
The damage risk evaluation of Aphis gossypii on wheat by host shift and fitness comparison in wheat and cotton
2018-03-07 11:39FANYinjunLIFenAbdAllahMohammedYIXiaoqinZHANGMinNicolasDesneuxGAOXiwu
FAN Yin-jun, LI Fen, Abd Allah A.H.Mohammed,, YI Xiao-qin, ZHANG Min, Nicolas Desneux,GAO Xi-wu
1 Department of Entomology, College of Plant Protection, China Agricultural University, Beijing 100193, P.R.China
2 General Directorate of Plant Protection and Biological Control, Ministry of Agriculture/Animal Resources and Irrigation, Khartoum State 11111, Sudan
3 French National Institute for Agricultural Research (INRA), Université Côte d’Azur, CNRS, UMR 1355-7254, Institute Sophia Agrobiotech, Sophia Antipolis 06903, France
1.lntroduction
Host plant shifts are common in herbivorous insects.This allows insects to escape from natural enemies or competition and seek better nutrition (Blair et al.2010; Zvereva et al.2010).In addition, host plant shifts of herbivorous insects may cause sympatric speciation and can create new pests for agricultural crops (Bass et al.2013; Mattsson et al.2015).Life table analysis is able to provide some useful biological and demographic parameters for studying insect populations under specific conditions, and these parameters can be used as indicators for assessing the potential of the growth of a pest population under new conditions (Maia et al.2000; Diaz and Fereres 2005; Obopile and Ositile 2009).Therefore,in our study, the risk of species shift between cotton and wheat was assessed using this life table.These results will be used to monitor the occurrence of wheat aphids and to control aphid damage.
2.Materials and methods
2.1.Aphid collection
Aphids were collected from a wheat field (during wheat filling stage) in Wenshang County (Shangdong Province, China)in May of 2015 and were reared and maintained for four generations on wheat seedlings under controlled laboratory conditions (a climatic chamber with temperature of (22±1)°C,relative humidity (RH) of (70±10)%, and a photoperiod of 18 h L:6 h D).Apteral individuals were used as specimens for molecular identification.In addition, a colony of cotton aphid, which was maintained on cotton seedlings under the same laboratory conditions mentioned above for more than 17 years (since 1999) without exposure to insecticides, was used for host shift assessment.
2.2.Molecular identification
Individual was crushed in a 1.5-mL microcentrifuge tube and genomic DNA was extracted using DNAVzol(Vigorous Biotechnology Beijing Co., Ltd., China),according to the standard protocol recommended by the manufacturer.The primer sequences of LEP-F1(5´-ATTCAACAATCATAAAGATAT-3´) and LEP-R1(5´-TAAACTTCTGGATGTCCAAAAA-3´) were used to amplify mitochondrial gene COI (Lee et al.2015).The primer sequences of tRNALysAf2 (5´-GACTGAAAAGC AAAGTAATGATCTCT-3´) and CO3WWRD (5´-TCW CGAATWACATCWCGTCATCA-3´) were used to amplify ATP6 gene (Lee et al.2014).The length polymorphism of an intron in SCP was sequenced using the primers Aph13(5´-GATTGAATCAATGTGGGACTGCTTAC-3´) and Aph15(5´-ACTCAGCAACAACGCCAAGAAAAG-3´) (Carletto et al.2009a).All primers were synthesized by Huada Corporation,Beijing, China.
PCR reactions were performed in a 20-μL of reaction volume.Each reaction mixture contained 13 μL of water,0.5 μL of Taq polymerase, 2.5 μL of dNTPs (2.5 mmol L-1),2.0 μL of 10×buffer (+Mg2+)(TaKaRa Biotechnology Co., Ltd.,Dalian, China), 1 μL of 10 μmol L-1of each primer and 1 μL of DNA template (approximately 10 ng).PCR was performed using a GS1 Thermo-Cycler (Gene Technologies Ltd., UK)according to the following procedure: initial denaturation at 94°C for 5 min, followed by 30 cycles at 94°C for 30 s;an annealing temperature of 55°C for 30 s; an extension at 72°C for 40 s and a final extension at 72°C for 10 min.The PCR products were sequenced in both directions by using an ABI 3730xl Sequencer (Applied Biosystems,USA).Raw sequence data were examined and assembled using Sequencher 4.7 (Gene Codes Corporation, USA).Sequences were then aligned with Clustal X.
