
北江干流河岸带不同植被类型植物物种多样性分析
2017-09-12 07:36赵清贺徐珊珊马丽娇吴长松
水土保持研究 2017年5期
赵清贺, 徐珊珊, 马丽娇, 吴长松
(1.河南大学 黄河中下游数字地理技术教育部重点实验室, 河南 开封 475004; 2.河南大学 环境与规划学院, 河南 开封 475004)
北江干流河岸带不同植被类型植物物种多样性分析
赵清贺1,2, 徐珊珊1,2, 马丽娇1,2, 吴长松1,2
(1.河南大学 黄河中下游数字地理技术教育部重点实验室, 河南 开封 475004; 2.河南大学 环境与规划学院, 河南 开封 475004)
以北江干流河岸带4种典型植被类型(竹林、草地、混交林和桉树林)为研究对象,采用样方法调查分析其群落特征和物种多样性,运用冗余分析(RDA)探讨北江河岸带植物物种多样性与环境因子之间的关系。结果表明:北江河岸带植物物种受热带季风气候影响,植物种类繁多,共调查到59科116属136种植物,群落科属组成分散;4种植被类型的物种丰富度、Shannon-Wiener多样性和均匀度指数变化规律一致,均以混交林最高,竹林次之,桉树林最低,但各植被类型间差异并不显著,说明北江河岸带不同植被类型维持物种多样性的异质性较小,其作用更多体现在为不同物种提供栖息地;RDA结果显示,植物物种丰富度、均匀度和多样性指数与乔木层高度、草本层盖度和海拔均呈正相关,而与至河流距离和土壤含水量呈负相关,在选取的7个环境因子中,海拔对北江河岸带植物物种多样性的影响最大,土壤含水量影响次之,至河流距离影响最小。
河岸带; 物种多样性; 环境因子; 冗余分析; 北江流域
河岸带是水生生态系统和陆地生态系统的交错过渡带,具有明显的边缘效应和独特的生态功能与服务,同时是敏感和脆弱的生态区域。植被是河岸带生态系统的一个重要组成部分,其类型、组成和结构配置的变化将引起流域生态环境的一系列变化,因此对河岸植被进行结构特征的分析是研究河岸带生态系统功能与服务的基础[1-2]。物种多样性能有效地指示群落组成和结构异质性以及群落与自然环境因素的关系[3-5],因此国内外许多学者对不同生态系统的物种多样性及其与环境因子的关系作了大量研究,取得了诸多成果,如物种多样性和种间关系对环境因子的响应等,研究对象涉及森林、草原、沙地、湿地、草甸苔原等,而环境因子方面侧重于海拔、水热因子和土壤理化性质等[4,6-10],但是针对河岸带的研究较少。近年来,剧烈的人类活动如城市扩张、农田开垦、采砂、污染物的排放、水利水电工程建设以及旅游资源的开发等,给河岸带植物群落的组成、结构和演替及其物种多样性带来显著影响,河岸带生态系统功能的发挥和服务的持续供给面临严峻挑战,引起人们和生态学研究者的日益重视[2,11-16]。因此,揭示河岸带植物物种的组成和结构及其对环境因子的响应可在一定程度上为河岸带物种多样性的保护和生态系统服务的维持提供支持[13-14,17]。北江是珠江流域第二大水系,许多学者对其水文以及水质的报道比较多[18-20],而较少对其河岸带植被进行研究,尤其是关于其物种多样性及其与环境因子关系方面的研究。鉴于此,本文初步分析北江河岸带不同植被类型特征和物种多样性及其与环境因子之间的关系,以期为北江河岸带生态系统完整性的保护提供基础依据。
