
大竹蛏β-整合素基因SgβInt的特性分析和重组表达*
2018-04-03 08:31刘佳乐韦秀梅杨顶珑刘相全
渔业科学进展 2018年1期
刘佳乐 韦秀梅 杨顶珑 童 潼 刘相全
大竹蛏-整合素基因的特性分析和重组表达*
刘佳乐1,2韦秀梅3杨顶珑3童 潼1刘相全3①
(1. 广西壮族自治区海洋研究所 广西海洋生物技术重点实验室 北海 536000;2. 上海海洋大学水产与生命学院 上海 201306;3. 山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室 烟台 264006)
整合素作为一种细胞粘附因子,在细胞粘附、迁移、增殖、凋亡和吞噬等过程中发挥重要作用。本研究利用分子克隆技术获得了大竹蛏()-整合素基因()的cDNA全长,分析了其编码氨基酸序列的进化特点,并以SWISS-MODEL预测了氨基酸序列的三级结构。基因序列全长为1168 bp,5′和3′非编码区(UTR)分别为61 bp和18 bp,开放阅读框(ORF)为1089 bp,编码362个氨基酸,理论等电点约为4.98,分子量为30.0 kDa。通过荧光定量PCR法检测了在健康大竹蛏各组织中以及大竹蛏受到脂多糖、肽聚糖和葡聚糖等微生物多糖刺激后的表达。结果显示,在血细胞、鳃、肝胰腺、性腺、肌肉和外套膜等组织中均可表达,在鳃中的表达量最高;脂多糖、肽聚糖和葡聚糖都可以诱导表达量上调,的表达量分别在脂多糖和葡聚糖刺激后的3 h和48 h达到最高;肽聚糖刺激后表达量上调幅度最大,在6 h时达到最高,证实参与大竹蛏抵御外源微生物的免疫应答。利用基因重组技术构建了的重组表达载体,为进一步分析软体动物整合素的功能、揭示大竹蛏先天性免疫机制奠定了基础。
大竹蛏;整合素;免疫应答;荧光定量PCR
细胞粘附因子(Cell adhesion molecules)介导细胞与细胞之间以及细胞与细胞外基质之间的相互作用,参与信号传导、细胞间识别、细胞增殖和分化、炎症和伤口愈合等过程(Zhuang, 2008)。根据结构和功能的不同,细胞粘附因子分为整合素(Integrin)、钙粘素(Cadherin)、选择素(Selectin)、免疫球蛋白超家族(Ig-superfamily)和透明质酸粘素(Hyaladherin)五类。其中,整合素是一种由一个α亚单位和一个β亚单位通过非共价键连接而成的异二聚体跨膜糖蛋白。目前,已经确认的有18种α亚单位和8种β亚单位,共组成20余种整合素(Jia, 2015)。大部分α亚基只结合一种β亚基,少数的如αV亚基可以与不同的β亚基结合。由于不同的α亚基和β亚基的氨基酸序列存在不同程度上的同源性,使得它们在结构上有共同的特点。整合素在体内表达广泛,大多数细胞表面都可表达一种以上的整合素,在多种生命活动中发挥关键作用。脊椎动物的Integrin作为一种重要的粘附受体,首先在淋巴细胞表面发现,目前,对于Integrin发挥的免疫功能的研究已经比较深入。Integrin细胞外的“头部”结构域可以结合配体,“颈部”是激活单位,而长度可变的细胞质内结构域形成细胞骨架起到链接信号分子的作用。此外,Integrin蛋白是双向信号转导分子(Hynes, 2002; Schwartz, 1995),在调控免疫反应过程中也发挥必不可少的作用(Rose, 2002; von Andrian, 2003)。近些年的研究表明,Integrin还参与肿瘤细胞粘附,比如,Integrin在口腔癌癌前病变时,起串联受体的作用(Shahidul, 2010);Integrin还被证明是人肠道弧菌病毒(Nelsen- Salz, 1999)、腺病毒(Wickham, 1993)、手足口病毒(Jackson, 2000)、乳头瘤病毒(Coulson, 1997)等多种病毒的受体。
在无脊椎动物中,果蝇() Integrin被研究得较为透彻,现已发现3个不同的α亚基(αPS1、αPS2和αPS3)和2个β亚基,3个位置特殊的Integrin异构体(PS1、PS2和PS3)与已被发现脊椎动物的β1-Integrin最为相似(Relja, 2011; Stark, 1997; Yee, 1993),有研究表明,果蝇-Integrin参与调控其血细胞吞噬细菌(Mamali, 2009)。在其他无脊椎动物中也相继发现了Integrin,例如,在冈比亚按蚊()中克隆得到的-Integrin基因,推测其参与血细胞介导的对大肠杆菌的包囊作用,相应的结构特征可能使其具有吞噬革兰氏阴性菌的能力(Moita, 2006);在海洋无脊椎动物中,凡纳滨对虾()-Integrin被认为是细胞膜上的对虾白斑综合征病毒(WSSV)受体(Li, 2007);中华绒螯蟹()-Integrin作为细胞受体参与抗菌免疫(Huang, 2015);仿刺参()-Integrin起到细胞凋亡抑制剂的作用(Wang, 2016)。与高等动物和模式动物相比,目前海洋无脊椎动物Integrin的研究尚处在起步阶段,有关贝类Integrin的报道十分有限。