In order to more accurately identify whether the aphid is A.gossypii or not, the PCR primers (forward:5´-ATTTATAACCTATTATCAGCCATTT-3´ and reverse:5´-GAAATCTTAGTTTTATTCAAGTTAT-3´) were used to amplify COI of the aphid species, based on the complete mitochondrial gene of A.gossypii (KJ669654.1).The annealing temperature was 50°C, and other PCR conditions and methods were the same as above.
2.3.Phylogenetic analysis
In order to identify species, we retrieved 23 COI, 18 ATP6,and 5 SCP sequences (especially those of species closely related to A.gossypii and wheat aphids) from the GenBank database.These retrieved sequences and sequences of this species then were aligned with Clustal X (ver.2.0, 2007).All of the sequences to be analyzed were initially examined to merge forward and reverse strands using SeqMan Pro ver.7.1.0 (DNASTAR, Inc., Madison, WI).After the examination, the DNA sequences were aligned using MEGA Software.For the aligned data set, a neighbor-joining (NJ)tree was generated using MEGA based on the Kimura-2-Parameter (K2P) model.
2.4.Host shift
Laboratory experiments of the host shift between wheat and cotton were conducted to examine the survival and fitness of this aphid species and cotton aphids which have been maintained on cotton in the laboratory for more than 17 years(since 1999) with newly field-collected individuals added every two years without exposure to insecticides.Wheat cultivar Jimai 22 and cotton cultivar Xinluzao 7 were used in the study.In detail, four colonies of aphids were used in this experiment: I) aphids from a wheat field that were reared on wheat for four generations under laboratory conditions; II)aphids from the laboratory continued to rear on cotton under laboratory conditions; III) aphids from the first colony that were transferred to cotton and reared for four generations under laboratory conditions; and IV) aphids collected from the second colony that were transferred and reared on wheat for four generations under laboratory conditions.Thirty apterous adult aphids were taken from each colony and transferred individually to the hosts.Colonies I and III were transferred to the wheat seedlings while colonies II and IV were transferred to the cotton seedlings.After 24 h, 40 newborn nymphs from the mother aphids were transferred independently to plastic Petri dishes (3.5 cm in diameter) that contained a leaf of wheat or cotton on the internal surface.To prevent water loss and maintain ventilation, each Petri dish was punctured with two holes,and then they were covered with filter paper, wheat or cotton leaves, and the Petri dish lid successively.Forty newborn nymphs were individually inoculated in each Petri dish to start the life cycle of each colony.Petri dishes were placed in a climatic cupboard at (22±1)°C, (70±10)% RH,and for a 18 h L:6 h D photoperiod.The cotton or wheat leaves were replaced with fresh ones every two days, and the nymphal development was recorded every 24 h until the adult stage.After adults emerged, the survival and number of nymphs produced by females were recorded daily until the death of all adults.
比尔会在那里等他的,他们会顺着狄斯河向南划到大熊湖。接着,他们就会在湖里朝南方划,一直朝南,直到麦肯齐河。到了那里,他们还要朝着南方,继续朝南方走去,那么冬天就怎么也赶不上他们了。让湍流结冰吧,让天气变得更凛冽吧,他们会向南走到一个暖和的赫德森湾公司的站头,那儿不仅树木长得高大茂盛,吃的东西也多得不得了。
2.5.Data analysis
The variable developmental rate among individuals was taken into consideration, especially for the different host plants.These host shift data for aphids were analyzed according to the age-stage two-sex life table theory (Chi and Liu 1985; Chi 1988).The age-stage survival rate (sxj,the probability that a newborn aphid will survive to age x and stage j), the age-stage-specific fecundity (fxj), the agespecific survival rate (lx), and the age-specific fecundity(mx) were calculated based on recorded daily survival and reproduction rates.According to Chi and Liu (1985), the lxand mxfor the two-sex life table were calculated as follows:

Where, k is the number of stages.