1 研究区概况
北江属珠江水系,位于广东省北部,流域面积为46 710 km2,占珠江流域面积的10.3%,涉及的行政区主要为韶关市、英德市和清远市[19-20](图1)。流域内多为山地丘陵,年平均气温18~21 ℃,但春季由于冷暖空气交替频繁,常出现“乍冷乍暖”天气,年最高温在7月、8月,常年≥35 ℃。年降雨量1 300~2 400 mm,其空间分布是由南向北递减,夏季常有暴雨,年降水主要集中在4—9月,其中5月降水持续时间最长,强度比较大,往往造成洪涝[18]。本区农业发展的主要土壤资源是红壤、赤红壤、黄红壤及黄壤,占总面积的70%,良好的渗水蓄水能力和养分可为各种植物和作物的生长发育提供条件。同时,受亚热带季风气候影响,研究区具有高温多雨和湿热同期的环境特征,可为植物、动物、微生物等提供良好的栖息地,具有我国南方物种基因库的美誉[21]。另一方面,近20 a来北江流域内城市建设造成的不透水面积增加、农业和其他活动造成的水土流失和植被下降等降低了流域的贮水能力,间接地增加了径流量,加之地形起伏大,导致流域内水土流失的进一步加剧,植被覆盖率呈下降趋势[19-20]。

图1 研究区位置与样点分布
2 材料与方法
2.1 研究样地调查
2014年6月,采用群落样方调查方法对北江流域河岸带植被物种多样性进行野外调查(图1)。共设置26个样地,每个样地设置面积为20 m×20 m的标准样方(每个样方内设置5个1 m×1 m草本小样方)。各样地观测植物群落特征和种类组成,记录每个样方的乔木层、灌木层和草本层的总盖度、植物物种名、群落盖度、株数和平均高度,同时用环刀法采集样方内0—20 cm土样,用自封袋密封,带回实验室测定土壤湿重(精确到0.001 g),然后在85℃烘干箱内烘干至恒重后称重,测定土壤容重与含水量。使用GPS对调查样地进行定位,记录样地海拔、经纬度和土壤质地等。
2.2 数据处理及分析
植物群落物种多样性指数在生态学上通常用来表示群落内植物种类的丰富程度及物种分布的均匀程度[3]。根据北江河岸带的植物数据,26个样地涉及4种植被类型,包括7个竹林样地、5桉树林样地、8个混交林样地(主要为常绿阔叶林)和6个草地样地。根据Braun-Blanquet的盖度—多度等级制确定盖度等级[19],来反映北江河岸带的植物群落特征。采用PC-ORD 5软件,选取物种丰富度指数(S)、均匀度指数(E)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)为物种多样性指标,对研究区不同植物类型的草本层物种多样性进行分析和比较,并利用SPSS 17对物种多样性指数进行单因素方差分析以分析不同植被类型的差异显著性。4个多样性指数计算公式如下:
