
中心硅藻梅尼小环藻淡水株和海水株亚显微结构的形态学差异
2017-12-27 06:21李长玲陈健冰黄翔鹄
海洋科学 2017年8期
李长玲, 陈健冰, 黄翔鹄
中心硅藻梅尼小环藻淡水株和海水株亚显微结构的形态学差异
李长玲, 陈健冰, 黄翔鹄
(广东海洋大学水产学院, 广东 湛江 524088)
本文利用18S rRNA基因对4株经形态学初步鉴定的梅尼小环藻()进行了分子鉴定。结果显示, 这4株分离自淡水环境(2株)和海水环境(2株)的梅尼小环藻的18S rRNA基因序列基本一致, 并与已经报道的梅尼小环藻(登录号为GQ148712和AY496212)聚为一支, 不同方法所构建的系统树的支持率分别为99%(邻接法)和98%(最大似然法), 表明这4株来自不同环境的藻为同一物种——梅尼小环藻。随后作者利用扫描电镜技术在亚显微水平上对这4株梅尼小环藻进行了观察, 结果显示: 梅尼小环藻淡水株和海水株具有相当不同的亚显微结构特征。生活在海水的梅尼小环藻藻株具有更大的细胞以及更多的中央支持突。此外, 本文还对造成梅尼小环藻种内高水平形态差异的原因以及18S rRNA基因序列是否可以作为小环藻属物种鉴定的分子标记做了分析和讨论。
硅藻; 梅尼小环藻(); 亚显微结构; 扫描电镜; 18S rRNA
梅尼小环藻()隶属于硅藻门(Bacillariophyta), 中心藻纲(Centricae), 圆筛藻目(Stephanodiscales), 圆筛藻科(Stephanodiscaceae), 小环藻属(), 是一种广盐性浮游藻类, 在养殖池塘中较为常见。梅尼小环藻作为初级生产者, 在养殖池塘生态系统的食物链中占据着重要的地位, 同时也是水产养殖生产中水质调控的有益藻类[1]。此外, 它在渔业生产中也非常重要, 是鱼、虾、贝类等经济水产动物优质的饵料[2]。梅尼小环藻还被认为是生产多不饱和脂肪酸和短链饱和脂肪酸的优良藻株, 在生物能源方面具有一定的开发潜力[3]。本文利用18S rRNA基因序列对4株经形态学初步鉴定的, 来自淡水和海水的梅尼小环藻进行了分子鉴定, 并用扫描电镜技术对这4株梅尼小环藻进行了亚显微结构的观察, 首次在亚显微水平上对生长在不同环境下的梅尼小环藻进行了形态学差异比较研究, 以期为梅尼小环藻进一步的开发研究提供形态学数据参考。
1 材料和方法
1.1 藻种来源和培养
淡水梅尼小环藻藻株FACHB-1030、FACHB-1661分离自武汉东湖, 由中国科学院淡水藻种库提供(Freshwater Algae Culture Collection at the Institute of Hydrobiology, Wuhan, China, FACHB)。海水梅尼小环藻藻株GDOU1和GDOU2分离自湛江对虾养殖池塘。
藻种用 250 mL三角瓶中培养, 培养光照度为2 000 lx, 光照周期12L︰12D, 培养温度为22℃。其中, 藻株FACHB-1661、FACHB-1030培养在DM培养基里[4], 藻株GDOU1、GDOU2培养在改良后的f/2海水培养基里[5]。
1.2 DNA提取, PCR扩增和测序
总基因组DNA用植物DNA试剂盒(Qiagen, Germany)进行提取。18S rRNA基因序列的扩增用Taq PCR混合试剂盒进行。反应体系为25 μL: 12.5 μL Taq mix 2×酶; 上下游引物各1.0 μL (引物保存浓度10 μmol/L); DNA模版1.0 μL; 最后加9.5 μL ddH2O定容到25 μL。PCR循环扩增在iCycler(Bio-Rad, Hercules, CA, USA)中进行。PCR反应条件为: 94℃预变性4 min, 94℃变性30 s, 57℃退火30 s, 72℃延伸2 min共35个循环, 再72℃延伸10 min。
PCR扩增产物用2%的琼脂糖凝胶电泳检测, EB染色, 凝胶成像系统记录实验结果。随后用UNIQ-10 DNA纯化试剂盒, 根据使用说明的实验步骤进行胶纯化回收(Sangon, China), 并对纯化后的PCR扩增产物进行克隆, 克隆成功后送至生工生物工程(上海)股份有限公司进行测序。