The intrinsic rate of increase (r) was estimated using the iterative bisection method from the Euler-Lotka equation with age indexed from 0 (Goodman 1982):

The net reproductive rate (R) was calculated as:
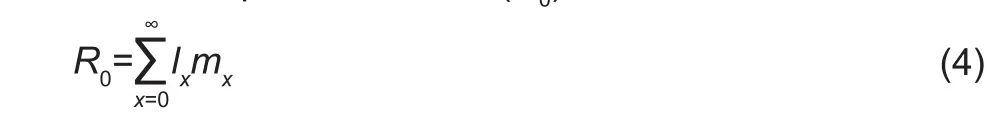
Mean generation time (T) is defined as the length of time that a population needs to increase to R0-fold of its original size, whereby the stable rate of increase (the intrinsic rate of intrease r and the finite rate of increase λ) is reached:

The finite rate of increase (λ) was calculated as:

Statistical analysis of raw data and the calculation of life table parameters were used for the computer program,TWOSEX-MSChart.The SE for the development time,fecundity, and life table parameters was estimated using the bootstrap technique (Huang and Chi 2012).
3.Results
3.1.Molecular identification
After alignment and excluding the primer sites, 676, 654 and 815 bp for COI, ATP6 and SCP were acquired.These sequences were blasted in NCBI; all the three sequences showed similarity to that of A.gossypii.Moreover, the detail alignment information between sequences of the aphid collected from wheat and sequences of A.gossypii were shown in Appendix A.The sequences of aphid collected from wheat and some other aphids were used for NJ analysis of distances (Fig.1).The NJ tree inferred from the COI,ATP6, and SCP genes showed that COI, ATP6, and SCP genes of the species collected from wheat and those of the A.gossypii are the same clade, and the bootstrap percentages are 80, 99, and 100, respectively.Nevertheless,they were not in the same clade with other species of wheat aphids S.avenae, R.padi (L.), and S.graminum (Rondani).
3.2.Fitness of A.gossypii collected from wheat and cotton
The results showed that the time of total pre-adult duration,adult longevity, and total longevity of the aphid population approximately were not affected when the aphids were transferred from wheat seedlings to cotton seedlings.However, the fecundity of the aphid population was significantly decreased when aphids were transferred from wheat seedlings to cotton (Table 1).Additionally, when the cotton aphids were transferred from cotton to wheat seedlings, the time of total pre-adult duration, adult, and longevity of the aphid population significantly prolonged.Meanwhile, after the cotton aphids were transferred from the cotton seedlings to the wheat, their fecundity significantly increased (Table 2).
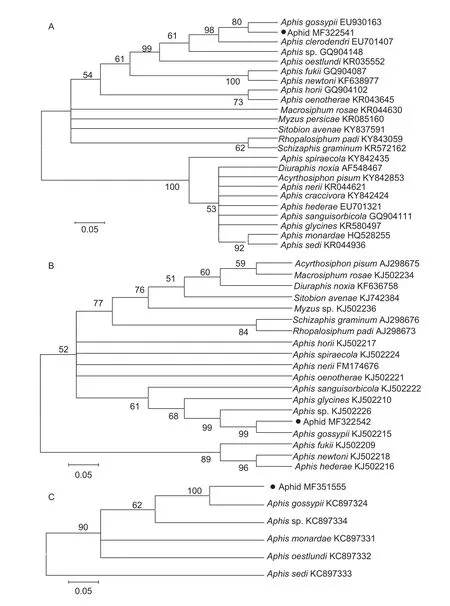
Fig.1 Neighbor-joining phylogenetic tree based on COI (A), ATP (B) and SCP (C) sequences using Kimuras two parameter model.refers to the aphid species collected from the wheat field in Wenshang County of Shangdong Province, China.
The intrinsic rate of increase, finite rate of increase, net reproductive rate, and mean generation time for cotton aphid fed on both wheat and cotton seedlings are shown in Tables 3 and 4.The intrinsic rate of increase, finite rate of increase, and net reproductive rate for cotton aphid population transferred from wheat seedling and fed on cotton seedlings were significantly lower than those initially reared on wheat seedlings.However, the effect of alternate hosts (from wheat to cotton) on the mean generation time for aphids showed no significant difference (Table 3).Accordingly, when aphid populations reared on cotton then transferred to wheat seedlings, their finite rate of increase was greatly lower than that of A.gossypii on cotton.The net reproductive rate and mean generation time of the aphids increased when the aphids transferred from cotton seedlings to wheat, but there was no significant effect on the intrinsic rate of the aphids.