物种丰富度(S)
S=样地内出现的物种数
(1)
Pielou 均匀度指数(E)
E=H/lnS
(2)
Shannon-Wiener 多样性指数(H)
H=-∑PilnPi
(3)
Simpson多样性指数(D)
(4)
式中:Pi为第i种植物的重要值;S为样方内出现的物种数。
本研究选取的环境因子主要包括海拔、至河流距离、土壤含水量、乔木层高度和盖度、草本层的高度和盖度,共7个指标,使用Canoco for windows 4.5软件进行冗余分析(RDA)[22],通过分析北江河岸带植物物种多样性与环境因子的关系,揭示影响研究区物种多样性的主导环境因子。
3 结果与分析
3.1 北江河岸带植物物种组成
根据26个样地的调查结果统计,北江河岸带植被物种共调查到136种,隶属59科116属(表1)。其中,禾本科(Gramineae)、菊科(Compositae)、大戟科(Euphorbiaceae)、蓼科(Polygonaceae)、豆科(Leguminosae)、桑科(Moraceae)和锦葵科(Malvaceae)等科植物物种数目较多(>4种)、多度较大(>10),天南星科(Araceae)、苋科(Amaranthaceae)和荨麻科(Urticaceae)等科物种数目相对较少(<4种),但多度较大(>10)。在所有植物科中仅含一个物种的有39科,占总科数的66.1%(表1);在所有植物属中,仅含有一个物种的有99属,占总属数的85.3%;所有物种中,出现频率较高的有吊丝球竹(Dendrocalamopsisbeecheyana)、空心莲子草(AlternantheraPhiloxeroides)、海芋(Alocasiamacrorrhiza)、香丝草(Conyzabonariensis)、狗脚迹(Phyllanthuscochinchinensis)、火炭母(Polygonumchinense)、假臭草(Praxelisclematidea)、苎麻(Boehmerianivea)、五月艾(Artemisiaindica),但仅在一个样地中出现的物种有62种,占总物种数的45.6%,表明研究区植物物种种类多,但组成比较分散。不同植被类型对比表明,仅有8种植物在4种植被类型中均有出现(表2),但出现频率差异较大,其中在混交林中出现频率最高,其次是竹林,草地最低;仅在单一植被类型样地出现的物种数较多(表3),竹林、桉树林、混交林和草地中分别为18,13,27,15种。
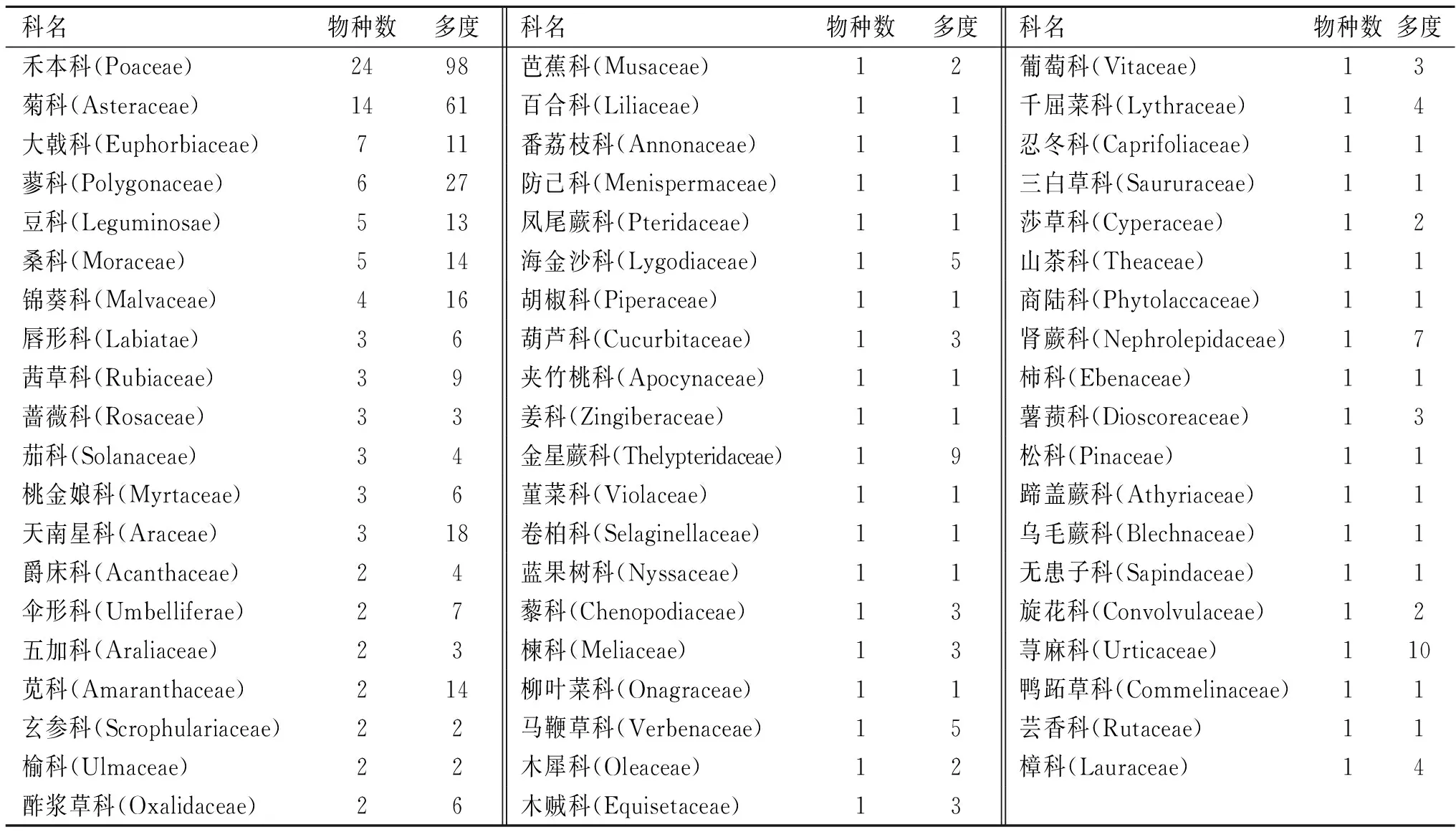
表1 北江河岸带植物科的物种数和多度
3.2 不同植被类型物种多样性分析
