
秦皇岛海域野生牙鲆的遗传多样性研究
2017-11-06 20:41任建功王青林宫春光
河北渔业 2017年10期
任建功+王青林+宫春光

摘 要:采用微卫星标记的方法对秦皇岛海域的野生牙鲆进行了遗传多样性分析,结果显示,12对微卫星标记的多态信息含量PIC为0.784~0.949,平均为0.896;12对标记的等位基因数NA为13~108个,平均为51;本研究中观测杂合度Ho为0.286~0.957,平均为0.764,期望杂合度He为0.809~0.953,平均为0906。结果表明,秦皇岛海域野生牙鲆的遗传多样性处于较高水平,是优良的牙鲆种质资源宝库。
关键词:野生牙鲆;微卫星标记;等位基因;遗传多样性
向自然水域投放人工培育的苗种是国内外通行的一种恢复渔业种群资源、改善水域生态环境和增加渔业效益的有效手段[1-2]。《农业部关于做好“十三五”水生生物增殖放流工作的指导意见》指出,“十三五”期间将继续加大对水生生物资源的投入力度,每年增殖重要渔业资源品种的苗种数量达到400亿单位以上。牙鲆是渤海重要的增殖放流鱼类,我国自20世纪80年代在该海域进行牙鲆的增殖放流活动,对天然渔业资源的恢复起到了积极的作用[3]。但是大规模放流人工培育苗种对自然群体遗传多样性的影响受到人们的关注[4]。《农业部办公厅关于进一步加强水生生物经济物种增殖放流苗种管理的通知》(农办渔[2014]55号)明确指出用于繁育增殖放流苗种的亲本应来自该物种原产地天然海域、水产种质资源保护区或省级以上原种场保育的原种。因此,在分子水平阐明野生牙鲆遗传结构,对开展牙鲆增殖放流以及保护牙鲆种质资源具有重要意义。
1 材料与方法
1.1 实验材料
1.1.1 实验鱼
实验鱼为2015-2016年收购的从秦皇岛海域捕获的野生牙鲆(作为放流牙鲆的亲鱼),共计231尾,全长57.73±7.18 cm,体重3 737.55±1 231.55 g,其中雌性109尾,雄性122尾。
1.2 實验方法
1.2.1 样品采集和DNA提取 剪取野生牙鲆胸鳍平铺于采样纸上,37 ℃烘干后保存。使用海洋动物基因组DNA提取试剂盒(TIANGEN,DP324)提取基因组DNA。提取完成后,采用琼脂糖凝胶电泳和紫外分光光度计分别检测DNA的质量和浓度。
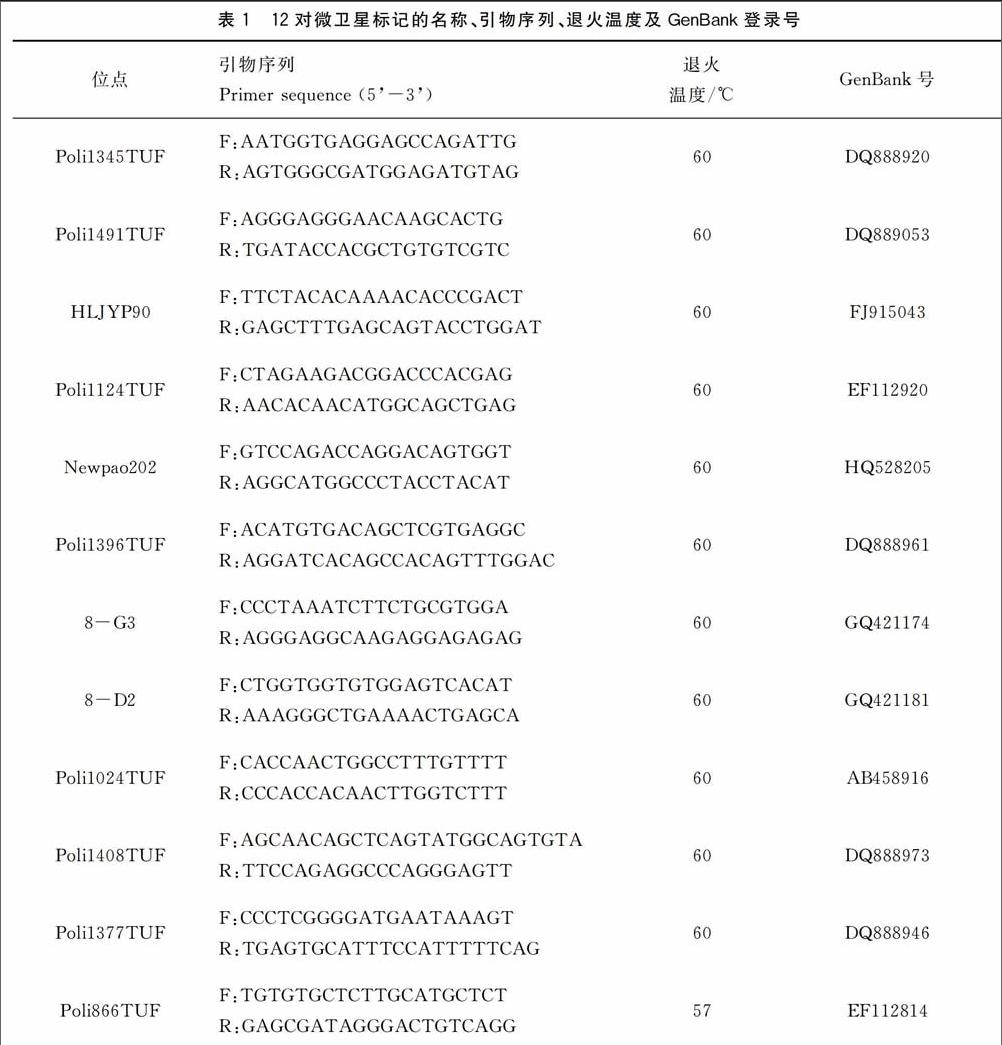
1.2.2 微卫星分析 从马晓冰等(2012)[5]和刘永新等(2013)[6]发表的72对牙鲆微卫星标记中选取12对多态性高的标记用于本研究。各标记的名称、引物序列、退火温度等见表1,引物由上海生工生物工程技术有限公司合成。PCR反应体系为20 μL,包括dH2O 7.4 μL,2×Es Taq MasterMix 10 μL,上下游引物(10 pmol/μL)各0.3 μL,DNA模板2 μL。PCR反应程序为:94 ℃预变性5 min;94 ℃变性30 s,54 ℃复性35 s,72 ℃延伸40 s,共35个循环;最终72 ℃延伸3 min。PCR扩增在PE 9700型PCR仪上进行。将甲酰胺与分子量内标按100∶1的体积比混匀后,取15 μL加入上样板中,再加入1 μL稀释10倍的PCR产物。然后使用3730 XL测序仪进行毛细管电泳,利用Genemarker中的Fragment(Plant)片段分析软件对测序仪得到的原始数据进行分析,将各泳道内分子量内标的位置与各样品峰值的位置进行比较分析,得到片段大小。