The results of age-stage survival rate for A.gossypii showed that the developmental rate among individuals varied (Figs.2 and 3).The population of A.gossypii either reared on wheat or on cotton was capable to survive and reach the stage of adult.The adult stage of A.gossypii on wheat began at 5 days and ended at 34 days with a range of 30 days.The adult stage of A.gossypii on cotton that was transferred from wheat began at 6 days and ended at 34 days.The adult stage of A.gossypii on cotton began at 6 days and ended at 29 days, which is significantly shorter than that of A.gossypii on wheat that was transferred from cotton (beginning at 5 days and ending at 40 days).
The results for age-specific survival and fecundity of the aphids are shown in Figs.4 and 5, respectively.There was no significant difference of the age-specific fecundity among the four A.gossypii populations.However, the highest age-specific fecundity of the aphids decreased when they were transferred from wheat to cotton, whereas the highest age-specific fecundity of the cotton aphid was practically not affected when the host was shifted from cotton to wheat.
4.Discussion
A.gossypii is an extremely polyphagous and diverse species that colonizes more than 300 host plants (Wu et al.2013).This species can damage many crops, including melon,marrow, zucchini, potato, eggplant, cotton, ornamental hibiscus, and citrus fruit trees (Carletto et al.2009b).Moreover, A.gossypii is a phenotypically flexible species in response to a variety of environmental factors according to host plant diversity, localities, and abiotic conditions(Agrawal 2000; Margaritopoulos et al.2009).Additionally,it has formed various host plant specializations, including cotton-melon aphids, cotton-specialized aphids and cucurbits-specialized aphids (Liu et al.2008; Komazaki et al.2010) and the sequences of COI gene in cotton- and cucurbits-specialized biotypes of A.gossypii exhibit geneticdivergence (Zhang and Liu 2012).In our study, the strain collected from wheat can’t reproduce on cucumber but can reproduce on cotton by host shift experiments.In addition,no host plant specialization was found between wheat and cotton, and the effect of shifting hosts (from wheat to cotton or from cotton to wheat) on the viability of A.gossypii for a four-generation interval was not significant.Therefore, our results suggest that the A.gossypii colony from wheat has not yet developed wheat specialization, but the species was cotton-specialized biotype.
