
蝴蝶兰Phalaenopsis‘Frigdaas Oxford’和Phal.SH49正反交后代观赏性状遗传倾向研究
2017-05-30 10:48李佐肖文芳陈和明刘金维吕复兵
热带作物学报 2017年1期
关键词:蝴蝶兰
李佐 肖文芳 陈和明 刘金维 吕复兵

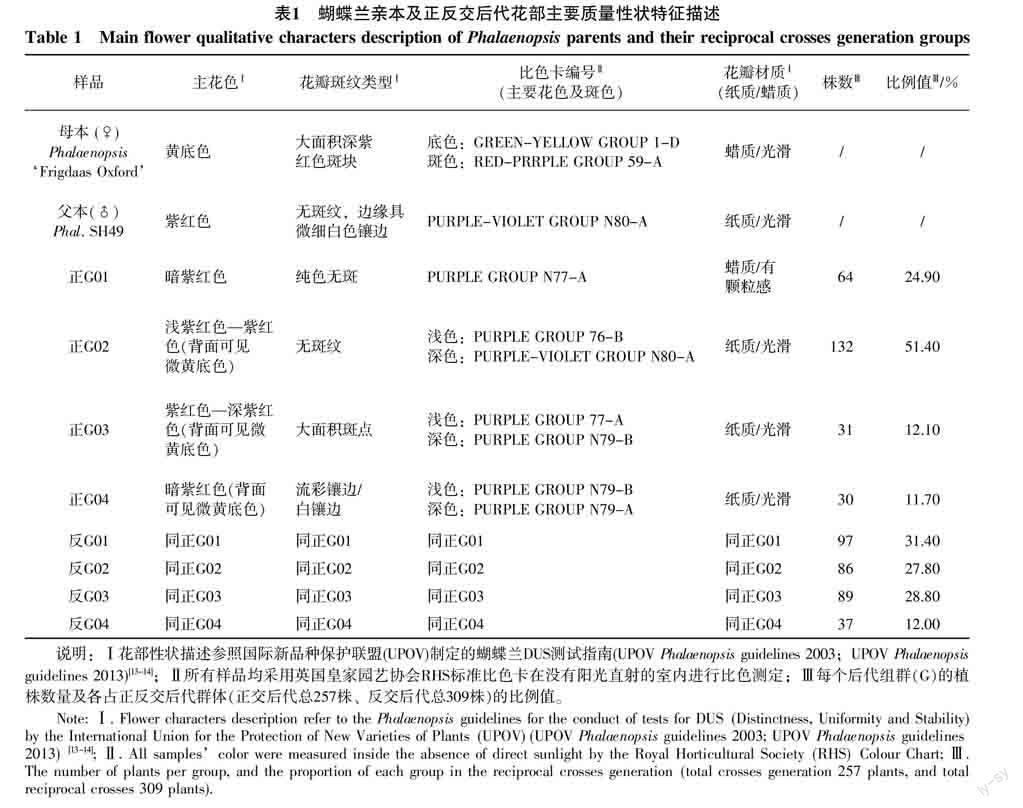
摘 要 以花色、花斑类型、花朵质地、株型大小、花朵大小等为蝴蝶兰杂交育种的目标性状,对Phalaenopsis ‘Frigdaas Oxford和Phal. SH49及其正反交后代群体的观赏性状进行分析,研究育种目标性状在后代中的表现与分离。结果表明:正反交F1代的分离规律和变化趋势基本一致,根据主要花部性状特点分为4个组群(G01~G04),杂交F1代在9个数量性状的平均值表现均较中亲值明显降低,适合选育小株型或小花型后代个体;杂交F1代在花底色、花斑类型、花朵材质等质量性状上变化丰富,不同花色的遗传能力与覆盖能力有所差别,存在较深花斑色覆盖遮挡住较浅花底色的花色表现,且出现了亲本没有的花斑变化,这些性状的分离特征和遗传倾向将给今后新个体或新单株的目标性状预测筛选提供更多参考和依据。
关键词 蝴蝶兰;正反交F1代;观赏性状;性状分离
中图分类号 S682.31 文献标识码 A
Genetic Tendency of Ornamental Characters in Reciprocal Crosses
Generation Between Phalaenopsis‘Frigdaas Oxford
and Phal. SH49 Cultivars
LI Zuo1,2, XIAO Wenfang1,2, CHEN Heming1,2, LIU Jinwei1, Lü Fubing1,2 *
1 Environmental Horticulture Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong 510640, China
2 Guangdong Provincial Key Laboratory of Ornamental Plant Germplasm Innovation and Utilization, Guangzhou, Guangdong 510640, China
Abstract Phalaenopsis‘Frigdaas Oxford, Phal. SH49 cultivars and the F1 offspring of reciprocal crossing were used to analyze the genetic tendency of the ornamental characters, such as flower color, flower pattern, flower texture, plant size and flower size, etc. The results showed as follow that the separation tendency was basically the same in both the cross and reciprocal cross generation. The F1 offspring were divided into 4 groups(G01-G04)based on the flower characteristics. The mean values of the 9 quantitative characters in the F1 offspring were significantly lower than the midparent value, which were suitable for the breeding of small plant size or small flower type offspring. The qualitative characters of the F1 offspring showed various separation on flower ground color, flower pattern and flower texture, etc. The heritability and the coverage degree differed in different color, that the darker color pattern could cover the lighter ground color, and furthermore there were new color pattern appears. These characteristics separation and genetic tendency could support in selecting new individuals or new cultivars in the future, and this research could supply directed breeding reference for certain floral traits.