1.3 系统学分析
将测序成功获得的序列上传到美国国立生物信息中心(the National Center for Biotechnology Information, NCBI, http: //www.ncbi.nlm.nih.gov/)网站上, 应用Blast搜索工具(Basic Local Alignment Search Tool, BLAST)对实验获得的基因序列进行 Blast相似性比对分析, 确定目标片段是否正确, 同时进行同源性检测。
根据 Blast 比对结果, 从GenBank中下载包括外群在内的其他相关的基因序列用于系统树构建。用Clustal X 1.83[6]进行比对, 比对后的序列用MEGA 5.0软件手动切掉两端不整齐的冗余序列, 然后进行系统发育分析, 并采用邻接法(Neighbor Joining, NJ)和最大似然法(Maximum Likelihood, ML)构建系统进化树, 计算bootstrap值, 重复1 000次[7]。
1.4 电镜样品制备及拍摄
取50 mL左右对数生长期的新鲜小环藻样品, 3 000 r/min离心10 min, 浓缩成藻泥。硅藻壳的制备是在离心浓缩后的样品中加入同等体积97%的浓硫酸, 放60℃水浴加热10~20 min, 直至原生质被除掉。之后用蒸馏水清洗5次, 将样品洗至中性。干燥后镀金, 用中国农业科学院的Hitachi S-4800扫描电子显微镜观察, 拍照。
2 结果
2.1 基于18S rRNA基因的系统树构建
利用最大似然法和邻接法构建的18S rRNA基因序列的系统树具有相似的拓扑结构, 本文仅显示了以最大似然法构建系统树的拓扑结构, 如图1所示。系统进化树结果显示: 本次实验所采用的4株梅尼小环藻与已经报道过的梅尼小环藻[8-9](登录号为GQ148712、AY496212)聚为一支, 其中邻接法构建的系统树支持率为99%, 最大似然法构建的系统树支持率为98%。表明这4株来自不同环境的藻为同一物种——梅尼小环藻。
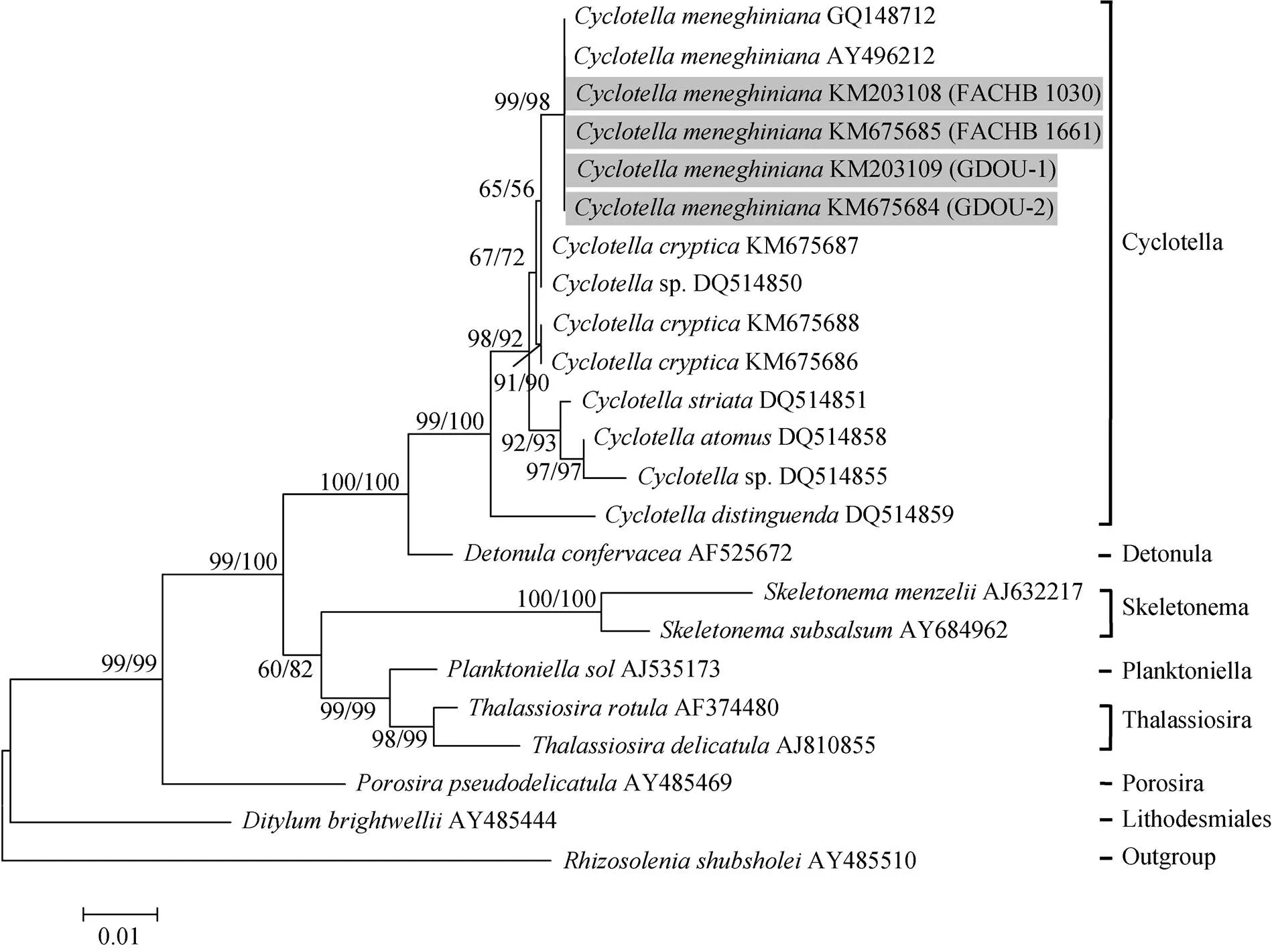
图1 基于18S rRNA基因序列的ML系统树
内部节点上显示重复1 000次计算的自展支持率, 分别由邻接法和最大似然法产生。分支的长度代表进化距离, 用0.01的标尺显示。具有加粗字体和灰色背景的序列为本研究获得的新序列
Bootstrap values are shown at the internal nodes for neighbor joining (NJ: 1 000 replications) and maximum likelihood (ML: 1 000 replications), respectively. Branch lengths correspond to evolutionary distances. A distance of 0.01 is indicated by the scale. The new sequences obtained in our study are bold and shaded in gray
2.2 细胞亚显微结构观察
2.2.1 梅尼小环藻淡水株的亚显微结构
生活在淡水的梅尼小环藻, 直径范围在5~25 μm。从小环藻样品的外观来看(图2中的1和3), 壳面的中央区或多或少有点起伏, 中央区大小约占壳面直径的0.4~0.6。壳面中央区不规则地分布着小的硅质颗粒(Siliceous granules, SG)。边缘区域条纹和非条纹区间隔分布, 围绕着中央区, 每5 μm有5~8个条纹。每个条纹包含四行网纹从中央区与边缘区之间的过渡区域开始逐渐扩大到七行延伸到壳套处。在壳面和壳套接合处的非条纹区存在着刺状突起(Spine, S)。从内部壳面观来看(图2中的2和4), 一般情况壳面只有一个中央支持突(Central fultoportula, cFP), 围着3个围孔。在边缘区, 有6~13个壳缘支持突(Marginal fultoportula, mFP)围绕着2个围孔, 位于每一个或者每两个肋脉之间。单一的唇形突(Rimoportula, RP)在向上方向形成一个长茎和唇形开口。