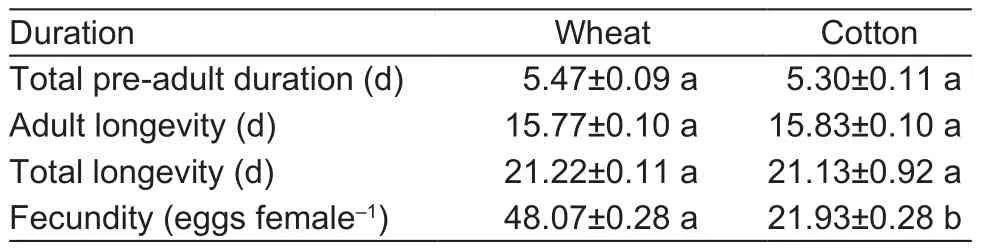
Table 1 Changes in the total pre-adult duration, adult longevity,total longevity, and fecundity of cotton aphids when they were transferred to cotton from wheat
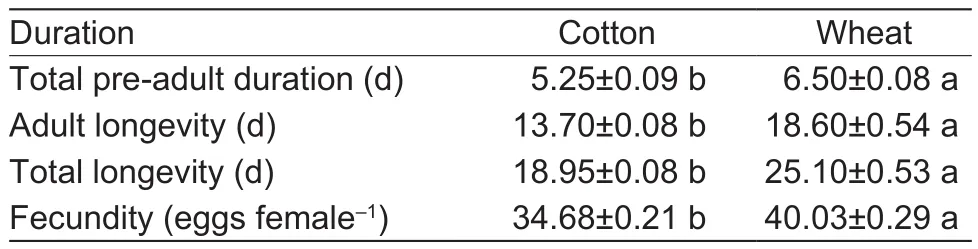
Table 2 Changes in the total pre-adult duration, adult longevity,total longevity, and fecundity of cotton aphids when they were transferred to wheat from cotton
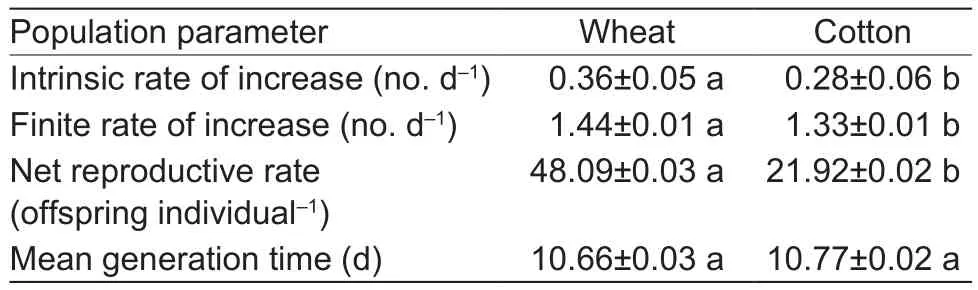
Table 3 Population parameters of Aphis gossypii that originated on wheat and then fed on wheat and cotton
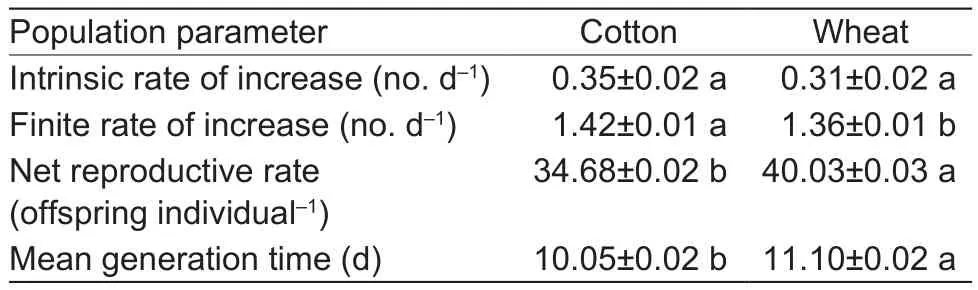
Table 4 Population parameters of Aphis gossypii that originated on cotton and then fed on cotton and wheat
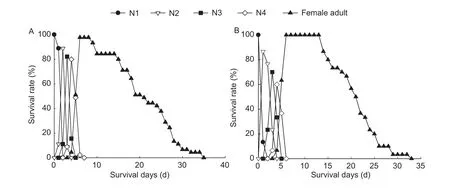
Fig.2 Age-stage survival rate (sxj) of Aphis gossypii that collected from wheat field of Wenshang County (Shangdong Province,China) and shifted to wheat (A) and cotton (B), respectively.N1, N2, N3 and N4 represent 1st, 2nd, 3rd and 4th nymphal instars of A.gossypii, respectively.
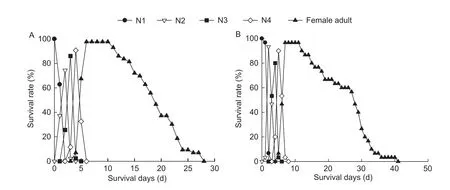
Fig.3 Age-stage survival rate (sxj) of Aphis gossypii that collected from cotton maintained in laboratory without insecticide exposure and shifted to cotton (A) and wheat (B), respectively.N1, N2, N3 and N4 are 1st, 2nd, 3rd and 4th nymphal instars of A.gossypii,respectively.
Plant chemistry, natural enemies and competition are considered as important factors related with host shifts in herbivorous insects (Murphy 2004; Zvereva et al.2010;Wu et al.2013).In our study, host shifts of A.gossypii between wheat and cotton could not be explained by the plant chemical similarity hypothesis because wheat and cotton belong to different plant families.Cotton aphid and common wheat aphids have similar (or same) enemies(Parajulee et al.1997; Men et al.2004).Thus, we presumed that A.gossypii feeding on wheat might be due to a special biochemistry-based mechanism which enables it to overcome the defense system of wheat, similar to Bass et al.(2013) who reported that Myzus persicae nicotianae that can feed on tobacco and its host shift resulted from the over expression of a cytochrome P450 enzyme (CYP6CY3)that was involved in nicotine-detoxifying.