1.3 数据分析
利用Genepop 4.0软件分析分型结果,统计各位点等位基因数(NA)、观测杂合度(Ho)、期望杂合度(He)和多态信息含量(PIC),并分析群体的Hardy-Weinberg遗传平衡状态。
2 结果和讨论
2.1 实验结果
12对微卫星标记的遗传多样性参数NA、Ho、He、PIC值见表2。其中,12对微卫星标记的多态信息含量PIC为0.784~0.949,平均为0. 896;12对标记的等位基因数NA为13~108个,平均为51;本研究中观测杂合度Ho为0.286~0.957,平均为0.764,期望杂合度He为0.809~0.953,平均为0.906。
2.2 讨论
多态信息含量(PIC)为衡量标记遗传信息含量高低的主要参数,当PIC>0.5时,表明该遗传标记具有高度的可提供遗传信息性,即高度多态;当0.25 该研究中平均NA为51,远高于马晓冰等[5]和刘永新等[6]对该海域野生牙鲆的研究,说明野生群体的遗传信息含量非常丰富。杂合度包括观测杂合度(Ho)和期望杂合度(He),是反映微卫星标记在群体中遗传变异程度的最优参数[8],是描述遗传多样性的最适参数。杂合度越高,表明群体内遗传多样性就越高,遗传变异程度就越大;反之则群体内遗传变异程度就小[9]。本研究中观测杂合度Ho平均为0.764,期望杂合度He平均为0.906,平均He远高于邵长伟等[10]和马晓冰等[5]研究中的养殖群体和野生群体,说明秦皇岛海域野生牙鲆的遗传多样性处于较高水平。利用野生牙鲆作为放流亲本繁育增殖放流苗种可以改善放流群体的遗传多样性。尽管生产中存在牙鲆亲本对后代贡献率失衡的现象,导致放流群体遗传多样性低于亲本群体[11]。增加繁殖亲本的数量,或者采用生殖细胞移植技术可以有效地解决这个问题[12-13]。 哈迪-温伯格平衡检验结果显示,在12个位点中有8个显著偏离哈迪-温伯格平衡状态(P<0.05)。遗传不平衡现象在已发表的关于牙鲆群体的研究中普遍存在,可能的原因很多,包括捕捞、放流以及海区环境污染造成野生群体等位基因丢失,无效等位基因,样本量较小,性别比例失衡,近交衰退,自然选择等。考虑到偏离哈迪-温伯格平衡的位点的观测杂合度和期望杂合度的差异都比较大,我们推测无效等位基因是引起偏分离的主要原因,但是人为原因导致的环境污染、过度捕捞以及苗种培育过程中的近亲繁殖等因素同样不能忽视。
综上,秦皇岛海域野生牙鲆群体遗传信息含量丰富、遗传多样性水平较高,是优良的牙鲆种质资源宝库,利用野生牙鲆作为放流亲鱼可以提高放流群体的遗传多样性,同时对保护自然种群遗传多样性意义重大。
参考文献:
[1] Seddon PJ,Armstrong DP,Maloney RF.Developing the science of reintroduction biology[J].Conservation Biology,2007,21(2):303-312.
[2] 李繼龙,王国伟,杨文波,等.国外渔业资源增殖放流状况及其对我国的启示[J].中国渔业经济,2009,27(3):111-123.
[3] 金显仕,邱盛尧,柳学周,等.黄渤海渔业资源增殖基础与前景[M].北京:科学出版社,2014,1-407.