图2 梅尼小环藻淡水株的亚显微结构
外壳面观. 1和3; 内壳面观. 2和4; 1和2的比例尺为3 μm, 3的比例尺为5 μm, 4的比例尺为4 μm; SP =刺状突起; SG =硅质颗粒; cFP =中央支持突; mFP =壳缘支持突; RP =唇形突
Exterior of the valves: 1 and 3; Internal views of the valves: 2 and 4. The scale bar: Fig 1 and 2 is 3 μm, Fig 3 is 5 μm, Fig 4 is 4 μm; SP = Spine; SG = Siliceous granules; cFP = central fultoportula; mFP = marginal fultoportula; RP = rimoportula
2.2.2 梅尼小环藻海水株的亚显微结构
生活在海水的梅尼小环藻常以单细胞存在, 很少形成短链(如2~3个细胞), 直径为12~30 μm。在扫描电子显微镜下, 从外壳面观来看(图3中的5和7), 壳面被分成两个完全不同的区域, 中央区域几乎平整, 可见三开口的中央支持突, 中央区占壳面半径的0.3~0.5。壳面边缘区具有相等长度的条纹, 4行网隙从壳面的中央区扩增为6行延伸到壳套, 每5 μm 有4~6个条纹。在壳面和壳套接合处的非条纹区可以发现有刺状突起。从内壳面观来看(图3中的6和8), 壳面是平滑的, 每个壳面有3~14个中央支持突和16~23个壳缘支持突, 所有这些支持突都向外开口, 并且每一个支持突都围绕着3个围孔, 具有唇形开口的唇形突, 但并不明显。

图3 梅尼小环藻海水株的亚显微结构
外壳面观. 1和3图; 内壳面观. 2和4图; 3图比例尺为10 μm, 其他图的比例尺为5 μm; SP = 刺状突起; SG =硅质颗粒; cFP =中央支持突; mFP =壳缘支持突
Exterior of the valves: 1 and 3; Internal views of the valves: 2 and 4. Except the scale bar of Fig 3 is 10 μm, others are 5 μm; SP = Spine; SG = Siliceous granules; cFP = central fultoportula; mFP = marginal fultoportula
2.2.3 梅尼小环藻淡水株与海水株的形态学比较
成熟的梅尼小环藻的海水株与淡水株在形态学上有着明显的不同, 见表1。其中, 海水株的直径略大于淡水株。每5 μm的条纹数目, 海水株有4~6个, 而淡水株有5~8个。海水株边缘支持突的个数要远多于淡水株, 可达16~23个, 一般每一个或两个非条纹区就有一个边缘支持突; 而淡水株的边缘支持突数为6~13个, 一般每两个或三个非条纹区才有一个边缘支持突。除此之外, 在边缘支持突的围孔数上两者也存在着差异, 海水株边缘支持突的围孔数一般为3个; 而淡水株一般为3个, 但也有2个围孔数的边缘支持突被观察到。海水株在每个壳面中央支持突的个数也比淡水株多, 一般在2~14个左右, 而淡水株仅有1个, 但两者中央支持突的围孔数相同, 每个中央支持突的围孔数都为3个。两者都存在一个唇形突, 但海水株的唇形突不明显。此外, 两者在壳面和壳套接合处的非条纹区都存在刺状突起, 在壳面中央区也都不规则地分布着硅质颗粒。
表1 梅尼小环藻淡水株与海水株的亚显微结构特征比较

Tab.1 Comparison of the ultrastructures of freshwater and seawater strains of C. meneghiniana
3 讨论
