
辣椒雄性不育系及其保持系材料花蕾生理生化特性研究
2017-05-18 17:27侯杰韩玉珠李娇张晓旭李爽
长江蔬菜·学术版 2017年4期
侯杰++韩玉珠+李娇++张晓旭++李爽++杨美军++祝聪宇+王凯++张广臣

辣椒是常异花授粉植物,其杂交产生的一代杂种具有品质好、抗性强、产量高等显著优势,因此,辣椒杂交育种成为获得优良品种的普遍应用方法[1]。目前辣椒杂交制种包括人工去雄杂交制种和雄性不育杂交制种,传统的人工去雄不仅费时费工,而且影响坐果率和果实质量,致使制种成本高、产量不稳、杂交种纯度难以保证。而利用雄性不育系生产种子可以省去人工去雄的过程,是简化制种手续、节约生产成本和提高种子纯度的有效方法[2]。自Martin等[3]和Peterson[4]报道辣椒細胞核雄性不育和质核互作雄性不育以来,如何在辣椒产业发展中研究利用雄性不育性状受到育种家们的高度重视。植物的生长发育涉及到酶、营养物质、生长物质等各类成分,只有各方面协同作用才能正常生长发育,其中任一环节出现问题都会导致性状变异[5]。花粉的发育与雄蕊尤其是花药密切相关,若在花药发育的过程中遇到激素、酶的异常调控、营养物质缺乏、能量代谢异常等现象,都会影响花的育性[6]。孙立全等[7]和刘金兵等[8]及一些国内学者研究发现,辣椒不育材料和可育材料的过氧化物酶 (POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)的活性以及可溶性糖、可溶性蛋白和游离脯氨酸的含量变化会影响花的育性。本试验通过对辣椒雄性不育材料O-6、O-9和其保持系小孢子各个发育时期部分生化指标的测定分析,揭示辣椒雄性不育材料O-6、O-9小孢子的败育机理,以期为今后的研究提供理论依据。
1 材料与方法
1.1 试验材料
材料为羊角椒不育系O-6、O-9及其保持系,由吉林农业大学辣椒课题组提供,待在其盛花期时按形态特征分为5个等级[9](表1),采其花蕾进行试验测定。
1.2 试验方法
于盛花期采摘分级后的花蕾,测定生理生化指标。可溶性糖含量采用蒽酮法测定,可溶性蛋白含量采用G-250考马斯亮蓝法测定,游离脯氨酸含量采用酸性茚三酮比色法测定,POD活性采用愈创木酚法测定,SOD活性采用氮蓝四唑法测定,丙二醛含量采用TBA法测定[10]。