由图2看出,北江河岸带植物群落的物种丰富度(S)总体较高,仅2个样地(1个竹林和1个草地)低于10,说明河岸带维持物种丰富度作用明显。4种植被类型以混交林物种丰富度最高,草地次之,桉树林最低;竹林的物种丰富度在不同样地间变异最大(35.1%),桉树林样地间变异最小(6.1%)。桉树为速生丰产林,广泛分布于广东乃至华南地区,因其对水分和土壤养分需求极大,对当地原生物种有极大的抑制性,致其林下草本层的减少造成桉树林物种丰富度较低。草地阳光较为充足,加之草本植物植株较小,可充分利用微环境斑块增加丰富度[21]。研究区物种均匀度指数(E)为0.08~0.62,不同植被类型间变异较大,混交林物种均匀度明显高于竹林和草地,说明北江河岸带4种典型群落的物种组成相对不均。北江河岸带植物的Shannon-Wiener多样性指数(H)变化范围为0.14~1.83。一般认为H>1属正常,H<1时则可能受到其他环境因子的扰动,本次调查的26个样地中有9个样地的H<1,其中竹林和桉树林均为2个(占本类型样地比例分别为28.6%和40.7%),混交林1个(占12.5%),草地4个(占66.7%)。Simpson多样性指数(D)常用来反映物种在不同群落中的地位与作用,包涵较多的群落结构信息,同时较物种丰富度具有更高程度的定量化。北江河岸带混交林Simpson多样性最高、物种组成最复杂,草地最低、组成最为简单。整体上,北江河岸带植物多样性维持在较高水平,虽浮动空间较大,但单因素方差分析结果表明,物种丰富度、物种均匀度、物种多样性在不同植被类型间均无显著差异(p>0.05)。

表2 在4种植被类型样地中均出现的物种
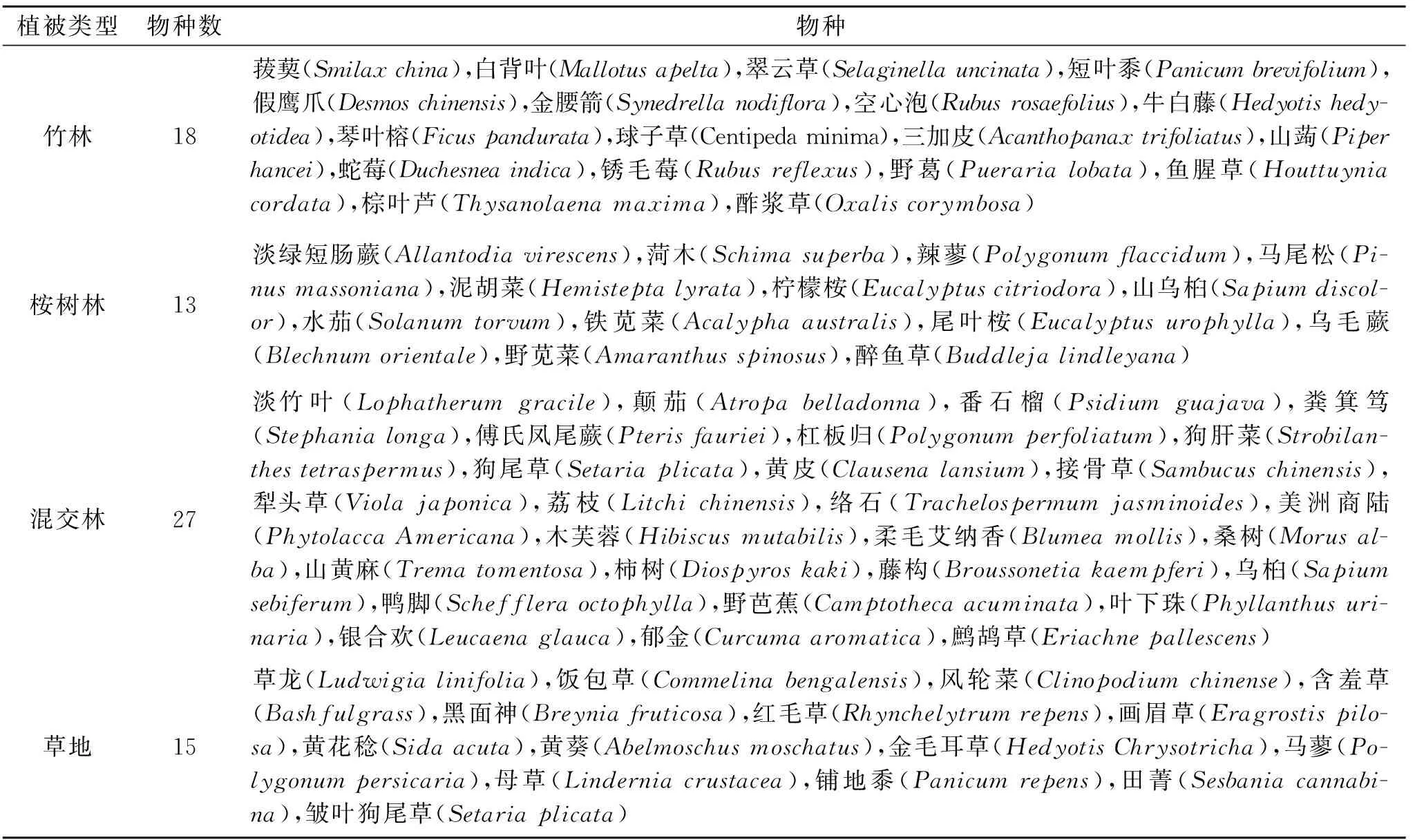
表3 仅在单一植被类型样地出现的物种
3.3 北江河岸带植物物种多样性与环境因子关系
植物物种多样性格局的形成离不开环境因子的作用。表4显示了4种植被类型样地7种环境因子(海拔、至河流距离、土壤含水量、乔木层高度和盖度、草本层高度和盖度)的统计特征,表5和图3分别呈现了北江河岸带物种多样性与环境因子的RDA统计特征与二维排序图。结果表明,物种丰富度指数(S)、均匀度指数(E)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)与乔木层高度、草本层盖度和海拔均呈正相关,与至河流距离、土壤含水量和草本高度呈负相关。即随乔木层高度、草本层盖度和海拔的增加,物种多样性增加,随至河流距离、土壤含水量和草本高度的增加,物种多样性降低。其中,S与至河流距离,E,D,H与乔木层盖度、草本层高度,相关性相对较弱。7个环境因子中海拔因子对河岸带植被的影响最大,其次为土壤含水量,而至河流距离影响最小。乔木层高度和草本层盖度受海拔的影响,海拔越高草本层盖度和乔木层高度就随之升高,反之就会减少。土壤的含水量对草本层高度影响比较大。至河流距离对土壤含水量影响较小,呈较弱的负相关性。
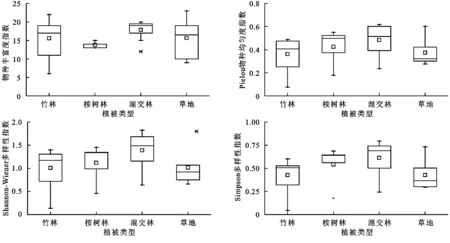
图2不同植被类型植物物种多样性

表4 不同植被类型样地环境因子统计特征

