性的基因型 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b的材料分别有95份(77.2%)和27份(22.0%);材料中未发现最低LOX活性的基因组合 TaLox-B1a/ TaLox-B2b/ TaLox-B3b,其中 TaLox-B1b/ TaLox-B2b/ TaLox-B3b是新发现的基因组合,在新疆材料中仅有1份(0.8%),其LOX活性尚未检测。而99份新疆冬小麦材料中,具有 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b基因组合的材料分别有89份(89.9%)和10份(10.1%)。24份新疆春小麦材料中,具有 TaLox-B1a/ TaLox-B2a/ TaLox-B3a和 TaLox-B1b/ TaLox-B2a/ TaLox-B3a以及 TaLox-B1b/ TaLox-B2a/ TaLox-B3b和 TaLox-B1a/ TaLox-B2a/ TaLox-B3b和 TaLox-B1b/ TaLox-B2b/ TaLox-B3b基因组合的材料分别有6份(25.0%)、17份(70.8%)和1份(4.2%)(表5)。总的来看,新疆冬小麦品种具有最高LOX活性的基因型频率远高于春小麦。8个冬小麦(石冬8号、新冬18号、新冬20号、新冬21号、新冬29号、新冬30号、新冬33号、新冬36号、伊农19、新冬7号)和2个春小麦自育品种(新春27号和新春29号)以及1个地方品种(库车白冬麦)具有最高LOX活性基因组合 TaLox-B1a/ TaLox-B2a/ TaLox-B3a,上述11个材料可作为选育携带高LOX活性基因的优异种质材料。
表4 不同类型春小麦品种(系)的 TaLox-B1、 TaLox-B2和 TaLox-B3基因的分布频率
Table 4 Frequency of TaLox-B1, TaLox-B2 and TaLox-B3 in different types of spring wheat varieties

基因型Genotype全部品种(系)Allvarieties样品数No.ofsamples频率Frequency/%早期品种(系)Earlyvarieties样品数No.ofsamples频率Frequency/%晚期品种(系)Latevarieties样品数No.ofsamples频率Frequency/%TaLox-B1a312.500315.0TaLox-B1b2187.54100.01785.0TaLox-B2a/TaLox-B3a625.04100.0630.0TaLox-B2a/TaLox-B3b1770.8001365.0TaLox-B2b/TaLox-B3b14.20015.0
表5 新疆小麦品种不同 TaLox-B等位基因的组合频率
Table 5 Frequency of Xinjiang wheat cultivars with different TaLox-B allelic combinations

等位变异组合Alleliccombination全部品种(系)Allvarieties样品数No.ofsamples频率Frequency/%冬小麦品种(系)Winterwheatvarieties样品数No.ofsamples频率Frequency/%春小麦品种(系)Springwheatvarieties样品数No.ofsamples频率Frequency/%TaLox-B1a/TaLox-B2a/TaLox-B3a2923.62727.328.3TaLox-B1b/TaLox-B2a/TaLox-B3a6653.76262.6416.7TaLox-B1b/TaLox-B2a/TaLox-B3b2419.588.11666.7TaLox-B1a/TaLox-B2a/TaLox-B3b32.422.014.2TaLox-B1b/TaLox-B2b/TaLox-B3b10.80014.2
3 讨 论