[4] Taranger GL,Karlsen ,Bannister RJ,et al.Risk assessment of the environmental impact of Norwegian Atlantic salmon farming[J].ICES Journal of Marine Science:Journal du Conseil,2015,72(3):997-1021.
[5] 马晓冰,王桂兴,刘海金,等.秦皇岛海域野生牙鲆群体遗传多样性分析[J].中国水产科学,2012,19(6):963-969.
[6] 刘永新,朱以美,刘英杰,等.牙鲆渤海自然群体的遗传多样性分析[J].水产学报,2013,37(11):1609-1617.
[7] McDonald GM,Wiley J.Space,Time and Life[M].Seattle:University of Washington Press,2003:109-120.
[8] 刘海金,朱晓琛,孙效文,等.牙鲆5个养殖群体的遗传多样性分析[J].中国水产科学,2008,15(1):30-37.
[9] Nei M,Maruyama T,Chakraborty R.The bottleneck effect and genetic variability in populations[J].Evolution,1975,29(1):1-10
[10] 邵长伟,廖小林,田永胜,等.牙鲆3个养殖群体遗传结构的微卫星分析[J].渔业科学进展,2009,30(1):41-46.
[11] 陈睿毅,王桂兴,刘海金,等.牙鲆亲本对子代贡献率的实验研究[J].中国水产科学,2013,20(4):698-705.
[12] Lacerda S,Costa GMJ,Campos-Junior PHA,et al.Germ cell transplantation as a potential biotechnological approach to fish reproduction[J].Fish Physiology and Biochemistry,2013,39(1):3-11.
[13] Silva MA,Costa GMJ,Lacerda S,et al.Successful xenogeneic germ cell transplantation from Jundia catfish (Rhamdia quelen) into adult Nile tilapia (Oreochromis niloticus) testes[J].General and Comparative Endocrinology,2016,230:48-56.endprint
猜你喜欢
锦州医科大学报(2022年3期)2022-06-06
新课程·下旬(2018年9期)2018-11-14
中国医药导报(2018年14期)2018-08-30
长江蔬菜·学术版(2016年12期)2017-01-12
天津农业科学(2015年5期)2015-05-30
青少年科技博览(中学版)(2015年10期)2015-01-11
中学生物学(2008年6期)2008-08-29
