
模块育种的理论与方法
2017-11-17 02:54施利利王有伟孙宁张欣丁得亮崔晶王松文
天津农学院学报 2017年3期
施利利,王有伟,孙宁,张欣,丁得亮,崔晶,王松文,
模块育种的理论与方法
施利利1,2,王有伟1,孙宁1,张欣1,2,丁得亮1,2,崔晶1,2,王松文1,2,通信作者
(1. 天津农学院农学与资源环境学院,天津300384;2. 天津中日水稻品质食味研究中心,天津300384)
以‘扬稻6号’(‘93-11’)为出发品种,通过设计育种的方法,选育出水稻品种‘93-11Y’,并解析出该品种的5个模块。基于5个模块特征,将籼稻可以划分为两个亚群。在构建遗传群体和育种群体过程中,可以选择粳稻与籼稻亚群2杂交。依据构建遗传模块和育种模块的育种原理可以实施种质鉴定、亲本塑造、杂交组合的策略,这种模块育种的理论与方法可用于新时期育种。
水稻;模块;模块育种;理论
1996年,杨守仁等提出新品种的选育远不像“部件”(性状)之可以组装为“机器”(品种)[1],薛勇彪等提出“分子模块育种”[2]。围绕“模块育种”,一些学者提出了相关的理论与方法。2003年,Peleman Johan D 等提出了设计育种(Breeding by Design)分3步进行:(1)定位所有QTL(基因);(2)鉴定所有等位基因(Allele);(3)设计育种[3]。针对设计育种过程,提出了基于染色体片段代换系群体的育种。在理论上,应对产量、品质等这些复杂性状进行基因组改良。但以染色体片段代换系作为材料,只能是对少数基因(性状)进行遗传操作。2005年,Ashikari M等提出了作物定制改良(Tailor-made crop improvement,TCI)[4],实现了水稻的基因和聚合,培育出矮杆大穗新品种。2017年,曾大力等提出了优化设计育种[5],实现了3个品种、28个基因的聚合,选育出‘RD1’、‘RD2’、‘RD3’3个新品种。另外,还有核心种质育种[6]、超级杂交稻育种等新概念、新方法[7]。围绕模块育种有两种基本观点。一种观点认为是可行的,主要依据是多数农艺性状受多基因控制,并具有“模块化”特性[2]。另一种观点认为,生物是有机体,不能像机器那样,由部件进行组装[1]。本课题组认为,在某些条件下,可以近似于“部件组装成机器”,模块育种是可能的。本研究以‘扬稻6号’(1997年审定的新品种,又名‘93-11’)改良为例,解析了一些遗传模块和育种模块,选育出‘93-11Y’,以期为模块育种的理论探索和育种实践提供依据和参考。
1 材料与方法
1.1 材料
试验材料如表1所示。

表1 试验材料
1.2 方法
1.2.1 KASP标记与生物信息学分析
KASP是竞争性等位基因特异性PCR(Kompetitive Allele Specific PCR)的缩写[7]。设计KASP引物时,关键是设计两个竞争性的正向引物,这两个竞争性引物参与扩增反应的几率相等。因为在KASP标记中要用红蓝荧光分别标记两个竞争性正向引物[8]。所以随着PCR反应的进行,以两种不同的信号(红光、蓝光)自动记录下来。KASP技术的要点是SNP位点的两侧序列已知,3 000个水稻品种的重测序信息为KASP标记检测提供了可能[9]。KASP与SNP标记相比,KASP检测不像SNP那样基于基因芯片,KASP在检测SNP位点的同时还检测了SNP位点上游的多态信息,并实现了高通量。
按水稻基因组2~5 cM间隔,选取600个SNP标记,对51个品种进行基因组扫描和多态性筛选,选出363个多态性标记。363个标记按标记纯合、杂合以不同颜色显示基因型,杂合时以黑色表示。纯合带型以红、粉红等不同颜色标出其带型特征及位置。图示基因型使用Flapjack按照文献[10]进行。
1.2.2 模块鉴定
鉴定模块的基本标准是:(1)以‘93-11Y’(‘93-11’的染色体片段代换系(代换系))为材料,片段不是来自于其亲本‘广陆矮4号’;(2)片段来自于粳稻类型;(3)依据连续的两个标记(代表相应的染色体片段)片段(代表一个染色体片段),在确定3个区域(模块)的基础上,另选择了第1染色体片段(1个比标记片段)和第8染色体片段(1个标记片段)。这样,符合3个条件的模块共有5个模块(遗传模块)。模块育种的第一步是力求建立分布于整个基因组的多基因模块,而在不断更新的模块认知应用中关联于特定的育种计划。
1.2.3 ‘93-11Y’(‘93-11’染色体片段代换系)的选育
以‘津稻108’(‘广陆矮4号’ב东乡野生稻P16’)为基因供体与‘93-11’杂交,获得F1,再与‘93-11’回交,经选择获得‘93-11Y’。
1.2.4 籼稻两个亚群的划分
依据图示基因型选择‘93-11’的5模块(依据1.2.2的3个标准)确定籼稻亚群2,包括‘T001’、‘HC-66’、‘巴西稻’、‘淡绿叶’、‘IPM籼3’、‘褐颖101’、‘58025’、‘矮46’、‘广陆矮4号’等9个品种,其余14个籼稻划分为籼稻亚群1。
2 结果与分析
2.1 图示基因型
将KASP标记(SNP标记)的基因型转变成图示基因型信息,用Flapjack软件做出12个染色体的图示基因型,其中一个样品的第6条染色体的图示基因型如图1所示。
图1表明,样品47(‘93-11Y’)的KASP标记、A004086(第7个标记)、A004088(第8个标记)的带型与样品51(‘广陆矮4号’)不同(不是来源于‘广陆矮4号’),同样,样品7、10、12、45、31、39、40、15都不具有这个带型。而另外的籼稻具有这个带型。说明籼稻可以分为两个亚型,将样品7、10、12、45、31、39、40、15定名为籼稻亚型2,其余籼稻划分为籼稻亚型1,所有粳稻与亚型1亲缘关系远。