面粉及面制品颜色是小麦品质分级的重要指标[18-19]。长期以来我国消费者有喜欢高白度面食的习惯[20 ]。LOX通过与小麦面粉中的类胡萝卜素发生偶联氧化作用降低黄色素含量进而使小麦面粉变白[4],有望从根本上解决使用面粉增白剂对人体产生的潜在危害[21]。选育高LOX活性的小麦品种对提高面粉白度具有重要的作用。分子标记技术具有便捷、准确、稳定的特点,在小麦抗病性检测、品质分析以及与小麦各种农艺性状相关因素的检测方面发挥了巨大作用[22-23]。本研究所用的LOX活性相关功能标记LOX16、LOX18在Geng等[12]和相吉山等[14]研究中发现能有效应用于大群体小麦品种资源的检测以及LOX活性的判断,在育种实践中具有较高的应用价值;而Zhang等[16]在研究中也证明了Lox-B23的可靠性。利用LOX基因3个位点的功能标记能更有效地对高LOX活性材料进行选育。
相吉山等[14]利用 TaLox-B1位点的LOX16和LOX18功能标记检测195份新疆小麦资源,结果表明 TaLox-B1a基因的频率24.1%。许多研究利用LOX16、LOX18功能标记对不同区域小麦资源的检测结果均表明 TaLox-B1a基因型频率所占的比例较低[13,16,24-26]。本研究利用3对功能标记LOX16、LOX18和Lox-B23对123份新疆小麦资源进行检测,结果表明各位点等位变异类型的分布与前人研究结果基本一致。新疆冬小麦资源中的地方品种、自育品种和引进品种 TaLox-B1a基因型分布频率依次升高,其中地方品种(5.9%)和自育品种(23.3%)低于全国的分布频率(26.5%),但引进品种(48.7%)高于全国的分布频率,这可能与国内对LOX活性重视和研究起步较晚有关。自育品种 TaLox-B1a基因型分布频率高于地方品种但低于引进品种,这表明新疆小麦受地方品种的影响,使 TaLox-B1a基因型分布频率比较低,但随着引进品种不断引入新疆以及育种工作的进行,使高LOX活性 TaLox-B1a基因型频率升高,但新疆冬小麦 TaLox-B1a基因型分布频率总体水平仍然较低(30.3%),与国外品种存在差距。同时也表明,引种是小麦品质遗传改良的有效途径之一,应加大对国外优良品种的引进和自育品种的选育,进而缩小与国外优良品种的差距。
Zhang等[16]利用Lox-B23标记检测136份中国小麦资源,结果表明, TaLox-B1a/ TaLox-B2a/ TaLox-B3a或 TaLox-B1b/ TaLox-B2a/ TaLox- B3a基因组合的频率为62.5%。本研究利用Lox-B23功能标记对123份新疆小麦资源进行检测,结果最高LOX活性的基因组合频率为77.2%,高于Zhang等[16]的研究结果。在新疆冬小麦品种中最高LOX活性的基因组合频率高达89.9%,而在新疆春小麦品种中只有25.0%。虽然新疆小麦品种最高LOX活性的基因组合频率受春小麦品种的影响有所降低,但仍然高于全国水平(62.5%);新疆冬小麦品种最高LOX活性的基因组合频率高于全国水平,同时高于新疆总体水平,说明新疆冬小麦品种资源是培育高LOX活性品种的优异基因源。本研究还检测出新春21号品种的基因型是 TaLox-B1b/ TaLox-B2b/ TaLox-B3b,虽其LOX活性尚未检测,但其 TaLox-B1、 TaLox-B2、 TaLox-B3位点基因都与高LOX活性负相关,且低LOX活性材料具有更耐储藏的特性[1,16],因此新春21号小麦品种可能具有较长的储藏期。
本研究将3个LOX主效基因位点结合起来进行选择,很大程度上弥补了单一位点选择的片面和不足,将能更有效的选出高LOX活性材料。目前,对新疆小麦LOX活性的研究都集中在分子标记检测上,并用基因型来选择高LOX活性材料,还没有对新疆小麦材料LOX活性进行测定研究。同时,本研究所用春小麦样本数有限,均为自育品种,分类时未能与冬小麦材料采用同一标准,致使春小麦是按不同时期进行分类,虽然也能有效反映新疆小麦材料中不同等位变异类型分布的演化规律及其成因,但未能反映冬春麦之间演化规律的异同。下一步我们的工作是进一步扩大新疆春小麦材料数量,开展新疆小麦材料LOX活性的表型测定,并通过表型结合基因型对新疆小麦材料的LOX活性进行综合评价,这将提升高LOX活性选育效率,为小麦品质改良提供优异基因源和基础材料。
[1] 耿洪伟.小麦脂肪氧化酶(LOX)活性QTL定位与功能标记开发[D].乌鲁木齐:新疆农业大学,2010:9.