Key words Phalaenopsis; reciprocal crosses generation; ornamental characters; genetic separation
doi 10.3969/j.issn.1000-2561.2017.01.002
蝴蝶兰(Phalaenopsis)在丰富的兰科资源中,因其极高的观赏与经济价值成为目前国内及国际最流行的盆栽花卉品種之一。选育具有更强竞争力的新品种,提高市场的占有率,满足不同消费人群和消费市场的需求,是蝴蝶兰产业发展的必然方向[1-3]。如何提高蝴蝶兰杂交育种的效率、增强杂交后代性状预测的准确度越来越引起重视。
观赏花卉育种的本质是对花卉自然发生的或通过人工方法创造的遗传性变异进行选择,获得符合育种目标的遗传性变异,并增加变异的频率,从而从中选择并育成优良品种的过程。花卉育种的总体目标遵循“多样化、优质、低耗”,然而育种的具体目标会因地因时发生动态的变化。中国台湾地区在20世纪80年代树立了世界蝴蝶兰育种的中心地位,产业发展兴旺,而内地于20世纪90年代后期才开始快速发展,现已成为全球重要的蝴蝶兰生产和消费地之一,但由于育种工作起步晚及品种保护等原因,自主培育的品种极少,而新品种引进受到一定程度的制约,导致蝴蝶兰新品种应用发展缓慢,与产业规模形成巨大反差,育种相关的研究报道和文献资料也不多见[3-5]。
在蝴蝶兰杂交育种过程中,花色与花斑是最直观反映新品种外观特点的重要指标,增加对亲本花朵底色及斑纹类别遗传分离表现的了解程度,将能更有目的的选择亲本,提高目标性状的育种效率。如育种家们曾采用多脉蝴蝶兰(P. Venosa)作为黄花蝴蝶兰的重要原种亲本,其底色为显性遗传,不褪色,而杂交后代的斑纹与花型会受到另一亲本的控制;再如扁梗蝴蝶兰(P. fasciata),又称横纹蝴蝶兰,是黄花蝴蝶兰之母,原生种的花色由淡黄到深黄均有,不褪色,上有红褐色的同心圆条纹,这种条纹能显性地遗传给子代;又例如P. Paifangs Queen的优良大花个体‘Brother被用于进一步育种,花朵白底色上布满大块红色斑块,常被用作亲本培育红花脉纹蝴蝶兰后代[6-7]。本课题组一直持续多年对蝴蝶兰优异种质杂种后代的重要观赏性状和遗传倾向等方面开展跟踪深入的研究分析[8-11],在长期的杂交育种工作中筛选出重要的优良亲本进行目标性状的育种,如Phalaenopsis‘Frigdaas Oxford(黄金豹)是个优异的中小花型黄底紫红斑品种,其底色艳丽,块状紫红斑似豹斑别具特色,花瓣硬挺呈蜡质状,由于其花部性状的明显特征,且育性较好,而成为课题组多年来对黄色系蝴蝶兰及斑纹育种跟踪研究的一个重要亲本。因此本研究选择以Phalaenopsis‘Frigdaas Oxford与Phal. SH49及其正反交后代为材料,探讨花色、花斑类型、花瓣厚度、花朵大小等蝴蝶兰育种中重要的目标观赏性状的遗传规律,为可持续蝴蝶兰亲本的选配及后代观赏性状表型的预测等提供参考。
1 材料与方法
1.1 材料
供试材料为广东省名优花卉种质资源圃的一个杂交组合的亲本和杂交后代群体。以Phalaenopsis ‘Frigdaas Oxford(♀)×Phal. SH49(♂)为正交,Phal. SH49(♀)×Phalaenopsis‘Frigdaas Oxford(♂)为反交。
亲本:Phalaenopsis‘Frigdaas Oxford(黄金豹),黄色蜡质中花,具深紫红色斑块,由本课题组于2007年7月10日,在英国皇家园艺学会登陆;并于2010年通过广东省农作物品种委员会审定。亲本:Phal. SH49,紫红色纸质大花,具细微白镶边,花型圆整。
1.2 方法
