
沙蒿对局部沙埋的反应及其克隆整合的意义
2018-02-02 07:39童跃伟吴春玲
安徽农业科学 2018年4期
童跃伟,吴春玲,彭 政,周 欣
(景德镇市林业科学研究所, 江西景德镇 333000)
20世纪80年代以来,植物的克隆生长(clonal growth)及其生态效应的研究受到植物生态学家们越来越多的关注[1-9]。目前,已被提出的克隆植物对策包括基于克隆构型(clonal archetecture)可塑性(plasticity)变化的躲避反应[10]和基于生理整合(physiological integration)的忍耐反应[11],以及相联分株间通过劳动分工(division of labour)对资源的利用[12-13]等。克隆生长一个重要特征就是克隆植物的分株之间通过生理连接(匍匐茎、根茎等)而相互传送光合产物、水分和矿质养分等资源。克隆植物的这种行为被称为生理整合或者克隆整合。克隆基株常常受益于这种生理整合[14-17],可使整个克隆在高度异质性生境中生存下来。克隆植物的这种行为不仅可以使克隆分株在一定意义上逃避不利的环境条件,而且可以使之充分利用有利的资源条件[15],因而是植物克隆植物一个比较突出的优势性状[18]。一些实验室和野外研究表明,处于生理整合的克隆分株比相互分离的分株具有明显的生长优势[9,16,19-20]。另一方面,克隆整合可使克隆植物忍耐处于物理性干扰、来自其他植物个体的竞争压力、植食动物的去叶干扰、沙埋等[21-27]。然而,对于不同生境或不同物种,其克隆整合的重要性也有较大差异,因此更多地关注和研究这种变异产生的原因,有利于更好地理解在自然界克隆整合的意义[9]。
在内陆干旱沙丘环境中,不同程度的沙埋是植物经常遭遇的事件。沙埋可以改变植物所处的环境条件。如可以改变植物的湿度、温度、根部的氧气含量、土壤有机物含量等,从而影响植物生长和存活能力。由于沙丘植被覆盖率和风速的差异,植物所受沙埋的程度也不同。轻微程度的沙埋可以通过排除没有适应新环境能力的植物,从而增加植物根系周围的湿度和空间,改善植物的生长环境。然而,沙埋超过一定程度后,就会影响植物的生长及其存活[28]。不同植物所能忍受的沙埋程度不同。沙埋使得长期生长在沙丘生境中的植物在进化过程中拥有了各种忍受沙埋的能力。在沙丘环境中生长的许多克隆植物,常常是沙地的“先锋植物”。因为克隆整合作用的存在,可能使克隆植物对沙埋具有比一般植物更大的忍受能力[29]。对这些有固沙作用及应用价值的克隆植物的忍受沙埋能力进行研究,对于帮助干旱地区环境恢复及制止土地大面积沙化具有重要作用。因此,笔者对沙丘常见的一种草本克隆植物——沙蒿在沙埋状况下的存活、生长、生根以及克隆整合的作用进行了研究,旨在为促进植物生长及加快沙丘固定提供借鉴。
1 材料与方法
1.1研究地概况研究地点位于浑善达克沙地南端,内蒙古自治区中部的多伦县境内。多伦县地理坐标为115°50′~116°55′ E,41°46′~42°39′ N。位于中温带,属于半干旱半湿润大陆性气候。春季干旱多风,夏季湿热多雨,雨热同期,秋季凉爽短促,冬季寒冷漫长。年平均降水量385.5 mm,年平均温度1.6 ℃,年平均日照时数3 400 h。该地区为低山沙丘,波状起伏。海拔高度为1 200~1 300 m,最大相对高差165.5 m。植被以豆科、菊科、禾本科、毛茛科为主。