GENG H W.Mapping QTLs for lipoxygenase activity and development of functional markers for LOX gene in common wheat [D].Urumqi:Xinjiang Agricultural University,2010:9.
[2] 胡瑞波,田纪春.小麦主要品质性状与面粉色泽的关系[J].麦类作物学报,2006,26(3):96.
HU R B,TIAN J C.Relationship between main quality characteristics and wheat flour color [J].JournalofTriticeaeCrops,2006,26(3):96.
[3] 汪 帆,郑文寅,黄建华.20个小麦品种(系)籽粒LOX活性和类胡萝卜素含量及全麦粉色泽的研究[J].麦类作物学报,2012,32(1):68.
WANG F,ZHENG W Y,HUANG J H,etal.Lipoxygenase activity,carotenoids content and wholemill colors in grains of twenty wheat varieties [J].JournalofTriticeaeCrops,2012,32 (1):68.
[4] 郑文寅,汪 帆,司红起.普通小麦籽粒LOX、PPO活性和类胡萝卜素含量变异及对全麦粉色泽的影响[J].中国农业科学,2013,46(6):1087.
ZHENG W Y,WANG F,SI H Q,etal.Variations of LOX and PPO activities and carotenoid conten as well as their influence on whole flour color in common wheat [J].ScientiaAgricultureSinica,2003,46(4):1087.
[5] SHEWRY P R,TATHAM A S,BARRO F,etal.Biotechnology of breadmaking-unravelling and manipulating the multi-protein gluten complex [J].Biotechnology,1995,13(11):1185.
[6] BORRELLI G M,TROCCOLI A,FONZO N,etal.Durum wheat lipoxygenase activity and other quality parameters that affect pasta color [J].CerealChemistry,1999,76(3):335.
[7] 王 慧,郑文寅,樊 宏,等.不同小麦品种籽粒中LOX活性及基因型和环境互作分析[J].中国粮油学报,2011,26(1):11.
WANG H,ZHENG W Y,FAN H,etal.Lipoxygenase activity and its genotype and environment interactions for different wheat varieties [J].JournaloftheChineseCerealsandOilsAssociation,2011,26(1):11.
[8] PSHENICHNIKOVA T A,OSIPOVA S V,PERMYAKOVA M D,etal.Mapping of quantitative trait loci (QTL) associated with activity of disulfide reductase and lipoxygenase in grain of bread wheatTriticumaestivumL. [J].RussianJournalofGenetics,2008,44(5):567.
[9] FENG B,DONG Z Y,XU Z B,etal.Molecular analysis of lipoxygenase (LOX) genes in common wheat and phylogenetic investigation of LOX proteins from model and crop plants [J].JournalofCerealScience,2010,52(3):387.
[10] HART G E,LANGSTON P J.Chromosomal location and evolution of isozyme structural genes in hexaploid wheat [J].Heredity,1977,39(2):263.
[11] GENG H W,ZHANG Y,HE Z H,etal.Molecular markers for tracking variation in lipoxygenase activity in wheat breeding [J].MolecularBreeding,2011,28(1):117.
[12] GENG H W,XIA X C,ZHANG L P,etal.Development of functional markers for a lipoxygenase gene TaLox-B1 on chromosome 4BS in common wheat [J].CropScience,2012,52(2):568.
[13] 张钰玉,王晓龙,张晓科,等.陕西小麦品种(系)脂肪氧化酶活性基因的遗传多态性分析[J].麦类作物学报,2012,32(4):616.
ZHANG Y Y,WANG X L,ZHANG X K,etal.Genetic polymorphism of lipoxygenase genes in Shaanxi wheat cultivars and lines [J].JournalofTriticeaeCrops,2012,32(4):616.