1.2.1 采样方法 2012年进行正反交授粉,同年获得正反交果荚进行无菌播种与培养,2013年出瓶种植正反交后代群体(5 cm苗),之后2014年3~5月杂交后代群体首次开花(8 cm苗)。连续两年对正交后代257株、反交后代309株进行重要农艺及观赏性状的调查、测量、记录并拍照。
1.2.2 测定指标 2015~2016年对父母本及杂种后代重要观赏性状进行测量与描述。测量的数量性状9个,包括:株幅、花枝长、花序梗粗度、花朵数、花朵纵径、花朵横径、花瓣长度、花瓣宽度、花瓣厚度;描述调查记录的质量性状3个,包括:花瓣底色、花斑类型、花朵质地[12]。
1.2.3 计算公式 杂种后代的遗传倾向分析相关计算公式如下:变异系数(CV)= 杂种后代标准差/杂种后代平均值×100%;遗传传递力(Ta)=杂种后代平均值/亲中值×100%;杂种优势率(H)=(杂种后代平均值-亲中值)/亲中值×100%。
1.3 数据分析
花朵底色及斑色采用英国皇家园艺协会RHS标准比色卡来判定;数据处理采用Excel软件。
2 结果与分析
2.1 正反交后代群体花部性状特征描述及各分类组群的分离比率
综合花朵主色、花斑类型及花型3个最直观重要的花部性状特征,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代均基本一致的分离为四类组群,连续编号从G01~G04(图1)。杂交亲本Phalaenopsis‘Frigdaas Oxford是黄底色-深紫红斑,由于花斑颜色较深且面积较大时,通常会遮挡住较浅的底色;本实验正反交后代的花朵便存在这种现象,即从正面几乎看不到黄底色的遗传分离,但从花朵背面可以直观的看到黄底色的存在及分离情况;正反交后代的三类组群(G02、G03和G04)花朵正面均被斑色所覆盖,呈现出的主花色均是紫红色,而从背面则可以看到淡黄底色在杂交后代中的存在,仅有组群G01完全呈现深紫红色的花色性状;因此图1示出亲本及正反交后代的正面及背面照能直观清晰地观察区别。另外,由于同一组群中花朵主色因个体差异不完全一致,存在深浅程度的略微差别,因此每组类群示出最浅及最深两朵代表花;具体特征见图1照片所示和表1对应详细描述。
每个后代组群的植株数量及各占正反交后代群体(正交后代总257株、反交后代总309株)的比例值见表1所示;按所占比例值正交后代组群数量大小排列如下:正G02>正G01>正G03>正G04;反交后代組群大小排列如下:反G01>反G03>反G02>反G04,结果表明不同组群的后代类型在正反交中出现的规律有所差别,其中第4种类型(G04)的后代组群在正反交后代中所占的比例都最少。
2.2 正反交后代群体数量性状分离变异
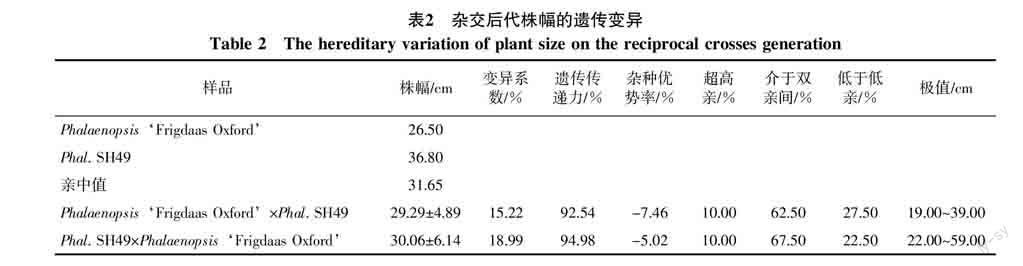
2.2.1 株幅在正反交后代中的表现与分布 株幅大小决定着植株的体量,小株幅是蝴蝶兰选育小株型品种的目标性状之一。试验结果表明,Phalaenopsis‘Frigdaas Oxford与Phal. SH49株幅大小为多基因控制的数量性状遗传。从表2可以看出,正反交后代株幅指数平均值都小于亲中值,成趋中变异。后代的株幅值分别在19.00~39.00、22.00~59.00 cm,CV分别为15.22%、18.99%,说明株幅变异程度大;正反交后代低于低亲的比率分别为27.50%和22.50%,表明Phalaenopsis‘Frigdaas Oxford作母本时更易于将它较小株幅遗传给后代。
2.2.2 花枝长度在正反交后代中的表现与分布
从表3可以看出,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代的花枝长指数平均值都小于亲中值。后代的花枝长度值分别为17.50~51.00、22.00~54.00 cm,变异系数分别为21.72%、26.50%,说明花枝长度变异程度大;正反交后代低于低亲的比率分别为60.00%和75.00%,说明后代中出现短花枝的机率较大,有利于低矮花枝目标性状的获得。