硅藻的祖先来自海洋, 但很多时候它们独立地生活在淡水中, 盐度被认为是硅藻分布的一个屏障。然而, Alverson[10]的研究结果显示盐度屏障也没有作者先前想象的那么强大, 例如,已经被推测作为一个大枝系, 在海洋, 半咸水和淡水都有生存的物种。海水-淡水转变的方向目前还不清楚, 在历史事件中, 藻株在海水和淡水之间转变成功的例子非常少, 而且繁殖后代是不可逆的。在淡水和海水中, 或者在这两个中间的栖息地都能发现小环藻物种, 因此, 小环藻被称为“广盐性”物种, 以及预测它可能代表为数不多由海洋栖息地到淡水转变的分类群的例子[11]。
梅尼小环藻是“广盐性”硅藻物种中最常见的一种[12], 其形态特征已经被很多文献报道过[13-17], 但也有将隐秘小环藻()在形态学观察下错误的推断为梅尼小环藻的报道[18]。由此可见, 形态学特征并不总是小环藻界定物种和系统发育关系的可靠指标, 如果没有分子水平上的研究, 仅仅依据显著的形态学差异来鉴定物种, 那么来自淡水和海水的梅尼小环藻很有可能会被错误的鉴定为不同的物种。利用分子生物技术可以提高作者在硅藻物种的分类和界定研究中发现和提取细微差异的能力[19-20], 因此, 作者对4株经形态学初步鉴定为梅尼小环藻的藻株进行了以18SrRNA基因为分子标记的分子鉴定, 用以确定所研究的藻株为梅尼小环藻无误, 并在此鉴定的结果上进行形态学上的比较研究, 这样则更有利于人们推断不同环境下造成同一种藻不同形态的原因。
作者研究结果显示, 来自不同环境下的4株梅尼小环藻有着几乎完全一致的18S rRNA基因序列, 但却在亚显微结构上存在较大的差异。这一结果与已经报道的另外一个硅藻物种的情况相似, 而被认为是同一物种的不同生态表型[21]。除此之外, 物种适应不同环境条件的能力与细胞形态多变的特征有关, 如Borowitzka等[22]指出,中空泡结构的不同可能是其生活在特定条件的适应能力的一个重要因素。因此, 在不同环境下(淡水和海水)梅尼小环藻具有不同的形态结构的主要原因作者更倾向于是物种自身对环境条件的一种形态学响应。目前, 作者还无法得知它是如何做出响应的。而生活在海水的梅尼小环藻藻株具有更大的细胞以及更多的中央支持突, 可能是其生存在高盐度、不稳定环境的适应能力的一个重要因素。这些研究结果将会帮助作者更加准确地区分生活在不同环境下, 形态不同的同一物种。另一方面, 有研究指出, 18S rRNA基因的DNA序列处于中度保守, 更适合属水平的系统发育研究[23]。从本研究的结果来看, 来源于不同区域的梅尼小环藻聚为一支, 并与其他小环藻属的物种有着明显的差异, 如图1所示, 说明18S rRNA基因序列在小环藻属物种间存在明显的遗传差异, 有潜力作为小环藻属物种鉴定的分子标记。但是人们同时也看到, 针对于来自不同环境或区域的梅尼小环藻, 种内不同株系之间的18S rRNA基因序列遗传差异非常小, 因此, 18S rRNA基因序列在种内不同株系之间显得过于保守, 可能不适合作为小环藻种内不同株系物种鉴定的分子标记。
[1] 黄光华, 江林源, 卢小花, 等. 小环藻在罗氏沼虾人工育苗中的应用研究[J]. 安徽农业科学, 2011, 39(13): 7849-7850. Huang Guanghua, Jiang LinYuan, Lu Xiaohua, et al. Study on the Application ofsp. In the Artificial Breeding of[J]. Journal of Anhui Agri. Sci., 2011, 39(13): 7849-7850.
[2] 杨海明, 尹邵武, 吴朝晖, 等. 梅尼小环藻(Kiits)的培养与利用[J]. 湖南师范大学: 自然科学学报, 1997, 20(3): 56-61. Yang Haining, Yin Shaowu, Wu Zhaohui, et al. On Artifical Cultivation and Utilization of[J]. Acta Sci Nat Univ Nom Hunan, 1997, 20(3): 56-61.
[3] 冀颐之, 刘杨平, 黄迎春. 梅尼小环藻产油培养工艺的优化[J]. 安徽农业科学, 2012, 40(11): 6360-6362. Ji Yizhi, Liu Yangping, Huang Yingchun. The Optimization of Culture Conditions On Lipid Produced by[J]. Journal of Anhui Agri. Sci., 2012, 40(11): 6360-6362.