[14] 相吉山,穆培源,桑 伟,等.新疆小麦品种资源脂肪氧化酶活性基因 TaLox-B1的分布特征研究[J].麦类作物学报,2013,33(2):279.
XIANG J S,MU P Y,SANG W,etal.Distribution characteristics of lipoxygenase gene TaLox-B1 in wheat varieties from Xinjiang [J].JournalofTriticeaeCrops,2013,33(2):279.
[15] 杨淑萍,张宏纪,刘文林,等.黑龙江春小麦脂肪氧化酶活性基因多态性分析[J].核农学报,2014,28(11):1972.
YANG S P,ZHANG H J,LIU W L,etal.Polymorphism of lipoxygenase genes in Heilongjiang spring wheat cultivars[J].JournalofNuclearAgriculturalSciences,2014,28(11):1972.
[16] ZHANG F Y,CHEN F,WU P,etal.Molecular characterizeation of lipoxygenase genes on chromosome 4BS in Chinese bread wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2015,128(8):1467.
[17] LAGUDAH E S,APPLES R,MCNEIL D.The Nor-D3 locus ofTriticumtauschii:natural variation and genetic linkage to markers in chromosome 5 [J].Genome,1991,34(3):387.
[18] 何中虎,晏月明,庄巧生,等.中国小麦品种品质评价体系建立与分子改良技术研究[J].中国农业科学,2006,39(6):1091.
HE Z H,YAN Y M,ZHUANG Q S,etal.Establishment of quality evaluation system and utilization of molecular methods for the improvement of Chinese wheat quality [J].ScientiaAgricultureSinica,2006,39(6):1091.
[19] 胡瑞波,田纪春,邓志英,等.中国白盐面条色泽影响因素的研究[J].作物学报,2006,32(9):1338.
HU R B,TIAN J C,DENG Z Y,etal.Factors related to Chinese white salted noodle color [J].ActaAgronomicaSinica,2006,32(9):1338.
[20] 张 晓,田纪春.若干高白度小麦的色泽优势及形成因素分析[J].中国农业科学,2008,41(2):347.
ZHANG X,TIAN J C.The color advantage of Chinese wheat with high whiteness and analysis of factors affecting color formation [J].ScientiaAgriculturaSinica,2008,41(2):347.
[21] 田纪春.禁用面粉增白剂后保证中国传统面食品色泽品质的对策[J].麦类作物学报,2007,27(2):364.
TIAN J C.Strategy of keeping quality of wheat flour and food color when the flour bleacher inhibited [J].JournalofTriticeaeCrops,2007,27(2):364.
[22] 武玉华.分子标记辅助选择在小麦抗病和品质遗传育种中的应用[J].麦类作物学报,2007,27(5):928.
WU Y H.Application of marker-assisted selection in disease-resistance and high-quality wheat breeding [J].JournalofTriticeaeCrops,2007,27(5):928.
[23] 王长有,吉万全,薛秀庄.分子标记技术在小麦遗传育种中的应用现状[J].麦类作物学报,2000,20(4):75.
WANG C Y,JI W Q,XUE X Z.The application of molecular marker techniques in the wheat genetic and breeding research [J].JournalofTriticeaeCrops,2000,20(4):75.
[24] 吴培培,宋 双,张福彦,等.黄淮麦区部分小麦种质脂肪氧化酶活性分析及等位基因检测[J].中国农业科学,2015,48(2):207.
WU P P,SONG S,ZHANG F Y,etal.The allelic variation of lipoxygenase genes in bread wheat cultivars from the Yellow and Huai wheat areas of China [J].ScientiaAgricultureSinica,2015,48(2):207.
[25] 杨淑萍,张宏纪,刘文林,等.黑龙江春小麦脂肪氧化酶活性基因多态性分析[J].核农学报,2014(11):1972.
YANG S P,ZHANG H J,LIU W L,etal.Polymorphism of lipoxygenase genes in Heilongjiang spring wheat cultivars [J].JournalofNuclearAgriculturalScience,2014,28(11):1972.