1.3 数据处理方法
试验数据采用Microsoft Excel和DPS软件进行处理和统计分析。
2 结果与分析
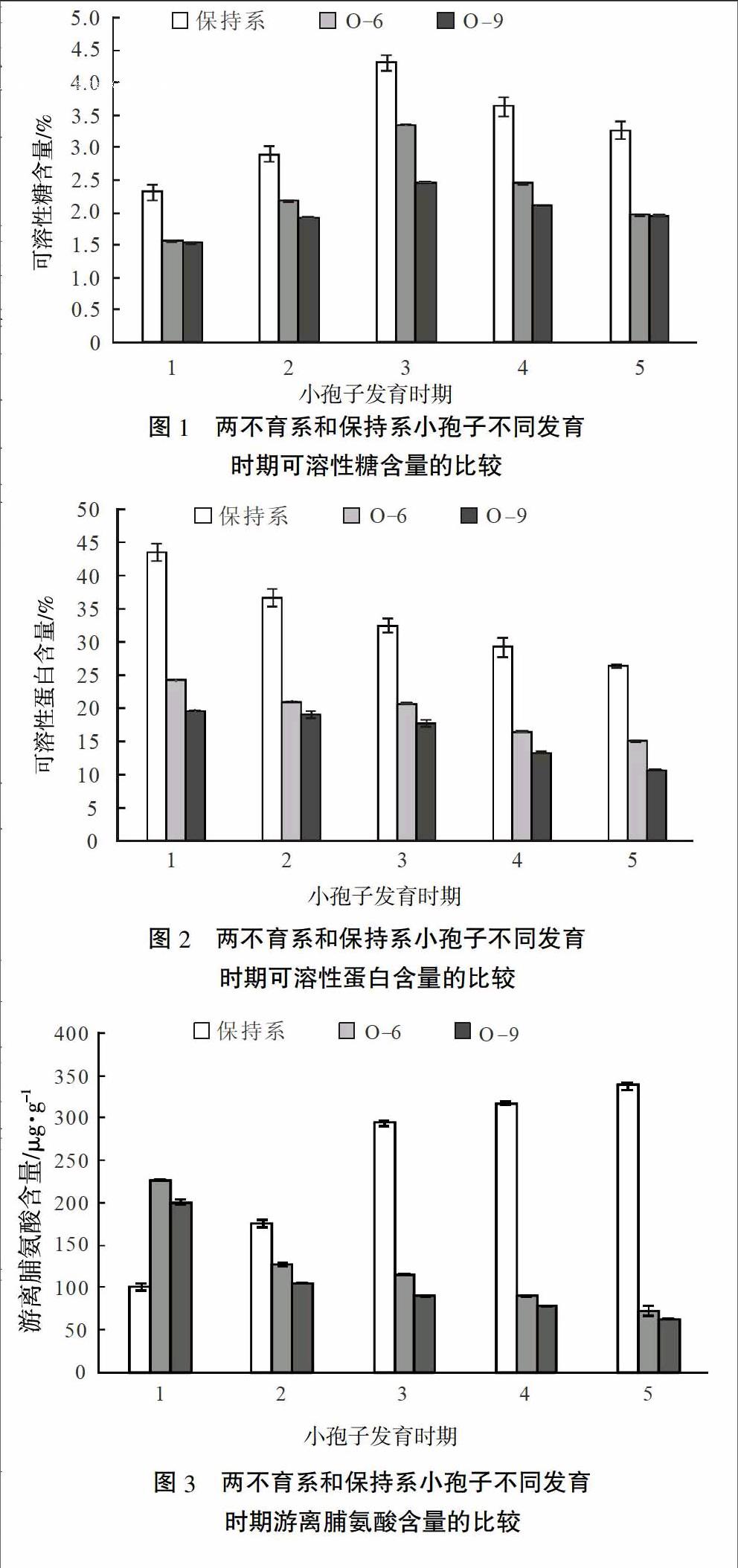
2.1 小孢子发育各时期可溶性糖含量
辣椒小孢子不同发育时期花蕾中可溶性糖含量的测定结果表明(图1),在花蕾发育的各个时期,保持系可溶性糖含量均高于不育系,与可溶性蛋白含量变化趋势相似。但随着花蕾的发育,可溶性糖含量在两系中均呈先上升后下降趋势,且在四分体时期最高。不育材料O-6与不育材料O-9的可溶性糖含量变化趋势相同,但O-9变化幅度较小。
2.2 小孢子发育各时期可溶性蛋白含量
辣椒小孢子不同发育时期花蕾中可溶性蛋白含量显示(图2),不育材料在各时期花蕾中的可溶性蛋白含量明显低于其保持系。随着花蕾的生长发育,各材料花蕾中可溶性蛋白含量均呈下降趋势,其中可溶性蛋白在造孢细胞时期含量最高,在成熟花粉粒时期含量最低。不育材料O-6各时期可溶性蛋白含量明显高于O-9。
2.3 小孢子发育各时期游离脯氨酸含量
辣椒小孢子不同发育时期花蕾中脯氨酸含量的测定结果表明(图3),在造孢细胞时期不育材料花蕾的游离脯氨酸含量明显高于保持系,但随着花蕾的生长发育,不育材料游离脯氨酸含量呈下降趋势,而保持系花蕾中游离脯氨酸含量则迅速积累,含量明显高于不育系,在花粉粒成熟期达到最大值。虽然2种不育材料游离脯氨酸含量均呈下降趋势,但O-6各时期花蕾中游离脯氨酸含量明显高于O-9。
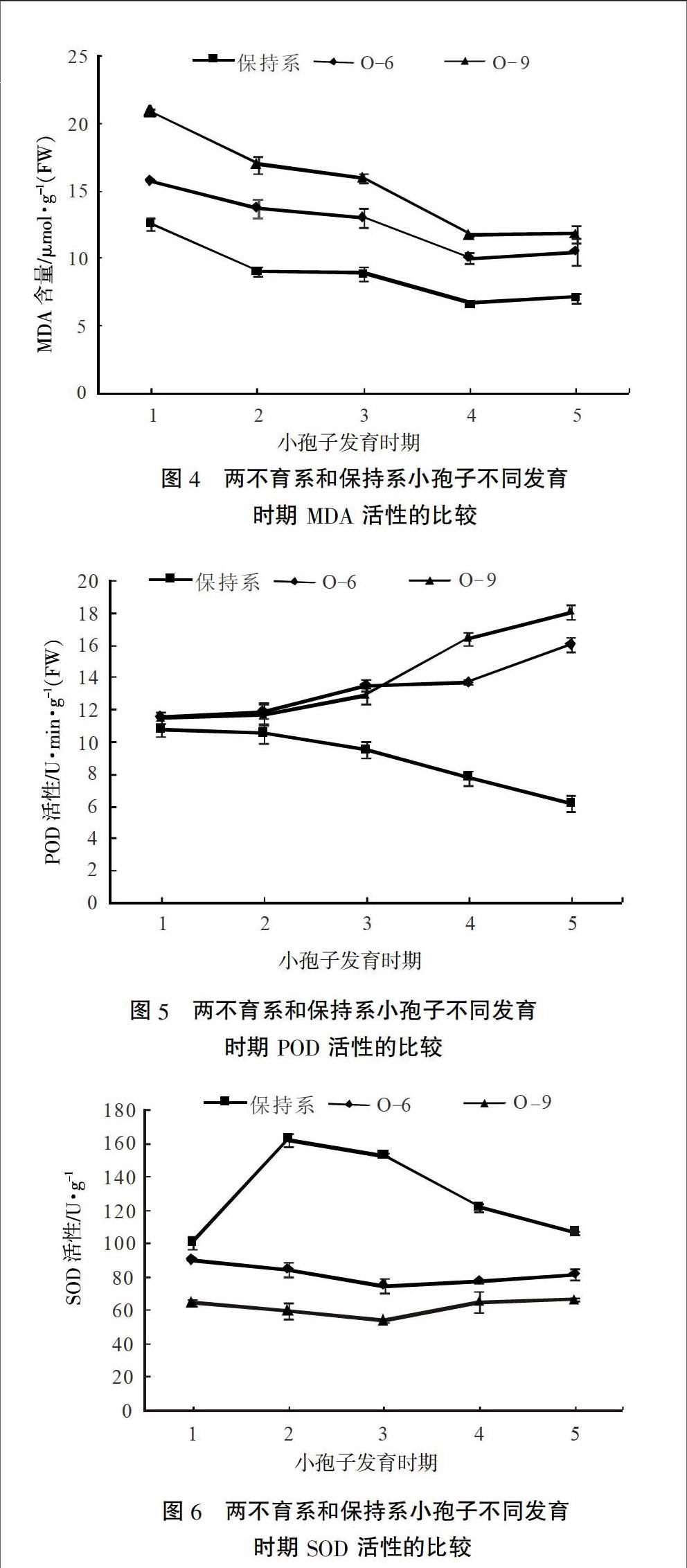
2.4 小孢子发育各时期MDA含量
辣椒小孢子不同发育时期花蕾中丙二醛含量的测定结果表明(图4),随着花蕾的发育,不育材料中丙二醛含量明显高于其保持系,且丙二醛含量均呈下降趋势,但在花粉粒成熟期含量又有所升高。不育材料O-9的丙二醛含量明显高于不育材料O-6。
2.5 小孢子发育各时期POD活性