[4] Beakes G W, Canter H M, Jaworski G H M. Zoospore ultrastructure ofand, two chytrids parasitizing the diatom[J]. Canadian Journal of Botany, 1988, 66(6): 1054-1067.
[5] Guillard R R. Culture of phytoplankton for feeding marine invertebrates [M]. Culture of Marine Invertebrate Animals, 1975: 29-60.
[6] Thompson J D, Gibson T J, Plewniak F, et al. The CLUSTAL X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools[J]. Nucleic Acids Research, 1997, 25(24): 4876- 4882.
[7] Tamura K, Peterson D, Peterson N, et al. MEGA 5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 2011, 28(10): 2731-2739.
[8] Jung S W, Han M S, Ki J S. Molecular genetics divergence of the centric diatomand(Bacillariophyceae) revealed by nuclear ribosomal DNA comparisons [J]. J Appl Phycol, 2010, 22(3): 319- 329.
[9] Beszteri B, Acs E, Medlin L K. Ribosomal DNA sequence variation among sympatric strains of thecomplex (Bacillariophyceae) reveals cryptic diversity[J]. Protist, 2005, 156(3): 317-333.
[10] Alverson A J, Jansen R K, Theriot E C. Bridging the Rubicon: phylogenetic analysis reveals repeated colonizations of marine and fresh waters by thalassiosiroid diatoms[J]. Molecular Phylogenetics and Evolution, 2007, 45(1): 193-210.
[11] Quijano-Scheggia S I, Garc S E, Lundholm N, et al. Morphology, physiology, molecular phylogeny and sexual compatibility of the crypticcomplex (Bacillariophyta), including the description ofsp. nov[J]. Phycologia, 2009, 48(6): 492-509.
[12] Finlay B J, Monaghan E B. Maberly S C. Hypothesis: the rate and scale of dispersal of freshwater diatom species is a function of their global abundance[J]. Protist, 2002, 153: 261-273.