表5 河岸带植物物种多样性与环境因子的RDA统计特征
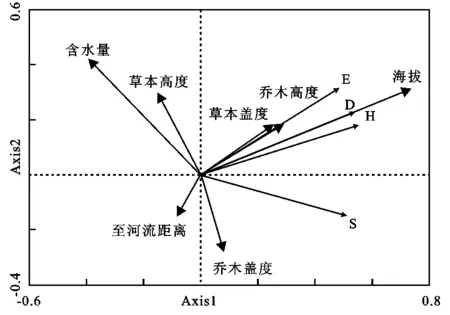
图中字母S代表物丰富度指数;E代表Pielou 均匀度指数;H代表Shannon-Wiener 多样性指数;D代表Simpson多样性指数。
图3北江河岸带植物物种多样性与环境因子关系的二维排序图
4 结论与讨论
在人为干扰景观中,河岸带是半自然生境留存的代表,具有固岸、养分循环、削洪、径流与温度调节、地下水补给、水质净化、为野生动物提供庇护所、为植物繁殖体提供运动与传输的廊道等多种功能[5,13],而其功能又与河岸植被的组成紧密相关[9,23]。但是,河岸带作为人类活动聚集的场所,其植被受人为干扰严重,如加剧的土地利用导致土壤侵蚀与河流泥沙沉积增加、营养输入增加、物理条件(如光照)改变,加之水库蓄水引起的洪水频率和幅度降低为外来植物建植和繁殖扩散创造理想的条件等,显著地改变了河岸原有植被组成[5,23]。因此,针对河岸植物物种多样性展开调查,分析其影响因子,十分必要。
本研究选择北江干流河岸带,开展物种物种多样性调查,共调查到59科116属136种植物。其中,禾本科、菊科、大戟科、蓼科物种数最多,显示了世界广布科对不同环境的适应性。在样地中出现频率最高(>42%)的物种中,空心莲子草是水生型植物,它的生长和大气温度有很大关系,在平均气温21℃左右迅速增长,叶面积急剧扩大;海芋为大型常绿草本植物,喜温暖、潮湿和半阴环境,生长在海拔200~1 100 m热带雨林及野芭蕉林中。物种的分布体现了北江河岸带植被的物种组成特点,北江流域属亚热带季风气候,水热条件好,极其适宜植物的生长,植物群落结构复杂,植株叶大茂盛,不同样地之间植物组成差异明显[19,21]。另外,受地形因子(如坡向、坡度、海拔等)和气候因子(如温度、湿度和降雨等)的影响,植物物种分布状况各异,群落科属组成分散。其中,45.6%的物种只出现在1个样地,分别有13.2%,9.6%,19.9%和11.0%的物种只出现在竹林、桉树林、混交林和草地1种植被类型中,说明河岸带在维持物种多样性的作用更多体现在为不同物种提供栖息地[24],这在不同植被类型间物种多样性分布中体现更为明显。
河岸带栖息地维持较高的物种丰富度可能涉及到各种各样的因素,包括干扰、生产力、便利的繁殖体扩散途径、水陆生态系统界面多样化的物理条件等[24]。有研究表明,生境类型的不同,导致了光照、土壤含水量等环境因子的不同,直接影响到植被类型,从而导致物种多样性产生差异。本研究中物种多样性各指数在不同植被类型上的分布趋势基本一致,但不同植被类型间物种多样性差异并不显著,说明河岸带不同植被类型维持物种多样性的异质性较小。尽管如此,不同植被类型间,桉树林物种丰富度最低,这与桉树对林下物理环境的抑制有关。另外,本研究只分析了不同植被类型间α多样性的差异,并没有讨论不同植被类型间物种的更替(turnover)或者β多样性,但从各植被类型间较高比例的只出现在1种植被类型中的物种数来看,河岸带作为物种多样性的“汇”(species pools)更多体现在物种之间的差异或增加区域物种多样性(γ多样性)方面[24]。因此,与其他栖息地类型相比,河岸带样方平均物种丰富度并不一定高,但相同栖息地内不同样方之间物种的差异方面,河岸带栖息地则高于其他栖息地类型。