图1 第6染色体图示基因型
2.2 模块鉴定与模块特征
2.2.1 模块
模块01-01 位于第1染色体。01代表第1 染色体,-01代表第1 染色体上的第1 个模块。在01-01 模块左侧有基因(多穗粒数基因),右侧有/基因。这样,-01-01-/构成一个育种模块(图2)。

图2 模块01-01
图3表明,遗传模块由位于第6染色体上6 948 457 bp处和8 231 471 bp处的染色体片段组成,育种模块包括左侧的基因(胶稠度)和右侧的基因(抽穗期)。

图3 模块06-01
图4表明,遗传模块由位于第7染色体上的7 651 467~8 123 732 bp的片段组成,育种模块由7 651 467~9 155 185 bp 的片段组成,包括基因(淀粉合成)和基因(单株产量)。
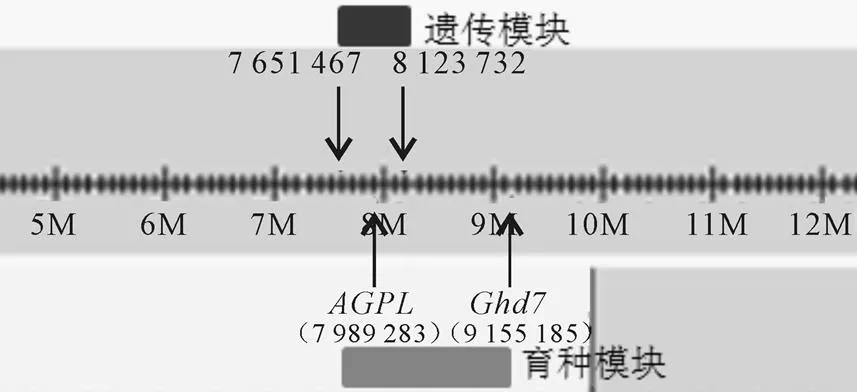
图4 模块07-01
图5表明,遗传模块是位于3 854 233 bp的区段,育种模块包括从3 854 233~5 353 700 bp的区段,包括基因(抽穗期)和基因(淀粉合成)。

图5 模块08-01
由图6可以看出,遗传模块在14 451 520~15 053 026 bp的区域,在距离较远的区域有一个育种模块,包括基因(光敏不育基因)。

图6 模块12-01
品种要求特异性、一致性、稳定性。从长期育种设计考虑,生产上需要多型品种。根据51个品种、363个KASP标记分析结果,本课题组解析了5个育种模块(图2~图6),基于5个模块的基因聚合,结合品种综合改良和5次品种更替趋势,籼稻品种可分为两个亚群(图7),亚群1包括第5代品种(包括最新育品种和新品系)。亚群2包括‘T001’、‘HC-66’、‘巴西稻’、‘淡绿叶’、‘IPM籼3’、‘褐颖101’、‘58025’、‘矮46’、‘广陆矮4号’等籼稻品种及衍生种。如果构建定位群体,则是选择亚群2品种与粳稻杂交。例如,国家水稻基因组计划就是利用与‘广陆矮4号’的近缘系‘矮脚南特’与‘东乡野生稻’(偏粳型野生稻)杂交获得的。同样,日本RGP则使用了同属于籼稻亚群2的籼稻与‘日本晴’(粳稻)杂交获得的后代群体(F2)。如果构建定位群体,则可适当放宽遗传距离,如本课题构建的J107S/Jingdao109(Jia,2001),sh-38/L02[11],如果是粳稻基因定位,多态性下限可设置为18%(未发表资料)。如果是构建育种群体,则更可放宽遗传距离,甚至是近缘系。