辣椒小孢子不同发育时期花蕾中过氧化物酶活性的测定结果表明(图5),在造孢细胞时期不育材料与其保持系花药中过氧化物酶活性很接近,但随着花蕾的发育,不育材料花蕾中过氧化物酶活性呈上升趋势,而其保持系花蕾中的过氧化物酶活性呈下降趋势。从四分体时期开始,不育材料的过氧化物酶活性明显高于其保持系,在花粉粒成熟期达到最大值,但不育材料O-6在单核小孢子时期的POD活性稍有减弱。
2.6 小孢子发育各时期SOD活性
辣椒小孢子不同发育时期花蕾中超氧化物歧化酶活性的测定结果表明(图6),保持系各时期花蕾中超氧化物歧化酶活性均高于不育系。随着花蕾的发育,保持系超氧化物歧化酶活性呈先上升后下降趋势,但不育系超氧化物歧化酶活性呈先下降后上升趋势,且在四分体时期时期发生转折。不育材料O-6超氧化物歧化酶活性变化趋势虽与O-9相同,但花蕾各时期SOD活性均高于O-9。
3 结论与讨论
在小孢子发育的各个时期,不育系花药中可溶性糖含量低于保持系,与李莹莹等[11]的研究结果一致。刘金兵等[12]则证明,可溶性糖含量无论是叶片还是花药中,均为保持系高于不育系。花药中多糖对小孢子发育极为重要,它既是细胞的组成部分,也是小孢子发生过程中积累的营养物质,还作为其发育过程中的信号物质,碳水化合物与材料不育有直接联系。邓明华等[13]研究发现,不育系辣椒小花蕾时期的可溶性蛋白含量比中花蕾、大花蕾的含量高,但保持系花蕾中可溶性蛋白含量持续升高。但本试验不育材料在小孢子发育各时期花蕾中的可溶性蛋白含量明显低于其保持系。随着花蕾的生长发育,各材料花蕾中可溶性蛋白含量均呈下降趋势,测定结果与逯红栋等[14]的研究结果一致。蛋白质是细胞的重要组成部分,在花药和花粉发育过程中也起着非常关键性的作用,不仅为花药和花粉发育提供营养物质,而且还作为信号物质影响花药和花粉的发育[15],不育材料可溶性蛋白含量比其保持系低,说明不育材料的不育性可能是由于可溶性蛋白的缺乏造成。
本试验结果表明,辣椒小孢子不同发育时期,造孢细胞时期不育材料花蕾中的游离脯氨酸含量明显高于保持系,但随着花蕾的生长发育,不育材料游离脯氨酸含量呈下降趋势,而保持系花蕾中游离脯氨酸含量则迅速积累,明显高于不育系,在花粉粒成熟期达到最大值,与逯红栋等[14]、刘金兵等[12]的结论一致。