[26] 杨 杰,亢 玲,王宪国,等.宁夏小麦脂肪氧化酶基因的检测与分布[J].麦类作物学报,2015,35(7):896.
YANG J,KANG L,WANG X G,etal.Detections and analysis of lipoxygenase genes in Ningxia wheat varieties [J].JournalofTriticeaeCrops,2015,35(7):896.
Molecular Identification and Distribution of TaLox-B1, TaLox-B2 and TaLox-B3 Loci in Xinjiang Wheat Cultivars
BAI Lu1,YU Feixiong2,YASENG Ruxianguli3,LÜ Wenjuan2,REN Yi2,XIE Lei2,GENG Hongwei2
(1.College of Science and Technology,Xinjiang Agricultural University,Urumqi,Xinjiang 830052,China; 2.Key Laboratory of Agricultural Biological Technology/College of Agronomy,Xinjiang Agricultural University,Urumqi,Xinjiang 830052,China;3.Bayingol Mongolian Autonomous Prefecture of Agriculture,Korla,Xinjiang 841000,China)
The color of the wheat flour is an important index of wheat quality traits. This study used the functional markers LOX16,LOX18 and Lox-B23,based on the development of wheat 4BS chromosome,and molecular markers were detected by TaLox-B1, TaLox-B2 and TaLox-B3 in 123 wheat varieties in Xinjiang. The results showed that,there were 33(26.8%)varieties with TaLox-B1a allele exhibited high Lox activity,90(73.2%)varieties with TaLox-B1b allele showed low Lox activity; 122(99.2%)varieties with TaLox-B2a allele exhibited high Lox activity,only 1(0.8%)varieties with TaLox-B2b allele showed low Lox activity; 95(77.2%)varieties with TaLox-B2a allele exhibited high LOX activity,28(22.8%)varieties with TaLox-B2b allele showed low Lox activity. In different types of winter wheat varieties,the frequency distribution of TaLox-B1a was in order of introduced varieties>self-cultivated varieties >local varieties. In different types of spring wheat varieties,the TaLox-B1a exists only in the late varieties,and the distribution frequency of both TaLox-B2a and TaLox-B3a was in order of early varieties (100%) >late varieties.The analysis of 3 loci genotype combinations indicated that there are 5 kinds of genetic combinations in Xinjiang wheat varieties,which were both TaLox-B1a/ TaLox-B2a/ TaLox-B3a and TaLox-B1b/ TaLox-B2a/ TaLox-B3a combinations exhibited the highest LOX activity,and were both TaLox-B1b/ TaLox-B2a/ TaLox-B3b and TaLox-B1a/ TaLox-B2a/ TaLox-B3b combinations exhibited high LOX activity,and TaLox-B1b/ TaLox-B2b/ TaLox-B3b showed the lowest LOX activity,and the distribution frequency was 23.6%,53.7%,19.5%,2.4% and 0.8%,respectively. The genotype frequency of high LOX activity in wheat cultivars in Xinjiang was higher than the national average. In this study,3 loci functional markers were combined more effectively applied to wheat quality molecular marker assisted selection.
Xinjiang wheat; Lipoxygenase; Function marker; Variation combination
时间:2016-11-04
2016-05-16
2016-06-05
国家自然科学基金项目(31260327);新疆自治区级大学生创新项目(20150016);2014年度留学人员科技活动择优资助项目(耿洪伟)
E-mail:bailu32@163.com
耿洪伟(E-mail:hw-geng@163.com)
S512.1;S330
A
1009-1041(2016)11-1456-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20161104.0924.014.html
猜你喜欢
作物学报(2022年6期)2022-04-08
上海金属(2021年6期)2021-12-02
国际医学放射学杂志(2021年5期)2021-10-22
昆明医科大学学报(2021年3期)2021-07-22
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
生物学通报(2019年3期)2019-02-17
电脑知识与技术(2018年19期)2018-11-01
麦类作物学报(2018年4期)2018-05-11
第一财经(2017年36期)2017-09-25