2.2.3 花序梗粗度在正反交后代中的表现与分布
从表4可以看出,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代平均花序梗粗度低于低亲亲本,低于低亲比例分别为95.00%、92.50%,遗传传递力分别为80.37%、80.91%,可见杂交后代花序梗粗细性状有明显趋小的遗传倾向。
2.2.4 花朵数在正反交后代中的表现与分布 从表5可以看出,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代的花朵数量平均值低于低亲亲本,且正反交均未有后代花朵数量超过双亲,低于低亲比例分别为87.50%、95.00%,呈明显花朵数减少的趋势。正反交后代分离广泛,变异系数分别为20.36%、23.71%,说明杂种后代平均花朵数的变异程度较大。
2.2.5 花朵大小在正反交后代中的表现与分布 花朵大小是蝴蝶兰选育大小花型品种的目标性状之一,花朵大小由花朵的纵径与花朵横径共同决定,花纵径与花横径的长度比例也将影响整个花朵的圆整形状。从表6可以看出,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代的花纵径平均值都小于亲中值,成趋中变异,变异系数分别为8.35%、8.05%;花横径呈现同样的表现趋势,即正反交后代的花横径平均值亦都小于亲中值,成趋中变异,变异系数分别为10.50%、8.23%;表明杂种后代花朵大小的变异程度不大,且大多数后代花朵介于父母本之间,多呈中型花大小。
2.2.6 花瓣厚度在正反交后代中的表现与分布
花瓣厚度与花瓣材质及植株的抗性具有一定的关联性,厚瓣花是蝴蝶兰增强品种抗性的育种目标之一。表7分析结果表明,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代的花瓣厚度指数平均值都小于亲中值,成趋中变异。后代的花瓣厚度值分别为0.56~1.12、0.67~1.14 mm,变异系数分别为12.60%、14.09%,说明花瓣厚度的变异程度大;正反交后代高于高亲的比率分别为5.00%、7.5%,表明Phalaenopsis‘Frigdaas Oxford作父本时更易于将它的较厚花瓣遗传给后代。
2.2.7 花瓣大小在正反交后代中的表现与分布
花瓣大小变化通常与花朵的大小呈正相关,从表8可以看出,Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交后代的花瓣长与花瓣宽的平均值均小于亲中值,都成趋中变异;大部分正反交后代的花瓣大小介于双亲间,未有超高亲后代,少量后代低于低亲值,从变异系数值可以看出杂种后代花瓣大小的变异程度都不大。
2.3 正交后代与反交后代的数量性状变异性综合对比
综合上述分析,从变异系数来看,正交后代与反交后代排在前3位的性状均是花枝长>花朵数>株幅,正交后代依次为21.72%、20.36%、15.22%,反交后代依次为26.50%、23.71%、18.99%;且反交后代的变异系数值均大于正交后代,表明正反交后代在花枝长、花朵数、株幅方面比双亲有更大的变异,尤其反交后代变异更大。如图2所示,同一数量性状在正交后代与反交后代中的变异由于组群不同而有明显差别,其中株幅、花序梗粗度、花纵径、花横径、花瓣长5个数量性状都是正交G03与反交G03(第三类群)差异最大;花枝长和花瓣宽两个数量性状是正交G02与反交G02(第二类群)差异最大;花朵数和花瓣厚两个性状是正交G01与反交G01(第一类群)差异最大;而正交G04与反交G04(第四类群)的9个数量性状差异相对都较小,说明Phalaenopsis‘Frigdaas Oxford和Phal. SH49无论正交还是反交得到的第四类群后代的变异都不大。
