
四川引种邓恩桉生殖生长特性观测及种子萌发研究
2017-06-28 15:10李佳蔓郭洪英
四川林业科技 2017年3期
黄 振,陈 炙,李佳蔓,郭洪英
(四川省林业科学研究院,四川 成都 610081)
四川引种邓恩桉生殖生长特性观测及种子萌发研究
黄 振,陈 炙,李佳蔓,郭洪英*
(四川省林业科学研究院,四川 成都 610081)
通过研究引种邓恩桉的花期发育和种子萌发能力,掌握邓恩桉生殖生长规律,为邓恩桉种子园的建设提供理论依据。本研究以位于绵阳梓潼县的邓恩桉引种试验林中开花的单株为研究对象,结果表明,引种邓恩桉花期2月中旬至10月中下旬;依据形态变化,花期可分为:花芽分化期、花序形成期、蒴盖变色期、蒴果膨大与脱落环形成期、盛花期和种子发育期。同一花序上的花蕾发育进程基本一致,同一花枝上靠近基部的花蕾发育速度比顶部花蕾的要快,不同位置花枝的发育表现出随机态势。蒴果内有3~4个子房腔,瘪粒数量多,有效种子数量少,平均每个蒴果内含有5粒能正常发育成苗的种子。
邓恩桉;生殖生长;花芽分化;花粉;种子萌发
桉树(Eucalyptus)是四川重要的用材林和工业原料林树种之一。早在20世纪80年代,四川省林科院就开始进行了桉树的区域性引种试验,经过多年的筛选,选择出很多适宜于四川栽种的桉树类型,如巨桉(E.grandis)、邓恩桉(E.dunnii)、直干蓝桉(E.maidenii)、赤桉(E.camaldulensis)等,其中邓恩桉(E.dunnii)兼有优良的纸浆材树种和大径材树种[1],同时具有较强的耐寒能力,能种植在川中丘陵区及位置靠北的盆周山地,是扩大桉树种植区域的理想树种[2]。
邓恩桉是桉树属的双蒴盖亚属(E.subgenussymphyomyrtus)蓝桉组(SectionMaidenaria)多枝桉系(seriesviminales)高大乔木[3]。与尾巨桉、柳桉等已实现工厂化生产的桉树相比,邓恩桉无论是组织培养[4,5]还是扦插繁殖[6],均存在生根困难的问题[7],技术瓶颈导致邓恩桉的工厂化生产尚需时日。目前邓恩桉的苗木培育以实生苗为主,但由于种子主要来源于澳大利亚新南威尔士州的天然林,且邓恩桉需要10a以上才会开花结实,种子产量小且价格昂贵。已有报道的邓恩桉种子园包括:贵州惠水县邓恩桉实生苗种子园[8]、广西环江县邓恩桉无性系种子园[9],但邓恩桉开花结实困难的问题依然严重,严重制约了种子园的发展。因此,研究邓恩桉的生殖生长规律,破解邓恩桉种子产量低的难题是当前邓恩桉育种的研究重点。
本研究以2015年底发现的开花结实邓恩桉为对象,以其生殖生长特征为研究目标,通过观察花芽分化和花序发育特点、花粉形态、蒴果和种子形态,种子萌发能力检测,旨在揭示引种在川北地区的邓恩桉生殖生长特性,为建立邓恩桉种子园奠定理论和实践基础。
1 材料与方法
1.1 引种试验林简介
邓恩桉种子于2010年春季播种育苗,2011年上山造林,造林密度为2 m×3 m。2015年秋,观察到3株邓恩桉出现了花芽分化现象。2015年以这3株邓恩桉中树高和枝下高较矮,相对容易取样和观测的1株为研究材料。
1.2 试验地概况