[13] Mann D G, Droop S J M. (1996) Biodiversity, biogeography and conservation of diatoms[M]//Kristiansen J. Biogeography of Freshwater Algae. Developments in Hydrobiology, Dordrecht: Springer, 1996: 118: 19-32.
[14] Round F E, Crawford R M, Mann D G. The diatoms: biology & morphology of thegenera [M]. Cambridge University Press, 1990.
[15] Cho K J. Fine morphology of some Cyclotella species from the freshwater zone of the Naktong River [J]. Algae, 1996, 11: 9-21.
[16] 胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. Hu Hongjun, Wei Yinxin. The Freshwarter Algae of China——Systematic, Taxonomy and Ecology[M]. Beijing: Science Press, 2006.
[17] 郭玉洁, 钱树本. 中国海藻志第五卷硅藻门第一册中心纲[M]. 北京: 科学出版社, 2003: 1-493. Guo Yujie, Qian Shuben. Marine Phycology of China——The Fifth Volume——Bacillariophyta—— The Fisrth Volume——Centricae[M]. Beijing: Science Press, 2003: 1-493.
[18] Mclachlan J. The growth of unicellular algae in artificial and enriched sea water media[J]. Canadian Journal of Microbiology, 1959, 5(1): 9-15.
[19] Sorhannus U, Fox M G. Phylogenetic analyses of a combined data set suggest that the Attheya lineage is the closest living relative of the pennate diatoms (Bacillariophyceae)[J]. Protist, 2012, 163(2): 252-62.
[20] Stepanek J G, Kociolek J P. Molecular phylogeny of(Bacillariophyta): An investigation into the monophyly and classification of the amphoroid diatoms[J]. Protist, 2014, 165(2): 177-195.
[21] Martino A D, Meichenin A, Shi J, et al. Genetic and phenotypic characterization of(Bacillariophyceae) accessions 1[J]. Journal of Phycology, 2007, 43(5): 992-1009.
[22] Borowitzka M A, Volcani B E. The polymorphic diatom Phaeodactylum tricornutum: ultrastructure of ITS morphotypes 1, 2[J]. Journal of Phycology, 1978, 14(1): 10-21.
[23] Coleman A W. ITS2 is a double-edged tool for eukaryote evolutionary comparisons[J]. TRENDS in Genetics, 2003, 19(7): 370-375.
(本文编辑: 梁德海)
Ultrastructure of the centric diatom: morphological differences between freshwater and seawater strains
LI Chang-ling, CHEN Jian-bing, HUANG Xiang-hu
(Fisheries College of Guangdong Ocean University, Zhanjiang 524088, China)
In this paper, four strainswere identified using 18S rRNA gene sequences. The results show that the 18S rRNA gene sequences of these four strains (two come from freshwater and two from seawater) are very similar and that these four strains are closely related to(GQ148712 and AY496212) with high support (99/98 for neighbor joining/maximum likelihood). According to the molecular results, the four strains are of the same species, i.e.,. Then, the ultrastructure of the four strains was studied using scanning electron microscopy (SEM). The results show that the ultrastructure of freshwateris different from that of seawater, which also have larger cells and contain higher amounts of central fultoportula. In addition, this paper analyzed the causes of high-level morphology differences withinspecies and discussed whether the 18S rRNA gene can be used as a molecular marker for the identification ofspecies.
diatom;; ultrastructure; scanning electron microscopy; 18S rRNA
[Marine Fishery Science and Technology and Industry Development Project of Guangdong Province, No.A201508B08]
Dec. 9, 2016
Q179.1
A
1000-3096(2017)08-0116-06
10.11759/hykx20161023002
2016-12-09;
2017-04-01
广东省海洋渔业科技与产业发展专项(A201508B08)
李长玲(1964-), 女, 河南焦作人, 教授, 硕士, 主要从事水域生态学研究, 电话: 0759-2396044, E-mail: ybcl901@126.com; 黄翔鹄, 通信作者, 教授, 博士, 电话: 0759-2396044, E-mail: hxh166@126.com
猜你喜欢
当代水产(2022年3期)2022-04-26
小哥白尼(野生动物)(2020年2期)2020-06-16
学生天地(2020年25期)2020-06-01
趣味(数学)(2019年12期)2019-04-13
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
作文周刊·小学一年级版(2016年39期)2017-03-03
中国继续医学教育(2015年2期)2016-01-06
少儿科学周刊·儿童版(2015年11期)2015-12-17
振动、测试与诊断(2014年6期)2014-03-01
现代检验医学杂志(2014年1期)2014-02-06