前人研究表明,游离脯氨酸是影响小孢子正常发育的关键因素,能够为花粉萌发和花粉管的伸长提供必需的能量及氮源,还能直接参与合成蛋白质,具有重要的生理功能[16]。Palif[17]认为,花药内游离脯氨酸含量与育性呈相关,并提出可作为花粉的育性指标。本试验不育材料中游离脯氨酸含量的减少可能是致使其不育的原因。
丙二醛(MDA)是自由基作用于脂质发生过氧化反应的终产物,MDA含量在一定程度上反映了膜脂受破坏的程度。李莹莹等[11]研究认为,在不育系中,随着花蕾发育MDA含量呈下降趋势。但本试验结果显示,不育材料小孢子发育各时期的MDA含量都明显高于其保持系,与安岩等[18]在胡萝卜(Daucus carota)上的研究结果一致。随着花蕾发育,MDA含量在不育材料和其保持系中均呈下降趋势,但在花粉粒成熟期又有所升高,说明MDA的含量可能还与其他因素有关,与彭婧等[19]的结论
一致。
辣椒小孢子在花蕾发育的各个时期,保持系各时期花蕾中SOD活性均高于不育系。随着花蕾的发育,保持系SOD活性呈先上升后下降趋势,但不育系SOD活性呈先下降后上升趋势,且在四分体时期时期发生转折,与李莹莹等[20]研究结论相同。
在小孢子发育的各个時期,不育材料过氧化物酶(POD)活性大于可育材料,与邓明华等[5]发现在花药发育的任何时期不育系POD活性均高于可育系结果一致,但李莹莹等[11]得出相反的结果。本试验在造孢细胞时期,不育材料与其保持系花药中过氧化物酶活性接近,但随着花蕾的发育,不育材料花蕾中POD活性呈上升趋势,而其保持系花蕾中过氧化物酶活性呈下降趋势。POD是一种复杂成分的酶,具有多种生理功能。许多研究表明,过氧化物酶(POD)与内源激素代谢息息相关,其活性与植物体内IAA呈负相关,若POD活性增强,IAA含量就会降低,导致新陈代谢紊乱,营养物质积累停止,造成小孢子败育[21]。本试验结果显示,不育材料花蕾在小孢子四分体期POD活性明显高于保持系,推测酶类活性的增强致使花粉粒内新陈代谢紊乱从而造成雄性不育,对这一结论有待进一步探讨。
参考文献
[1] 杨泽良,胡开林,党选民,等.基因工程雄性不育及辣椒杂种优势利用[J].辣椒杂志,2006(1):9-12.