Fig.4 Age-specific survival rate (lx), age-stage-specific fecundity (f(x,5)), age-specific fecundity (mx) and age-specific fecundity maternity (lxmx) of Aphis gossypii that collected from wheat field of Wenshang County (Shangdong Province, China) and shifted to wheat (A) and cotton (B).
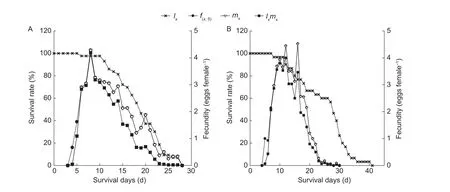
Fig.5 Age-specific survival rate (lx), age-stage-specific fecundity (f(x,5)), age-specific fecundity (mx) and age-specific fecundity maternity (lxmx) of Aphis gossypii collected from cotton that maintained in laboratory without insecticide exposure and shifted to cotton (A) and wheat (B).
The parameters of life table can be used to show the potential of pest population growth under new conditions(Diaz and Fereres 2005; Obopile and Ositile 2009).Considering the fitness of A.gossypii on wheat and cotton from our study, the fecundity and net reproductive rate of A.gossypii initially fed on wheat were significantly higher than that fed on cotton.These results suggest that when the population of cotton aphid fed on wheat may increase the reproduction rate compared with the population of this species fed on cotton.The different effects of wheat vs.cotton on the life table parameters of A.gossypii can result from plants with different antibiosis mechanisms which may refer to affect insect survival, size or weight, longevity, and reproduction (Sarfraz et al.2006; Rossa and Mclópez 2013).However, as we know, A.gossypii is very common in cotton fields, but the damage of A.gossypii to wheat in China has not previously been reported.This is possibly due to the fact that wheat is planted in fall/winter and harvested in summer in China (Ma et al.2006).Moreover, the reproduction mode of A.gossypii is sexual in fall/winter when their major hosts are Hibiscus syriacus (hibiscus), Punica granatum(pomegranate), and Zanthoxylum simulans (Chinese prickly ash) (Liu et al.2014; Wang et al.2016).Thus, we speculate that wheat is just a short-term host of asexual females early in summer when wheat is in a mature period and lacks adequate nutrition for aphids.
In many areas of northern China, cotton-wheat intercropping is an important cultural model beside monoculture cotton fields (Wang et al.2016).Winter wheat is sown from early- to mid-October with a designated row width left open for cotton, and is typically harvested from late May to mid-June of the following year.However,cotton is planted in mid-April, with the entire seedling stage overlapping the maturation phase of wheat (Ma et al.2006).Many natural enemies can hibernate in a wheat field.Several researchers have reported that populations of natural enemies increased significantly, whereas cotton aphid populations reduced in wheat-cotton intercropping,compared to monoculture cotton (Parajuleeet al.1997; Men et al.2004).Nonetheless, based on the investigation of our laboratory experiment, A.gossypii can feed on wheat, as well as it has a better reproduction rate than that of A.gossypii fed on cotton.In addition, the aphids were collected from wheat field (about 1 ha) located in smooth plain, and we did not observe a common host plant of A.gossypii nearby.Therefore, we doubt that there might be a risk that A.gossypii damage both wheat and cotton according to the cultural model of cotton-wheat intercropping.
5.Conclusion
The molecular identification of the aphid species collected from the wheat field of northern China was A.gossypii.Moreover, the results of host shift demonstrated that cotton aphid has potential to survive on wheat and has higher fecundity than that reared on cotton (i.e., the primary host plant).Therefore, this study raises a warning that the cotton aphid has potential to establish well on wheat and it may cause significant effects under specific circumstances.Therefore, future studies are required to evaluate the effects of A.gossypii on wheat production.
Acknowledgements
This research was supported by the Highland Barley Research System of China.
Appendixassociated with this paper can be available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
Agrawal A A.2000.Host-range evolution: Adaptation and trade-offs in fitness of mites on alternative hosts.Ecology,81, 500-508.
Bass C, Zimmer C T, Riveronc J M, Wilding C S, Wondji C S,Kaussmann M, Fielda L M, Williamsona M S, Nauen R.2013.Gene amplification and microsatellite polymorphism underlie a recent insect host shift.Proceedings of the National Academy of Sciences of the United States of America, 110, 19460-19465.