图7 籼稻亚群及杂交模式图
2.2.2 品种‘93-11’的模块育种
‘93-11’的改良利用了‘东乡野生稻’(P16),‘东乡野生稻’与位于水稻起源中心的东南亚水稻相比,偏粳。经与‘广陆矮4号’杂交、与‘93-11’复交,再用‘93-11’回交,残留的野生稻基因组片段已经很少。‘93-11Y’(93-11/// 93-11//广陆矮4号/东乡野生稻P16)性状已经与‘93-11’相似。综合性状优良(图8),携带‘东乡野生稻’或‘93-11’的优良基因(QTL),实现了模块育种的育种目标。

图8 ‘93-11Y’(‘93-11’改良系)的农艺性状
2.3 模块育种技术的综合运用
曾大力等采用3个亲本(‘日本晴’、‘93-11’、‘特青’)对28个重要农艺性状基因进行聚合(组装),培育出‘RD1’、‘RD2’、‘RD3’3个新品系[5],产量超过高产亲本‘特青’,品质超过优良亲本‘93-11’。以RD1为例,从供体亲本‘日本晴’中引入、、等重要农艺性状基因,引入‘日本晴’的13个外缘片段;从‘93-11’中引入、等优良基因,引入‘93-11’的7个片段,操作时利用‘特青’遗传背景,引入‘日本晴’和‘93-11’的优良基因和分子模块;用101个微卫星标记。历时5年,选育出‘RD1’、‘RD2’、‘RD3’3个改良系(品种),遗传背景为‘特青’基因组成分,在模块育种方面提供了一个成功例证。本研究以KASP标记作为背景标记和鉴定遗传模块标记,具有高效、低成本、定向性、快周期的特点。本研究解析位于第1、6、7、8、12染色体上的5个遗传模块和育种模块。遗传模块由KASP标记界定(一般位于两个KASP标记之间)。育种模块以重要农艺性状基因所在区域划分,主要考虑品种改良的育种效果(基因作用和遗传背景)。育种模块关联育种信息和性状信息。在育种程序中,综合运用了组织培养(野栽组合胚抢救)、快繁、进代、生态育种(海南周年试验)等技术。在KASP标记体系建立中,运用了51个品种(参照品种),600个KASP标记,一次分子标记辅助选择,实现了模块育种。
3 讨论
3.1 模块定义
模块首先像街区(block),这在DNA分子上比较形象。简单地看作一段DNA双链,有起始位点和终止位点。可能包括基因(基因模块),可能是一个特殊的遗传标记(不同于一般的分子标记,而较多地是至少两个分子标记的一个区间,特殊情形下为一个分子标记)。薛勇彪等更侧重“功能模块”,功能模块与结构模块有所不同,功能模块更侧重功能的实现[2]。
在特定的分子育种计划中,模块应当有可操作性。曾大力等认为来自‘日本晴’和‘93-11’ 中的某些片段具有明显的模块特征,尤其位于染色体7的RM6574 和RM3755 的片段(模块)包含、、3个基因[5],与本研究中的07-01 模块为同一模块。
3.2 模块育种与其他育种方式的兼容
在常规育种中培育出一些大面积推广的品种,如‘汕优63’、‘培矮64S’、‘广占63S’、‘扬稻6号’、‘稻花香2号’、‘越光’等都是超亲类型的选择。‘明恢63’育成以后,以‘明恢63’为亲本,培育出131个恢复系,以这些恢复系为亲本配置出的249个组合通过了国家或省区的审定[12]。实际上,育种专家会自觉地运用这些骨干系的模块。如:日本推广面积前10的优质食味品种都有‘越光’的血缘(即利用‘越光’育成),这说明“优质食味”也有若干“优质食味”模块。一般而言,高产育种采用高产亲本,优质育种采用优质亲本。“食味优先,高产与优质兼顾”的理论[13]对新时期育种有一定的理论价值和育种实践意义。国内外提出的大量育种理论多是兼容的,是在特定条件下的理论侧重和再定义。比如在实施模块育种当中,也进行基因聚合(Gene pyramiding)。在本研究的育种程序中,育种分为3步:(1)种质鉴定;(2)亲本塑造;(3)杂交组合。正常情况下,在亲本塑造过程中进行基因聚合(利用多个有利等位基因)。就育种实践而言,或基于某种理论指导,或遵循于育种家的育种实践和经验,最终都是扬长避短,实现品种或种质创新,进行品种持续改良和阶梯式育种,从一个基准水平(以一个出发品种为基准)达到一个新的水平(选育出一个新的品种,这个品种或增产显著,或产量相当而品质更优)。
[1] 杨守仁,张龙步,陈温福,等. 水稻超高产育种的理论和方法[J]. 中国水稻科学,1996,10(2):115-120.
[2] 薛勇彪,种康,韩斌,等. 开启中国设计育种新篇章——“分子模块设计育种创新体系”战略性先导科技专项进展[J]. 中国科学院院刊,2015,30(3):393-402.
[3] Peleman Johan D,van der Voort Jeroen Rouppe. Breeding by design[J].,2003,8(7):330-334.
[4] Ashikari M,Sakakibara H,Lin S Y. Cytokinin oxidase regulates rice grain production[J].,2005,309(741):741-745.
[5] Zeng D L,Tian Z X,Rao Y C,et al. Rational design of high-yield and superior-quality rice[J].,2017,3:17031.