[2] 黄炜.辣椒雄性不育系两用系创制及其不育机理研究[D].杨凌:西北农林科技大学,2012.
[3] Martin J A, Crawford J H. Several types of sterility in Capsicum frutescence[J]. J Am Soc Hortic Sci, 1951, 57: 335-338.
[4] Peterson P A. Cytoplasmic inherited male sterility in Capsicum annuum L.[J]. Am Nat, 1958, 92: 111-119.
[5] 邓明华.辣椒胞质雄性不育的分子生理机制及亲缘关系
研究[D].长沙:中南大学,2011.
[6] 黄厚哲.植物生长素亏素与雄性不育的发生[J].厦门大学学报,1994,23(1):82-96.
[7] 孙立全,霍治军,常彩涛,等.辣椒雄性不育系小孢子发育及脯氨酸等含量的研究[J].华北农学报,2003,18(4):39-41.
[8] 刘金兵,侯喜林,王述彬,等.甜椒胞质雄性不育(CMS)系及其保持系花药中游离氨基酸含量[J].江苏农业学报,2006,22(1):68-70
[9] 李国琴.辣椒雄性不育花器官败育特征的研究及辣椒
CaCP26的cDNA的克隆和分析[D].杨凌:西北农林科技大学,2011.
[10] 张治安,陈展宇.植物生理生化试验技术[M].长春:吉林大学出版社,2008.
[11] 李莹莹,魏佑营,张瑞华,等.辣椒雄性不育小孢子发育过程中物质代谢研究[J].西北农业学报,2006,15(3):134-137.
[12] 刘金兵,侯喜林,陈晓峰,等.甜椒胞质雄性不育系及其保持系生化特性研究[J].园艺学报,2006,33(3):629-633.
[13] 邓明华,邹学校,周群忻.辣椒细胞质雄性不育系与保持系生化特性研究[J].湖南农业大学学报:自然科学版,2002,28(6):492-494.
[14] 逯红栋,巩振辉,黄炜,等.9个辣椒雄性不育材料花蕾生理生化特性研究[J].西北植物学报,2006(26):832-835.
[15] 韩璐,王晶,郑蕊.蛋白质组学在植物雄性不育研究中的应用[J].北方园艺,2011(18):206-208.
[16] 朱广廉,孙超,曹宗巽.太谷核不育小麦可育花药内游离脯氨酸的来源、利用及与不育花药败育的关系[J].植物生理学报,1985,11(2):122-129.
[17] Palif G. The profine content of fertility of the pollen inbreed maize lines [J]. Acta Bet Acad Sci, 1981, 27(2): 179-187.
[18] 安岩,沈火林,乔志霞.胡萝卜雄性不育系和保持系生理生化特性的比较分析[J].中国瓜菜,2008(1):3-7.
[19] 彭婧,巩振辉,黄炜,等.辣椒雄性不育材料H9A小孢子败育机理[J].植物学报,2010,45(1):44-51.
[20] 李莹莹,魏佑营,张瑞华,等.辣椒雄性不育“三系”花蕾中3种同工酶活性的动态变化[J].中国农学通报,2005,21(4):226-229.
[21] 邓明华.辣椒胞质雄性不育株生理生化特性及离体培养研究[D].长沙:湖南农业大学学报,2003:8-14.
猜你喜欢
河南科技(2022年1期)2022-03-11
甘肃农业科技(2021年12期)2021-01-13
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
江苏农业科学(2017年19期)2017-11-22
江苏农业科学(2017年19期)2017-11-22
农家顾问(2016年7期)2016-05-14
江苏农业科学(2015年5期)2015-10-20
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年3期)2015-07-07