1.2研究物种沙蒿(Artemisiasphaerocephala)属菊科、蒿属的沙生半灌木,株部50~80 cm,且主根粗壮,深可达2 m,侧根发达而密集,多分布在50~60 cm的沙层内。沙蒿为喜沙性植物,主要生境为固定、半固定沙地。其生态幅宽,分布面积广,从内蒙古东部的科尔沁沙地到我国西部的新疆均有分布,在沙区人工植被中可天然更新。沙蒿具有抗旱、耐沙埋、耐贫瘠的特点。当沙埋不过顶时,枝条叶腋芽可产生不定根萌发出新枝条,增加根系吸收土壤水分与养分的功能,促进地上部植株的生长,增加固沙能力。沙蒿春季萌发早,出苗快,分枝多,生命力强。9月中下旬种子成熟,10月底停止营养生长。种子黑色或黑绿色,多吸水后产生黏稠状液体,起到固定种子周围土壤或沙粒的作用,因而轻微风蚀对其生长影响不大。
沙蒿枝叶为许多家畜所采食,尤其为骆驼所喜食,瘦果可食用。沙蒿也是我国干旱地区优良的固沙植物种,也是流动沙丘、半流动沙丘、固定沙丘分布最重要的先锋固沙植物之一,可以防风抗蚀,对于维持荒漠生态系统的稳定性具有特殊意义。
沙蒿的直立茎在风的作用下平卧于沙地上,尤其在被沙埋以后,会从茎节处向地面生出不定根,而在此节上的叶或分枝继续向上生长,这样就与地面以下的不定根一起构成了一个完整的在形态和生理上都相对独立的个体,这时如果将其与母体分离开来,它也能独立生活下来,这也是克隆性的表现,这样形成的独立个体也可以视为一个克隆分株。
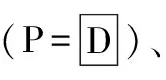
沙蒿在自然条件下生长30 d后,于2011年8月24日采集样本。将每个样本的近端和远端分别采集,记录存活状态、分株数,并测量株高及根长,进行记录。再将采取的样本分为地上绿部分、地上枯部分、茎杆、根,分装在信封中并做好标记。把所有的信封在烘箱中烘干48 h至恒重,确定各部分干重进行分析。
2 结果与分析
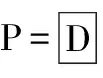
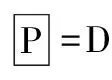
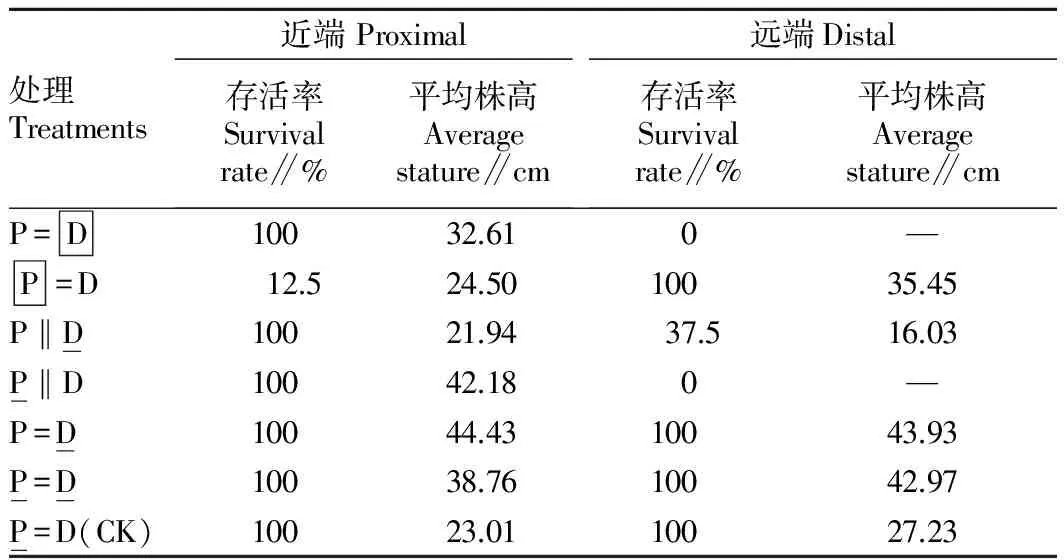
表1 沙蒿在沙埋处理下的存活情况
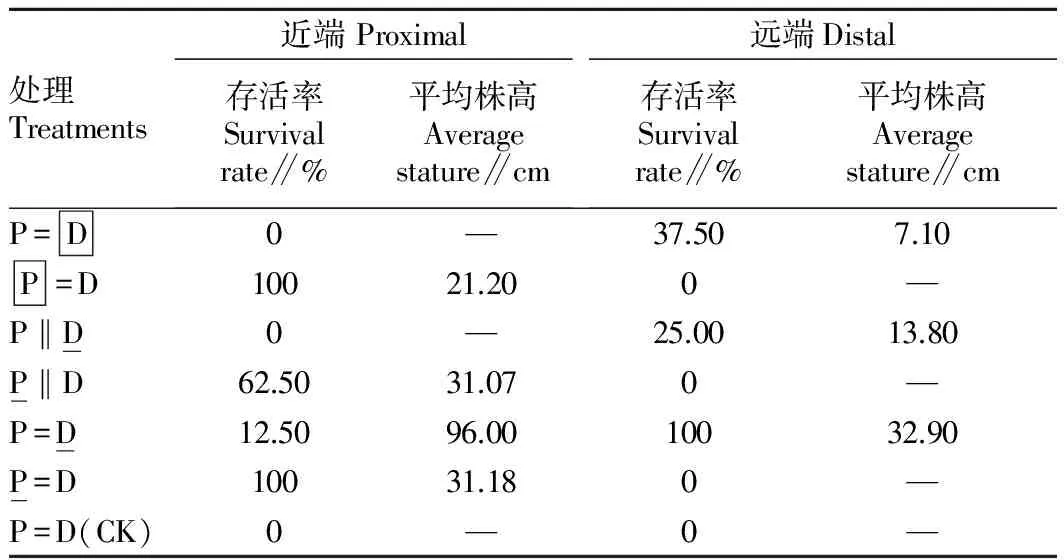
表2 在同沙埋处理下沙蒿的生根情况
2.4地上绿色生物量的增长100%沙埋处理下(P=D和P=D),即使保持茎的连接,也是被沙埋的部分没有生物量或极少(全部死亡或多数死亡),而未被沙埋的部分生物量与CK差异不明显。在切断近端与远端茎连接的情况下,50%沙埋远端时远端绿色生物量较少,这是由于部分死亡所致,而50%沙埋近端时,远端生物量为0,原因是此时远端全部死亡。在50%沙埋但同时保持茎的连接的情况下,无论近端和远端,地上绿色生物量都较大,尤其是远端50%沙埋时,地上绿色生物量明显高于CK(图1)。

图1 不同沙埋处理下沙蒿的地上绿色生物量Fig.1 Aboveground green biomass of Artemisia shpaerocephala under different sand burial treatments
3 结论与讨论
对于远端的存活来说,保持茎的连接是重要的。但是在切断茎联系的情况下,如果将远端进行部分(50%)沙埋,由于沙蒿的克隆性而有可能生根固定并生存下来,但100%沙埋由于完全限制了其光合能力导致死亡。无论近端或远端,能否生根与是否沙埋有关。近端即使被全部沙埋,可能地上部分死亡,但会生出很多不定根,在沙生干旱的生境中起到加强水分吸收的作用。