河岸带植物物种多样性与环境变量密切相关,如大尺度上的河流形态和小尺度上的生境因子(如海拔、河流方位与梯度、河谷宽度、土壤发育程度等)[14-16,25]。其中,植物物种多样性随海拔高度的变化规律一直是生态学家关注的问题,多海拔梯度下的物种多样性研究具有更强的可比性,可使多样性变化更明显地表现出来[3,11]。海拔对植物群落组成、结构和分布的影响,往往是与其他环境因子作用的共同体现[17,26],因为海拔变化可直接或间接导致其他环境因子产生变化。因此,在研究物种多样性与多种环境因子的相互关系时,海拔因子的存在可能会导致其他环境因子的作用被弱化[17,26]。北江流域多山地、丘陵,海拔变化相对明显,本文结果表明北江河岸带海拔不仅与植物物种多样性有密切关系,亦与其他环境因子关系密切。值得说明的是,除海拔因子外,群落内部环境的异质性同样影响物种的组成和分布,如乔木层高度和草本层盖度都与物种多样性密切相关。从群落结构来说不同层次的物种对群落的贡献是不一样的,一般来说群落结构越复杂,物种多样性就越高。本文调查中混交林物种丰富度指数明显高于其他植被类型,这与混交林群落结构较草地、竹林和桉树林群落结构复杂有关。另外,桉树林和竹林本身的速生植物特性,加上经济利益的驱动,5~6 a就会被轮伐,导致其物种多样性并不高。土壤与植被是一个整体系统,各种因子之间相互联系,共同作用,对于河岸带植被,水分条件是植物生长的重要因素[27]。北江河岸带水分比较充足,土壤的含水量是其植物多样性的重要影响因子。土壤是植物生长的重要物质基础,生长于其中的植物不仅受其理化性质变化的影响,而且对植物群落结构和功能有着重要影响[10,27-28]。本次土壤环境因子只分析了土壤含水量这一因子与河岸带植物多样性的关系,对于土壤各种盐分离子、土壤理化性质等土壤因子与植物多样性的相互作用有待进一步研究。但是,受地理位置、生态系统类型、气候条件、地形条件等多种因素影响,物种多样性与环境因子的关系颇为复杂,呈现的规律差异显著,而其间错综复杂的关系有待更多、更深入的研究。
[1] Pang Z, Huang T, Chen Y. Diminished groundwater recharge and circulation relative to degrading riparian vegetation in the middle Tarim River, Xinjiang Uygur, Western China[J]. Hydrological Processes, 2009,24(2):147-159.