3 讨论
经济作物可通过传统有效的杂交育种手段实现基因重组得到优良变异,进而获得新优品种或新优株系[15-16]。观赏花卉如兰花是异花授粉的高度杂合的观赏植物,在杂交后代中常产生大量的变异,且兰花观赏性状多属于数量性状,因此育种初期亲本的选配是杂交育种技术中最重要的环节之一。蝴蝶兰的主要观赏價值集中在花上,花色、花斑种类、花型、花朵大小、花朵质地等多样性的变化是培育新优品种的主要育种目标。如何进行有效合理的挑选亲本,丰富蝴蝶兰的遗传多样性,在新品种改良等方面具有实际指导意义[6,16]。本研究对比蝴蝶兰Phalaenopsis‘Frigdaas Oxford和Phal. SH49正反交组合结果表明,后代分离的类别和趋势差别不大;正反交组合的杂种F1代在以上数量性状的平均值表现均较中亲值明显降低,类似的现象也在其他蝴蝶兰品种的正反交组合[8]以及观赏花卉如非洲菊[17]、荷花[18]等植物中出现过。本研究发生的数量性状平均值降低的分离表现对现今选育小型或微型后代个体是正向的遗传分离趋势。同时正交组合与反交组合内的变异幅度都较大,尤其在质量性状上变化丰富,且出现了亲本没有的花斑变化,这将为新个体或新单株的选育提供更多的选择。
课题组多年来一直对采用Phalaenopsis‘Frigdaas Oxford(黄金豹)做亲本的子代群体开展花色和花斑分离规律的持续跟踪研究,本研究中将其与一个大红花纯色品种Phal. SH49进行正反交,实验结果发现底色与斑色发生了有规律的分离。正反交后代的3个组群(G02、G03和G04)的花朵由于红色斑色的覆盖,正面几乎看不到亲本Phalaenopsis‘Frigdaas Oxford黄底色的遗传分离,而从背面则可以清晰观察到淡黄底色在这些杂交后代中的存在,即亲本Phalaenopsis‘Frigdaas Oxford的黄色在后代遗传中明显变浅;同时本研究的正反交后代中均出现了近似亲本Phalaenopsis‘Frigdaas Oxford斑色的纯深紫红色-蜡质类型(G01),却未获得全底色的类型;值得关注的是该类群的花朵出现了与课题组前期两个实验一致的遗传变化,即花瓣与花朵大小较其他组群明显偏小,且花瓣与萼片形状都较其他组群更尖。对比课题组之前的两个研究,发现实验1 Phalaenopsis‘Frigdaas Oxford自交后代的花朵黄底色出现变浅或加深的分离变化[9],而紫红色斑块在子代中呈现出不同大小、面积和形状的分离变化,出现全斑完全覆盖住底色的类型花,但未发现无斑纯黄底色类型花;同时全斑类型的花型出现了明显变化,花瓣与萼片都变尖,花大小也较其他花色类型明显变小;实验2采用Phalaenopsis‘Frigdaas Oxford为母本,与大白花纯色品种Phal. 316进行杂交[10],研究发现杂交F1代在花色、斑纹、花朵质地、唇瓣须状物等方面都发生了分离,其中与实验1的自交后代相反的是获得了纯黄色无斑类型,却未获得全斑覆盖底色的类型;底色既有父本的白色,也有母本的黄色,但黄色出现变浅现象;紫色斑纹大小和形状成一定规律分离变化,其中大面积斑纹的后代组群中也出现了与本研究和实验1类似的花朵较小的分离特征。
综上分析表明,蝴蝶兰花色的遗传能力与覆盖能力依次是红色>黄色>白色,即通过杂交育种可获得黄底红斑、白底红斑的后代,而难获得红底黄斑、红底白斑的后代花色,因此通过对蝴蝶兰花底色、斑色、斑类型、花瓣材质等花部重点性状的更多了解,将能提高目的性育种的实现概率,为杂交亲本的选择提供更有效的指导性参考。
参考文献
[1] Frowine S A. Moth Orchids: the Complete Guide to Phalaenopsis[M]. London: Timber Press, 2008: 12-130.