邓恩桉引种试验林位于绵阳市梓潼县双板乡凉泉村,土壤紫黄泥土,属白垩系下统母岩发育而成,土壤胶粘重,色调浅黄,土体构造与紫泥土相似,小块结构,土壤稍紧,pH值7.6,呈微碱性。
1.3 试验方法
花期观测 从2月中旬开始至10月中下旬,每30 d上山观测花蕾变化情况,当发现花蕾形态和颜色发生重大改变时,观测周期缩短为15 d。分别记录枝条内和枝条间花蕾的发育速率。
减数分裂判断 样品固定:每次观测完花期后,用装配有11号刀片的3号解剖刀沿着花蕾上的脱落环把花蕾切成两半,把带有花药的蒴盖投入含有卡诺固定液的离心管中固定并带回带回实验室。小孢子发育观测:用小镊子从离心管中取出蒴盖,把花药从蒴盖内挑出放在载玻片上,切去花丝,把10个~20个花药集中放置,滴1滴醋酸洋红染色液,用镊子将花药反复夹碎,使花药内的小孢子尽量释放,轻轻盖上盖玻片,再用打火机火焰从载玻片下方烘烤1 s~2 s,使染色体充分进入细胞内。随后将载玻片放入光学显微镜(BX51)上观察。
蒴果结构观测 2017年1月上旬,采集邓恩桉蒴果带回实验室,蒴果放置在30℃烘箱中干燥,直至蒴果开裂。分别采集蒴果横切和纵切照片、种子形态图。
种子萌发 把一个蒴果内的种子和杂质全部撒在一个含有0.5 cm厚脱脂棉花的90 mm×90 mm方形塑料培养皿中,湿润棉花后,把培养皿放入25℃培养箱中培养,每隔1 d加1次蒸馏水,以保持水分。种子发芽后,取出幼苗拍照。
2 结果与分析
2.1 花期观测与发育节点判断
观察发现,邓恩桉花芽在分化后,即可在形态上与叶芽进行区分,花芽的发育在形态学上可以分为以下6个阶段。
(1)花芽分化期(图1-A):从2月中下旬开始,花芽开始萌动,从形态上即可分辨出花芽和叶芽,花芽中部饱满,显著大于梭型叶芽,在整个分化期,花芽逐渐长大,未分化出花蕾,该阶段持续到6月中下旬,消耗的时间最长。
(2)花序形成期(图1-B,C):花蕾表层的鳞片开始干裂脱落,各个幼嫩的花蕾从芽中长出并向各个方向伸展,形成伞房花序,该阶段持续20 d~30 d,持续的压片观察发现直到该阶段末期,小孢子母细胞才出现在花药囊。
(3)蒴盖变色期(图1-D):7月中旬,花蕾直径长至2 mm,蒴盖颜色开始变黄,能清晰分辨出蒴盖与蒴果。用解剖刀纵切花蕾,发现花药和花丝充满了花蕾,花丝卷曲在一起(图2-A),镜检发现此时小孢子母细胞开始减数分裂,主要集中在前期Ⅰ的细线期(图2-B),染色体呈单条细线状,看不到成双的染色体,染色体的排列呈无序的丝状排列。细胞核的体积很大,细胞质浓稠,核仁非常明显,部分染色体一端与核仁相连,整体形态似花束状。这说明蒴盖颜色的变化是减数分裂即将开始的形态学判别标志。

图1 邓恩桉花芽发育过程中形态学观测Fig.1 Morphology of E.dunnii developing flower budsA:花芽分化起始期;B-D:伞房花序形成期;E:蒴盖变色期;F:蒴果膨大与脱落环形成期,箭头示脱落环;G:盛花期;H:授粉后的蒴果形态。
(4)蒴果膨大与脱落环形成期(图1-E、F):8月份中上旬,花蕾直径长至3mm,蒴盖颜色持续加深,部分花蕾可见脱落环初步形成。用沿着蒴盖下方1 mm处轻轻环割花蕾,可将蒴盖完全去掉,露出花丝和柱头,花丝排列规则紧密,花药菱形呈透明状,花柱呈淡绿色,柱头呈深绿色(图2-C),此时小孢子母细胞减数分裂主要集中在中期Ⅰ,镜检可见:染色体运动到赤道板,细胞内出现纺锤丝并形成纺锤体(图2-D)。