Blair C P, Schlanger R V, Diamond S E, Abrahamson W G.2010.Nutrition as a facilitator of host race formation: The shift of a stem-boring beetle to a gall host.Ecological Entomology, 35, 396-406.
Carletto J, Blin A, Vanlerberghe-Masutti F.2009a.DNA-based discrimination between the sibling species Aphis gossypii Glover and Aphis frangulae Kaltenbach.Systematic Entomology, 34, 307-314.
Carletto J, Lombaert E, Chavigny P, Brévaul T T, Lapchin L,Vanlerberghe-Masutti F.2009b.Ecological specialization of the aphid Aphis gossypii Glover on cultivated host plants.Molecular Ecology, 18, 2198-2212.
Chi H.1988.Life-table analysis incorporating both sexes and variable development rates among individuals.Environmental Entomology, 17, 26-34.
Chi H, Liu H.1985.Two new methods for the study of insect population ecology.Bulletin of the Institute of Zoology Academia Sinica, 24, 225-240.
Diaz B M, Fereres A.2005.Life table and population parameters of Nasonovia ribisnigri (Homoptera: Aphididae) at different constant temperatures.Environmental Entomology, 34,527-534.
Fagundes A C, Arnt T.1978.The occurrence of the aphid Aphis gossypii on wheat in Rio Grande do Sul.Agronomia Sulriograndense, 14, 143-144.
Goodman D.1982.Optimal life histories, optimal notation, and the value of reproductive value.The American Naturalist,119, 803-823.
Huang Y B, Chi H.2012.Assessing the application of the jackknife and bootstrap techniques to the estimation of the variability of the net reproductive rate and gross reproductive rate: A case study in Bactrocera cucurbitae (Diptera:Tephritidae).Jounal of Agriculture and Forest, 61, 37-45.
Komazaki S, Shigehara S, Toda S.2010.Diversity of Japanese Aphis gossypii and comparison with other Aphis species based on the mitochondrial cytochrome oxidase I sequence.Annals of the Entomological Society of America, 103,916-924.
Lee W, Lee Y, Kim H, Kimoto S, Lee S.2014.Developing a new molecular marker for aphid species identification:Evaluation of eleven candidate genes with species-level sampling.Journal of Asia-Pacific Entomology, 17, 617-627.
Lee Y, Lee W, Lee S, Kim H.2015.A cryptic species of Aphis gossypii (Hemiptera: Aphididae) complex revealed by genetic divergence and different host plant association.Bulletin of Entomological Research, 105, 40-51.
Liu L J, Zheng H Y, Jiang F, Guo W, Zhou S T.2014.Comparative transcriptional analysis of asexual and sexual morphs reveals possible mechanisms in reproductive polyphenism of the cotton aphid.PLoS ONE, 9, e99506.
Liu X D, Zhai B P, Zhang X.2008.Specialized host-plant performance of the cotton aphid is altered by experience.Ecological Research, 23, 919-925.
Lu Y, Gao X W.2009.Multiple mechanisms responsible for differential susceptibilities of Sitobion avenae (Fabricius)and Rhopalosiphum padi (Linnaeus) to pirimicarb.Bulletin of Entomological Research, 99, 611-617.
Ma X M, Liu X X, Zhang Q W, Zhao J Z, Cai Q N, Ma Y A,Chen D M.2006.Assessment of cotton aphids, Aphis gossypii, and their natural enemies on aphid-resistant and aphid-susceptible wheat varieties in a wheat-cotton relay intercropping system.Entomologia Experimentalis et Applicata, 121, 235-241.
Maia A H N, Luiz A J B, Campanhola C.2000.Statistical inference on associated fertility life table parameters using jackknife technique: Computational aspects.Journal of Economic Entomology, 93, 511-518.
Margaritopoulos J T, Kasprowicz L, Malloch G L, Fenton B.2009.Tracking the global dispersal of a cosmopolitan insect pest, the peach potato aphid.BMC Ecology, 9, 13.
Mattsson M, Hood G R, Feder J L, Ruedas L A.2015.Rapid and repeatable shifts in life-history timing of Rhagoletis pomonella (Diptera: Tephritidae) following colonization of novel host plants in the Pacific Northwestern United States.Ecology and Evolution, 5, 823-837.