对比P‖D 和 P=D 处理中远端的生根情况,后者要比前者好得多,这主要是由于茎的连接状况不同。后者是保持茎连接的,近端会输送水分、养分、光合产物等给远端,以满足远端的需求,使之迅速生根。而前者切断了茎的连接,近端难以将物质输送给远端,远端只能依靠自身的贮备和光合作用产物以供其生根之需,这样生根自然会受到很大影响。这充分说明克隆整合对于克隆植物的重要性,尤其是在经常遭受干扰的环境中。如果将P‖D 和 P=D 2种处理中近端的生根情况进行对比,后者明显好于前者,这也是出于相似的原因,即保持茎连接有利于发挥克隆整合的作用。从这2种处理的对比来看,在保持茎的连接的情况下(P=D)的克隆整合是从远端向近端输送物质的。因此,对于沙蒿而言,克隆整合可以是双向的,即水分或养分等物质可以从近端输往远端,也可以从远端输往近端,关键是决定于哪个部分受到干扰或损伤。
在保持茎连接和50%沙埋情况下,沙埋远端比沙埋近端的生物量积累明显增多,可能是由于远端全部生根,而从土壤中吸收尽可能多的水分和养分,除了供给远端本身以外还通过克隆整合作用供给近端。因此,克隆整合可以缓解沙埋对沙蒿以及类似克隆植物存活的不利影响,是克隆植物对异质性沙化环境的生态对策之一[30]。基于克隆整合对克隆植物忍受沙埋能力的贡献,某些克隆植物可作为干旱和半干旱沙化环境中植被恢复的优良固沙物种。
[1] KROON H D,KNOPS J.Habitat exploration through morphological plasticity in two chalk grassland perennials[J].Oikos,1990,59:39-49.
[2] KROON H D,HUTCHINGS M J.Morphological plasticity in clonal plants:The foraging concept reconsidered[J].J Ecol,1995,83:143-152.
[3] GROENENDAEL J V,KROON H D.Clonal growth in plants:regulation and function[M].Hague:SPB Academic Publishing,1990.
[4] DONG M,KROON H D.Plasticity in morphology and biomass allocation inCynodondactylon, a grass species forming stolons and rhizomes[J].Oikos,1994,70:99-106.
[5] STUEFER J E.Potential and limitations of current concepts regarding the response of clonal plants to environmental heterogeneity[J].Vegetatio,1996,127:55-70.
[6] WIIJESINGHE D K,HUCHINGS M J.The effects of spatial scale of environmental heterogeneity on growth of a clonal plant:an experimental study withGlechomahederacea[J].J Ecol,1997,85:17-28.