(5)盛花期(图1-G):9月中旬,脱落环裂开一个口,在花丝张力的作用下,蒴盖从开口处被抬起并掉落,花丝从花蕾中全部伸展(图2-E),花药呈黄色或淡黄色,柱头呈淡绿色,上有粘液,可俘获风吹或昆虫携带的花粉。镜检可见成熟花粉粒,花粉粒为正四面体结构,未见明显萌发孔(图2-F)。
(6)种子发育期(图1-H):授粉结束后,花丝和柱头逐渐凋零,蒴果上表面逐渐由绿色变成黑灰色,质地由软变为坚硬。受精卵在子房内开始发育,次年1月发育成熟。
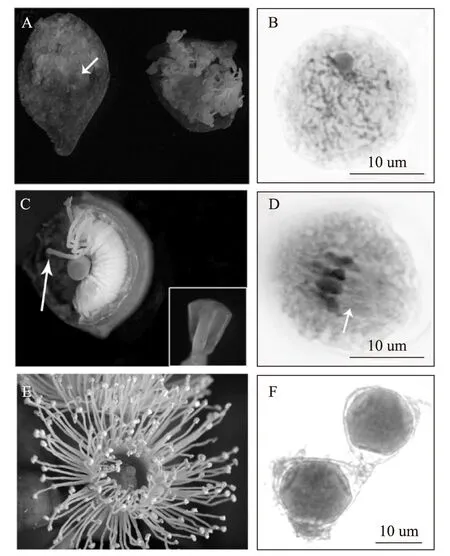
图2 花蕾内部形态与小孢子发育进程Fig.2 The internal morphology of E.dunnii flower bud and the development of microsporeA:蒴盖变色期花蕾纵切,箭头示花药位置;B:处于细线期的小孢子;C:蒴果膨大与脱落环形成期的花丝与柱头形态,箭头示花药位置,右下角为透明状菱形花药放大图;D:处于中期Ⅰ的小孢子,细胞中央为整齐排列的染色体,箭头示牵引染色体的纺锤丝;E:盛花期的花蕾,花丝完全伸展,花药黄色或淡黄色,柱头绿色;F:盛花期时,花药内完全成熟的花粉粒。
2.2 花枝内与花枝间花蕾发育进程比较
邓恩桉花序由5个~7个花蕾组成,同一个花序的花蕾直径相互之间差异不大,镜检显示花蕾内小孢子的发育进程基本一致。但不同位置上的花蕾平均直径不同,表现为:随着花序着生位置从基部向顶部,花蕾的平均直径逐渐变小。直径最大差值>2 mm。这说明基部花蕾的发育速度比顶部花蕾的发育速度要快。基部花蕾进入盛花期的时间要早也证明了此判断。
同一母株不同位置的花枝上花蕾的平均直径却表现出随机的态势,即花序的整体发育与小花枝所处的位置无关。向阳处花枝的花蕾平均直径显著大于阴处花枝的花蕾平均直径。这表明向阳处花枝花蕾的发育速度比较快,阳光对花蕾的发育有良性刺激。
2.3 蒴果形态与结构观测
研究发现,一个花序上一般只能存留2~4个蒴果,与盛花时花序盛开5~6个花蕾相比,这说明授粉后,有一半左右的花蕾或蒴果会凋落,其原因可能是率先完成授粉的蒴果会分泌激素致使花蕾凋落,也可能是母树营养不足,为了保证部分种子的正常发育,母树在其它花蕾基部形成离层促使其脱落。
邓恩桉蒴果形态见图3-A,B,从图中可以看出,蒴果内有3~4个子房腔,分别对应顶部开裂成“三菱形”和“十字形”。大部分邓恩桉蒴果以十字形为主。
两类蒴果的纵切结构完全一致,种子在室内呈上下排列成弧形(图3-D),上面的种子长度较长,能达到1 mm(图3-E),而中部的种子发育畸形,长度仅为0.5 mm左右,这些为未受精的瘪粒,不能发育成苗,室的底部仅留下一个黄色的萌发痕迹(图3-F),由于种子和瘪粒形态过小、颜色接近,难以统计蒴果内种子的数量。
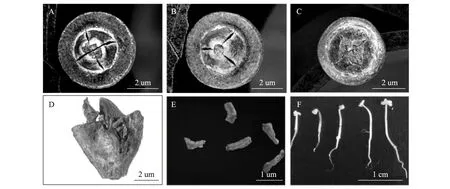