[2] Méndez-Toribio M, Zermeo-Hernández I, Ibarra-Manríquez G. Effect of land use on the structure and diversity of riparian vegetation in the Duero river watershed in Michoacán, Mexico[J]. Plant Ecology, 2014,215(3):285-296.
[3] Schaffers A P. Soil, biomass, and management of semi-natural vegetation: Ⅱ. Factors controlling species diversity[J]. Plant Ecology, 2002,158(2):247-268.
[4] Wang C, Cao G, Wang Q, et al. Changes in plant biomass and species composition of alpine Kobresia meadows along altitudinal gradient on the Qinghai-Tibetan Plateau[J]. Science in China Series C:Life Sciences, 2008,51(1):86-94.
[5] Pausas J G, Austin M P. Patterns of plant species richness in relation to different environments:An appraisal[J]. Journal of Vegetation Science, 2001,12(2):153-166.
[6] Bouman O T. Tree diversity in 30-year chronosequences of cool-humid forests[J]. Ecological Indicators, 2015,49:32-38.
[7] Hector A. Plant Diversity and Productivity Experiments in European Grasslands[J]. Science, 1999,276(5442):1123-1127.
[8] Schwilk D W, Keeley J E, Bond W J. The intermediate disturbance hypothesis does not explain fire and diversity pattern in fynbos[J]. Plant Ecology, 1997,132(1):77-84.
[9] Tornwall B, Sokol E, Skelton J, et al. Trends in Stream Biodiversity Research since the River Continuum Concept[J]. Diversity, 2015,7(1):16-35.
[10] 温璐,董世魁,朱磊,等.环境因子和干扰强度对高寒草甸植物多样性空间分异的影响[J].生态学报,2011,31(7):1844-1854.
[11] 许莎莎,孙国钧,刘慧明,等.黑河河岸植被与环境因子间的相互作用[J].生态学报,2011,31(9):2421-2429.
[12] 黄凯,郭怀成,刘永,等.河岸带生态系统退化机制及其恢复研究进展[J].应用生态学报,2007,18(6):1373-1382.
[13] Capon S J, Chambers L E, Nally R M, et al. Riparian Ecosystems in the 21st Century:Hotspots for Climate Change Adaptation[J]. Ecosystems, 2013,16(3):359-381.
[14] Bruno D, Gutiérrez-Cánovas C, Sánchez-Fernández D, et al. Impacts of environmental filters on functional redundancy in riparian vegetation[J]. Journal of Applied Ecology, 2016,53:846-855.
[15] Li J, Dong S, Yang Z, et al. Effects of cascade hydropower dams on the structure and distribution of riparian and upland vegetation along the middle-lower Lancang-Mekong River[J]. Forest Ecology & Management, 2012,284(3):251-259.
[16] Li J, Dong S, Peng M, et al. Vegetation distribution pattern in the dam areas along middle-low reach of Lancang-Mekong River in Yunnan Province, China[J]. Frontiers of Earth Science, 2012,6(3):283-290.
[17] Bartish I V, Hennekens S, Aidoud A, et al. Species pools along contemporary environmental gradients represent different levels of diversification[J]. Journal of Biogeography, 2010,37(12):2317-2331.
[18] 刘占明,陈子燊.广东北江流域前汛期降水与全球海温及遥相关的相关性研究[J].地理科学,2014,34(10):1239-1246.
[19] 李艳,陈晓宏,王兆礼.人类活动对北江流域径流系列变化的影响初探[J].自然资源学报,2006,21(6):910-915.
[20] 李艳,陈晓宏,张鹏飞.北江流域水文特征变异研究[J].自然资源学报,2013,28(5):822-831.