[6] 周少川,李宏,黄道强,等. 水稻核心种质的育种成效[J].中国水稻科学,2008,22(1):51-56.
[7] 袁隆平. 超级杂交水稻育种研究的进展[J]. 中国稻米, 2008(1):1-3.
[8] Rasheed A,Wen W E,Gao F M, et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat [J].,2016,129(10):1843-1860.
[9] Li Z K,Fu B Y,Cao Y M,et al. The 3 000 rice genomes project [J].,2014(3):7.
[10] Milne L,Shaw P,Stephen G,et al. Flapjack–graphical genotype visualization[J].,2010,26(24):3133-3134.
[11] Jia J H,Zhang D S,Li C Y,et al. Molecular mapping of the reverse thermo-sensitive genic male-sterile gene()in rice[J].,2001,103:607-612.
[12] 谢华安. 汕优63选育理论与实践[M]. 北京:中国农业出版社,2005.
[13] 王有伟,苗燕妮,姜鹏,等. 水稻产量、蛋白质及食味特性的关联研究[J]. 北京:中国农学通报,2017,33(5):1-5.
责任编辑:宗淑萍
Theory and Method of Module Breeding
SHI Li-li1,2,WANG You-wei1, SUN Ning1, ZHANG Xin1,2, DING De-liang1,2, CUI Jing1,2, WANG Song-wen1,2,Corresponding Author
(1. College of Agronomy and Resource Environment, Tianjin Agricultural University, Tianjin 300384, China; 2. China-Japan Joint Center on Palatability and Quality of Rice in Tianjin, Tianjin 300384, China)
The‘93-11Y’was bred by the method of design breeding by raising‘Yangdao6’(‘93-11’). The 5 modules were analyzed. Indica rice can be divided into two subgroups based on the characteristics of the 5 modules. In the process of constructing the genetic population and breeding population, japonica rice and indica rice 2 hybrid. According to the principles of constructing the genetic module and breeding module, we can carry out the strategy of germplasm identification, parental moulding and hybrid combination.
rice; module; breeding by modules; theory
1008-5394(2017)03-0032-05
S336
A
2017-04-10
天津市科技计划项目“基于SNP标记及全基因组选择的水稻设计育种”(16ZXZYNC00110)
施利利(1976-),女,天津市人,高级实验师,硕士,主要从事水稻育种研究。E-mail:dw186001@aliyun.com。王有伟(1989-),男,甘肃武威人,硕士在读,主要从事水稻育种研究。E-mail:641158595@qq.com。施利利和王有伟对本文具有同等贡献,为并列第一作者。
王松文(1958-),男,河北沧州人,教授,主要从事水稻育种研究。E-mail:dw186001@aliyun.com。
猜你喜欢
古今农业(2022年2期)2022-08-15
西南农业学报(2022年5期)2022-06-06
安徽农学通报(2022年8期)2022-05-06
中国医药科学(2022年5期)2022-05-05
中国糖料(2021年3期)2021-07-13
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
江西农业(2018年9期)2018-02-11
中国卫生标准管理(2015年3期)2016-01-14
世界热带农业信息(2014年3期)2014-08-12