图6 蒴果与种子形态及种子萌发Fig.3 Capsule and seed morphological characteristics and seed germination of E.dunniiA:四室蒴果俯视;B:三室蒴果俯视;C:四室蒴果横切;D:蒴果纵切;E:邓恩桉种子形态;F:萌发的邓恩桉幼苗。
2.4 种子萌发
由于种子颜色和大小与瘪粒很接近,因此,种子发芽试验采用了蒴果内含物一并播种的方法,试验共播种了6个培养皿,共获得了30株幼苗,平均5株·蒴果-1。
3 结论与讨论
本研究的邓恩桉引种试验林位于绵阳市梓潼县,该区域是邓恩桉分布的最高纬度,高于浙江台州市台州湾[10,11],远高于邓恩桉在湖南的北缘分布区[12]。尽管邓恩桉在我国南方普遍引种,但对其开花结实等生殖方面的报道甚少,据称在四川黑龙滩水库以及云南种植的邓恩桉有长势差的个别植株能开花,贵州林科院的个别植株也有开花结实,但不育[12],桂林引种的邓恩桉10a后开花,但无结实报道[13],广西邓恩桉无性系种子园尽管进行了母株生理生化特性[11]和赤霉素处理[14]的研究,但邓恩桉开花结实困难的问题依然严重。
通过对邓恩桉引种试验林的生殖生长特性进行观测并获得了自由授粉的子代苗,获得了引种邓恩桉在川北丘陵地带开花习性、花蕾和蒴果发育的时间和形态学特征等邓恩桉生殖生长特征。具体包括:
(1)邓恩桉在2月中下旬开始进入生殖生长,花期持续8个月,依据形态学分类,花芽的发育可分为6个阶段。雄花的减数分裂开始时期与蒴盖颜色的变化相关性较高,颜色的变化能作为减数分裂开始的形态学判别标志。这一现象与邓恩桉同一亚属——双蒴盖亚属——的尾叶桉(E.urophylla)、尾细桉(E.urophylla×E.tereticornis)和尾巨桉(E.urophylla×E.grandis)的研究结果是一致的[15,16]。
(2)花序的整体发育与花枝位置无关,向阳处花蕾的发育速度比较快。花枝基部花序的发育速度比顶部花序的发育速度要快。这些结论为邓恩桉种子园的营建奠定基础。
(3)引种邓恩桉蒴果中子房腔数量为3~4个,稍低于常见桉树子房腔的数量(4~5个)。而邓恩桉授粉后蒴果内种子的发育与其它桉树生殖生物学研究一致[3],即桉树内具有很多假胚珠,假胚珠长成瘪粒,胚珠形成的真正种子数量很少,每个蒴果4~10粒。蒴果内种子萌发能力测试获得了平均5株/蒴果的结果,说明引种邓恩桉种子发育正常,具有发育成苗的能力。
研究获得了邓恩桉生殖生长特性的初步资料,特别是花芽、花蕾、蒴果等阶段表型性状的变化特征,尚未深入研究其内部机理和调控措施,下一步既需要继续深入研究生殖生物学特性,如长中大小孢子的发育规律、授粉和花粉萌发、双受精过程等节点,还需要探索研究导致生殖生长的环境原因,比如土壤结构、养分组成、水分温度条件等因素,以寻求切实提高邓恩桉的开花结实率的技术方法,早日实现邓恩桉种子生产的实用化。
[1] 陈茜文,杨模华,李志辉,等.耐寒桉树的纤维形态特征的变异[J].中南林学院学报,2002,22(04):61~65.
[2] 罗建中,曹加光,卢万鸿.邓恩桉等6种桉树的耐寒能力研究[J].桉树科技,2011,28(02):34~38.
[3] 王豁然.桉树生物学概论.北京东黄城根北街[M].科学出版社,2010.