[7] ALPERT P.Clonal integration in Fragaria chiloensis differs between populations:Ramets from grassland are selfish[J].Oecologia,1999,120:69-76.
[8] DONG M,ALATEN B.Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grassPsammochloavillosain an Inner Mongolian dune China[J].Plant ecology,1999,141(1/2):53-58.
[9] PENNINGS S C,CALLAWAY R M.The advantages of clonal integration under different ecological conditons:A community-wide test[J].Ecology,2000,81(3):709-716.
[10] HUTCHINGS M J,KROON H D.Foraging in plants:The role of morphological plasticity in resource acquisition[J].Advances in ecological research,1994,25:159-238.
[11] PITELKA L F,HANSEN S B,ASHMUN J W.Population biology ofClintoniaBorealis:I.Ramet and patch dynamics[J].Journal of ecology,1985,73:169-183.
[12] BIRCH C P D,HUTCHINGS M J.Explitation of patchily distributed soil resources by the clonal herbGlechomahederacea[J].J Ecol,1994,82(3):653-664.
[13] STUEFER J E,KROON H D,DURING H J.Exploitation of environmental heterogeneity by spatial division of labour in a clonal plant[J].Funct Ecol,1996,10(3):328-334.
[14] STUEFER J F,DURING H J,KROON H D.High benefits of clonal integration in two stoloniferous species,in response to heterogeneous light environments[J].J Ecol,1994,82(3):511-518.
[15] WIIJESINGHE D K,HANDEL S N.Advantages of clonal growth in heterogeneous habitats:An experiment withPotentillasimplex[J].J Ecol,1994,82(3):495-502.
[16] SAITOH T,SEIWA K,NISHIWAKI A.Importance of physiological integration of dwarf bamboo to persistence in forest understorey:A field experiment[J].Journal of ecology,2002,90:78-85.
[17] YU F H,DONG M,KRÜSI B.Clonal integration helpsPsammochloavillosasurvive sand burial in an inland dune[J].New phytologist,2004,162(3):697-704.
[18] ALPERT P.Nutrient sharing in natural clonal fragments of Fragaria chiloensis[J].Journal of ecology,1996,84(3):395-406.
[19] AMSBERRY L,BAKER M A,EWANCHUK P J,et al.Clonal integration and the expansion of Phragmites australis[J].Ecological applications,2000,10(4):1110-1118.
[20] YU F H,CHEN Y F,DONG M.Clonal integration enhances survival and performance ofPotentillaanserina, suffering from partial sand burial on Ordos plateau,China[J].Evolutionary ecology,2002,15(4/5/6):303-318.
[21] HARTNETT D C,BAZZAZ F A.The integration of neighbourhood effects by clonal genets in Solidago canadensis[J].Journal of ecology,1985,73(2):415-427.
[22] SALZMAN A G,PARKER M A.Neighbors ameliorate local salinity stress for a rhizomatous plant in a heterogeneous environment[J].Oecologia,1985,65:273-277.
[24] EVANS J P.The effect of resource integration on fitness related traits in a clonal dune perennial,Hydrocotylebonariensis[J].Oecologia,1991,86(2):268-275.
[25] WILLIAMS D G,BRISKE D D.Size and ecological significance of the physiological individual in the bunchgrassSchizachyriumscoparium[J].Oikos,1991,62(1):41-47.
[26] HESTER M W,MCKEE K L,BURDICK D M,et al.Clonal integration in Spartina patens across a nitrogen and salinity gradient [J].Canadian journal of botany,1994,72(6):767-770.
[27] BREWER J S,LEVINE J M,BERTNESS M D.Effects of biomass removal and elevation on species richness in a New England salt marsh[J].Oikos,1996,80(2):333-341.
[28] 刘凤红,叶学华,于飞海,等.毛乌素沙地游击型克隆半灌木羊柴对局部沙埋的反应[J].植物生态学报,2006,30(2):278-285.
[29] 董鸣.克隆植物的等级结构和等级选择[M]//张新时,高琼.信息生态学.北京:科学出版社,1997:136-141.
[30] 于飞海.克隆植物对异质性环境的生态适应对策[D].北京:中国科学院植物研究所,2002:97-98.
猜你喜欢
中国农村科技(2020年8期)2020-09-05
新世纪智能(英语备考)(2019年12期)2020-01-13
快乐语文(2019年14期)2019-08-07
爱你(2018年24期)2018-08-16
爱你·阳光少年(2018年8期)2018-05-14
新高考(英语进阶)(2017年3期)2017-05-04
环境昆虫学报(2016年5期)2016-11-18
食品工程(2016年1期)2016-11-14
中国照明(2016年4期)2016-05-17
读者(乡土人文版)(2015年10期)2015-12-13