[21] 梁显强.北江流域综合管理的思考[J].广东水利水电,2012(9):28-31.
[22] Ter Braak C J F, Smilauer P. CANOCO reference manual and CanoDraw for Windows user's guide: software for canonical community ordination (version 4.5)[R]. www. canoco. com, 2002.
[23] You X, Liu J, Zhang L. Ecological modeling of riparian vegetation under disturbances:A review[J]. Ecological Modelling, 2015,318:293-300.
[24] Sabo J L, Sponseller R, Dixon M, et al. Riparian zones increase regional species richness by harboring different, not more, species[J]. Ecology Ecological Society of America, 2008,86(1):56-62.
[25] Kim K D, Park M H, Lee C S. Evaluation of the degraded riparian ecosystems in the geum river watershed in Korea[J]. Journal of Plant Biology, 2012,55(2):132-142.
[26] 赵从举,康慕谊,雷加强.准噶尔盆地典型地段植物群落及其与环境因子的关系[J].生态学报,2011,31(10):2669-2677.
[27] Tererai F, Gaertner M, Jacobs S, et al.Eucalyptuscamaldulensisinvasion in riparian zones reveals few significant effects on soil physico-chemical properties[J]. River Research and Applications, 2015,31(5):590-601.
[28] 赵景学,曲广鹏,多吉顿珠,等.藏北高寒植被群落物种多样性与土壤环境因子的关系[J].干旱区资源与环境,2011,25(6):105-108.
AnalysisofPlantSpeciesDiversityUnderDifferentVegetationTypesinRiparianZoneoftheMainStreamofBeijiangRiver
ZHAO Qinghe1,2, XU Shanshan1,2, MA Lijiao1,2, WU Changsong1,2
(1.KeyLaboratoryofGeospatialTechnologyfortheRegionsofMiddleandLowerReachesofYellowRiver,MinistryofEducation,Kaifeng,He′nan475004,China; 2.CollegeofEnvironmentandPlanning,He′nanUniversity,Kaifeng,He′nan475004,China)
Taking 4 kinds of typical vegetation types such as the bamboo forest, grassland, mixed forest, and eucalyptus forest in the riparian zone along the Beijiang River as the research samples, using the quadrat sampling method and the redundancy analysis (RDA), we conducted the study to investigate the community characteristics and species diversity of riparian vegetation as well as the relationship between plant species diversity and environmental factors. The results showed that, influenced by the tropical monsoon climate which can provide suitable hydrothermal conditions for the growth of plants, 136 species belonging to 116 genuses and 59 families were found in the riparian zone of Beijiang River. With regarding to species diversity of different vegetation types, changes in richness index, Shannon-Wiener diversity index, and evenness index of the riparian plant species were consistent with each other, namely, ranking from high to low in the order of mixed forest, bamboo forest, grassland, and eucalyptus forest. Nevertheless, there was no significant difference among the 4 vegetation types, indicating that there was less heterogeneity in maintaining species diversity between different riparian vegetation types, and then the services of riparian zone in maintaining species diversity mainly was reflected in providing habitats for different species. RDA result showed that, the richness, evenness, and diversity indexes of the plant species were positively correlated with the tree height, herbaceous coverage, and altitude, and were negatively correlated with the distance away from the river and soil moisture content. In general, among the 7 environmental factors we selected, altitude exhibited the greatest influence on the species diversity of the plants in the riparian zone along Beijiang River, and then followed by the soil moisture content, while the influence of the distance away from the river was the least.
riparian zone; species diversity; environmental factors; redundancy analysis; Beijiang Basin
2016-07-30
:2016-08-12
国家自然科学基金项目(41301197);中国博士后科学基金资助项目(2014M550382,2015T80766);河南省高校科技创新团队支持计划(16IRTSTHN012)
赵清贺(1982—),男,河南开封人,博士,讲师,主要从事流域景观格局与生态过程研究。E-mail:zqh410224@126.com
Q142
:A
:1005-3409(2017)05-0215-07
猜你喜欢
林业资源管理(2022年6期)2023-01-18
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
农家科技下旬刊(2019年7期)2019-09-27
生态环境学报(2019年4期)2019-05-31
珠江水运(2019年22期)2019-01-22
珠江水运(2018年22期)2018-12-25
畅谈(2018年11期)2018-08-26
绿色科技(2018年9期)2018-01-29