[2] Griesbach R J. Development of Phalaenopsis orchids for the mass-market[M]. Trends in new crops and new uses. Alexandria: ASHS Press, 2002: 458-465.
[3] 李振坚, 王 雁, 彭镇华, 等. 兰花在全球花卉贸易中的地位及发展动态[J]. 中国农学通报, 2008, 24(5): 154-159.
[4] 丁朋松, 郭文姣, 孙纪霞, 等. 蝴蝶兰杂交育种研究进展[J]. 安徽农业科学, 2014, 42(7): 1 954-1 956.
[5] 朱根发. 蝴蝶兰种质资源及杂交育种进展[J]. 广东农业科学, 2015(5): 31-38.
[6] 黄玮婷, 曾宋君, 吴坤林, 等. 蝴蝶兰属植物杂交育种研究进展[J]. 热带亚热带植物学报, 2012, 20(2): 209-220.
[7] Christenson E A. Phalaenopsis: A Monograph[M]. London: Timber Press, 2001: 39-291.
[8] 陈和明, 吕复兵, 朱根发, 等. 1个正反交蝴蝶兰若干性状在F1的遗传表现[J]. 华北农学报, 2011, 26(增刊): 28-33.
[9] Lü F, Yu N, Zhao X, et al. Genetic diversity analysis of Phalaenopsis ‘Frigdaas Oxford using SRAP markers with reference to those genes responsible for variations in the pigmentation of petals and sepals[J]. Journal of Horticultural Science & Biotechnology, 2011, 86(5): 486-492.
[10] 李 佐, 肖文芳, 陈和明, 等. 蝴蝶兰Phalaenopsis ‘Frigdaas Oxford 和Phal. 316杂交F1代性状分离研究[J]. 热带作物学报, 2014, 35(5): 854-861.
[11] 李 佐, 肖文芳, 陈和明, 等. 蝴蝶兰杂种一代的花朵材质、花斑及花型遗传分化的初步探讨[A]// 中国观赏园艺研究进展. 北京: 中国林业出版社, 2015: 67-70.
[12] 陈和明, 朱根发, 吕复兵, 等. 蝴蝶兰新品种DUS测试指南的研制[J]. 中国农学通报, 2014, 30(10): 182-185.
[13] International union for the protection of new varieties of plants (UPOV). TG/213/1. Phalaenopsis (Phalaenopsis Blume.) guidelines for the conduct of tests for distinctness, uniformity and stability[S]. Geneva, 2003. 04. 09. http: //www.upov.int/en/publications/tg-rom/tg213/tg_213_1.pdf.
[14] International union for the protection of new varieties of plants (UPOV). TG/213/1. Phalaenopsis (Phalaenopsis Blume.) guidelines for the conduct of tests for distinctness, uniformity and stability[S]. Geneva, 2013. 03. 20. http: //www.upov.int/edocs/tgdocs/en/tg213.pdf.
[15] 崔艷波, 陈 慧, 乐文全, 等. ‘京白梨 与 ‘鸭梨 正反交后代果实性状遗传倾向研究[J]. 园艺学报, 2011, 38(2): 215-224.
[16] 许 玲, 魏秀清, 章希娟, 等. 龙眼正反交杂种后代果实经济性状遗传倾向研究[J]. 热带作物学报, 2015, 36(2): 224-228.
[17] 李绅崇, 李淑斌, 蒋亚莲, 等. 非洲菊品种间杂交主要观赏性状在F1代的遗传表现[J]. 云南农业大学学报学报, 2007, 22(2): 197-201.
[18] 姜 莉, 陈发棣, 滕年军, 等. 荷花部分性状在F1代的遗传表现[J]. 南京农业大学学报, 2009, 32(3): 36-41.
猜你喜欢
作文与考试·小学高年级版(2022年19期)2022-05-30
现代苏州(2021年23期)2021-12-27
快乐作文(1.2年级)(2021年11期)2021-12-19
小学生优秀作文(低年级)(2020年12期)2021-01-14
青少年日记·小学生版(2019年1期)2019-09-02
故事作文·低年级(2019年8期)2019-08-20
现代园艺(2017年13期)2018-01-19
中华诗词(2017年3期)2017-11-27
小主人报(2015年18期)2015-09-16
小天使·一年级语数英综合(2015年8期)2015-07-06