[4] 宋建英.邓恩桉的组织培养和植株再生[J].林业科学,2010,46(6):138~145.
[5] 宋建英.邓恩桉种子组织培养的研究[J].中南林业科技大学学报,2008,28(6):75~80.
[6] 林成立.邓恩桉扦插繁殖试验[J].福建林业科技,2005,32(2):80~84.
[7] 宗亦臣,郑勇奇,常金财.邓恩桉国内引种和繁育技术研究进展[J].中国农学通报,2010,26(5):82~86.
[8] 李天会,邓伯龙,谢耀坚,等.邓恩桉种子园营建技术初报[J].林业资源管理,2007(02):82~85.
[9] 陈建波,李昌荣,项东云,等.一种选择建立邓恩桉嫁接种子园的方法.CN 104115677 A,2014年10月29日.
[10] 林益,瓮颖,李贺鹏,等.邓恩桉幼苗生长和光合特性对盐胁迫的响应[J].浙江林业科技,2014,34(03):33~38.
[11] 李昌荣,郭东强,周维,等.邓恩桉嫁接种子园开花生理生化特性比较[J].中国农学通报,2014,30(25):20~23.
[12] 刘友全,李志辉,马英,等.速生耐寒邓恩桉在桉树分布北缘区的适生性[J].中南林业科技大学学报,2007,27(2):66~69.
[13] 张建明.邓恩桉引种首次在桂林开花[J].广西林业,2001(6):21.
[14] 周维,李昌荣,郭东强,等.赤霉素对邓恩桉嫁接种子园花蕾生长的影响[J].福建林业科技,2015,42(02):16~19.
[15] Yang J; Lan J,Yao P,et al.Comparative microsporogenesis and flower development in Eucalyptus urophylla × E.grandis.Journal of Forestry Research,2016,27(2):257~263.
[16] Yang J; Yao P,Li Y,et al.Induction of 2n pollen with colchicine during microsporogenesis in Eucalyptus.Euphytica,2016,210(1):69~78.
A Study of Reproductive Growth Characteristics and Seed Germination ofEucalyptusdunniiIntroduced in Sichuan Province
HUANG Zhen CHEN Zhi LI Jia-man GUO Hong-ying*
(Sichuan Forestry Academy,Chengdu Sichuan 610081,China)
Studying the flowering development and seed germination of introducedEucalyptudunniiand mastering its reproductive growth law can provide a theoretical basis for seed garden.In this study,a blooming plant growing in Zitong County was selected as the experiment subject.The results were as follows:florescence date ofEucalyptudunniibeing from Mid and late February to mid and late October.Depending on morphologic change,flowering could be divided into 6 phases:Flower bud differentiation stage,inflorescence formation stage,opercle color change stage,opercle enlargement and shedding ring formation stage,flowering period,seed development stage.The development process of the buds on the same inflorescence was basically the same,and the growth of buds near the base of the same branch was faster than that of the top buds.The development of flower buds in different positions showed a random trend.Opercle had 3 to 4 ovary cavities,the number of shriveled seeds was far more than the number of effective seeds,and on a average,each capsule containing five normal seeds could develop into seedlings.
Eucalyptus,Reproductive growth,Flower bud differentiation,Pollen,Seed germination
2017-02-28
四川省基本科研业务费项目:邓恩桉花芽分化及小孢子发育的细胞学特性研究,(JB2016-14)。
黄振(1984-),男,博士,助理研究员,研究方向:林木遗传育种。邮箱:hz7210@126.com
郭洪英(1973-),女,硕士,研究员,研究方向:林木遗传育种。
10.16779/j.cnki.1003-5508.2017.02.002
S722.3
A
1003-5508(2017)03-0006-05
猜你喜欢
甘肃农业大学学报(2021年4期)2021-09-22
辽宁林业科技(2021年3期)2021-07-08
山西水土保持科技(2020年2期)2020-12-22
扬子江诗刊(2018年2期)2018-11-13
扬子江(2018年2期)2018-03-24
当代体育·扣篮(2016年15期)2016-09-27
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
中国粮油学报(2015年5期)2015-02-06