Men X, Ge F, Yardim E, Parajulee M.2004.Evaluation of winter wheat as a potential relay crop for enhancing biological control of cotton aphids in seedling cotton.Biocontrol, 49,701-714.
Migui S M, Lamb R J.2003.Patterns of resistance to three cereal aphids among wheats in the genus Triticum (Poaceae).Bulletin of Entomological Research, 93, 323-333.
Murphy S M.2004.Enemy-free space maintains swallowtail butterfly host shift.Proceedings of the National Academy of Sciences of the United States of America, 101, 18048-18052.
Obopile M, Ositile M.2009.Life table and population parameters of cowpea aphid, Aphis craccivora Koch (Homoptera:Aphididae) on five cowpea Vigna unguiculata (L.Walp.)varieties.Journal of Pest Science, 83, 9-14.
Parajulee M N, Montandon R, Slosser J E.1997.Relay intercropping to enhance abundance of insect predators of cotton aphid (Aphis gossypii Glover) in Texas cotton.International Journal of Pest Management, 43, 227-232.
Rossa F, Mclópez A.2013.Effects of pepper (Capsicum annuum) cultivars on the biology and life table parameters of Myzuspersicae (Sulz.) (Hemiptera: Aphididae).Neotropical Entomology, 42, 634-641.
Sarfraz M, Dosdall L M, Keddie B A.2006.Diamondback mothhost plant interactions: Implications for pest management.Crop Protection, 25, 625-639.
Wang L, Zhang S, Luo J Y, Wang C Y, Lv L M, Zhu X Z.2016.Identification of Aphis gossypii Glover (Hemiptera:Aphididae) biotypes from different host plants in North China.PLoS ONE, 11, e0146345.
Wu W, Liang X L, Zhao H Y, Xu T T, Liu X D.2013.Special plant species determines diet breadth of phytophagous insects: A study on host plant expansion of the host-specialized Aphis gossypii Glover.PLoS ONE, 8, e60832.
Xin J J, Shang Q L, Desneux N, Gao X W.2014.Genetic diversity of Sitobion avenae (Homoptera: Aphididae)populations from different geographic regions in China.PLoS ONE, 9, e109349.
Zhang F, Liu X D.2012.Variation of host-specialized and migratory biotypes of Aphis gossypii Glover based on mtDNA COI gene sequences.Jounal of Nanjing Agriculture University, 35, 65-70.(in Chinese)
Zhang Y J, Jiang Y Y, Feng X D, Xia B, Zeng J, Liu Y.2009.Occurring trends of major crop pests in national significances in 2009.China Plant Protection, 29, 33-36.(in Chinese)
Zvereva E L, Kruglova O Y, Kozlov M V.2010.Drivers of host plant shifts in the leaf beetle Chrysome lalapponica:natural enemies or competition? Ecological Entomology,35, 611-622.
猜你喜欢
作文周刊·小学一年级版(2022年24期)2022-06-18
戏曲研究(2020年1期)2020-09-21
飞言情B(2020年5期)2020-07-04
飞言情B(2020年3期)2020-05-21
飞言情B(2020年2期)2020-05-09
飞言情B(2019年9期)2019-11-18
科学导报(2019年39期)2019-09-03
小天使·一年级语数英综合(2019年2期)2019-01-10
小学生导刊(低年级)(2017年1期)2017-06-12
知识窗(2016年8期)2016-05-14
 Journal of Integrative Agriculture2018年3期
Journal of Integrative Agriculture2018年3期
- Journal of Integrative Agriculture的其它文章
- Characteristic analysis of tetra-resistant genetically modified rice
- A wheat gene TaSAP17-D encoding an AN1/AN1 zinc finger protein improves salt stress tolerance in transgenic Arabidopsis
- Characterization of GhSERK2 and its expression associated with somatic embryogenesis and hormones level in Upland cotton
- GmNAC15 overexpression in hairy roots enhances salt tolerance in soybean
- Molecular cloning and functional characterization of a soybean GmGMP1 gene reveals its involvement in ascorbic acid biosynthesis and multiple abiotic stress tolerance in transgenic plants
- Responses of the antioxidant system to fluroxypyr in foxtail millet(Setaria italica L.) at the seedling stage