本研究从前期已构建的大竹蛏() cDNA文库中克隆获得大竹蛏-Integrin ()基因的cDNA全长,分析了编码氨基酸的序列特征,通过荧光定量PCR法检测了基因的mRNA在健康大竹蛏不同组织中的表达谱,以及在大竹蛏受到脂多糖(LPS)、肽聚糖(PGN)和葡聚糖(Glucan) 3种不同病原相关分子(PAMPs)刺激后,血细胞中基因的响应规律,旨在了解基因在大竹蛏防御外源微生物的免疫应答中发挥的功能,为进一步阐释无脊椎动物Integrin的结构和功能、丰富无脊椎动物先天性免疫学研究提供支持。
1 材料与方法
1.1 实验动物
野生大竹蛏捕取自山东省烟台周边海区,体长为90 mm左右,共210只,实验前在20~22℃海水中暂养7 d。取其中10只大竹蛏用于解剖获得组织,剩余的200只随机均分为5组,用于PAMPs刺激实验,LPS、PGN和Glucan等3种PAMPs分别用PBS(0.14 mol/L NaCl, 3 mmol/L KCl, 8 mmol/L Na2HPO4, 1.5 mmol/L KH2PO4, pH=7.4)溶解,终浓度分别为0.5、0.8和1.0 mg/ml;以50 μl/只的剂量注射大竹蛏肌肉(Yang, 2011)。第1~3组分别注射LPS、PGN、Glucan,第4组注射PBS作为阴性对照,第5组未作任何处理作为空白对照。
1.2 RNA的提取和cDNA模板的合成
解剖大竹蛏,取肌肉、外套膜、血细胞、性腺、肝胰腺和鳃等,加入TRIzol (Invitrogen),存于–80℃。
PAMPs注射后3、6、12、24和48 h分别从实验组和对照组中随机取5只大竹蛏,注射器吸取预冷的抗凝剂(0.1 mol/L葡萄糖,13 mmol/L柠檬酸,15 mmol/L柠檬酸钠,0.45 mol/L NaCl,10 mmol/L EDTA,pH=7.0),抽取血淋巴,4℃ 800条件下离心5 min,收集血细胞,以1 ml TRIzol重悬,存于–80℃。
所有保存于TRIzol中的样品,按照TRIzol试剂盒说明书提取RNA,经DNase I (Promega)降解DNA,反应体系:RNA 8 μl, RQ1 RNase-Free DNase 5× Reaction Buffer 2.5 μl,RQ1 RNase-Free DNase 1 μl,RNase抑制剂0.6 μl,于37℃反应30 min;加入1 μl Stop solution,65℃ 10 min灭活DNase;加入2× Oligo dT 2.5 μ1,70℃反应5 min,冰浴2 min。反应体系:5×M-MLV反应缓冲液6.5 μl,RNase-Free dNTP 12.5 μl,RNase抑制剂1.25 μl,M-MLV反转录酶2.5 μl,RNase-Free 水1 μl,42℃反转录1 h,95℃ 5 min灭活反转录酶。模板存入–80℃,用于荧光定量PCR分析。
1.3 SgβInt基因cDNA全长的克隆
在本研究前期工作构建的大竹蛏cDNA文库中,用BLAST分析获得基因的EST序列,将对应质粒转入Trans5α感受态细胞(全式金)中,挑取单菌落,以pBluescript II SK*载体的通用引物M13F和M13R对其进行重新测序。
1.4 SgβInt基因的序列分析
在http://www.expasy.org/进行蛋白等电点和分子量预测;用BLAST软件进行序列同源性比较和相似性搜索;使用SingalP查找信号肽;使用MEGA 4.1软件,将基因和家蚕()、通讯鳌虾()、紫海胆()、双脐螺()、凡纳滨对虾、中华绒螯蟹、果蝇、冈比亚按蚊、仿刺参、鸭嘴海豆芽()、小菜蛾()、豌豆蚜()、长牡蛎()、瓜实蝇()、地中海实蝇()、意大利蜂()、华支睾吸虫()、粘虫()和大乳头水螅()等20个物种的-Integrin基因编码的氨基酸序列以邻接法(Neighbor- joining)构建系统进化树;使用SWISS-MODEL预测基因编码氨基酸序列的三级结构,利用pyMOL软件展示。
1.5 荧光定量PCR检测
将cDNA模板稀释100倍,使用Mastercycler ep realplex(Eppendorf)荧光定量PCR仪进行PCR扩增。反应体系20 μl:2×SYBR Green Master Mix (Applied Biosystems)10 μl、稀释100倍后的cDNA模板2 μl、正反引物各1 μl和6 μl水。的正向引物为5¢-AAGCGTGTCCGAGCAAATGTC-3¢,反向引物为5¢-TGTGGGGAGGGGTAGGATAAT-3¢;内参基因的正反向引物分别为5¢-TGTACGCCAACACTGTCCTGTC-3¢和5¢-CATCGTATTCCTGCTTGCTGATC-3¢。反应程序:50℃ 2 min,95℃ 10 min;94℃ 30 s,60℃ 60 s,40个循环。采用2-DDCt法分析实验数据,以检验分析显著性,<0.05为差异显著,<0.01为差异极显著。
1.6 SgβInt基因重组载体的构建
PCR反应扩增成熟肽DNA序列,反应体系:菌液模板1 μl,10×Buffer 2.5 μl,2.5 mmol/L dNTP 2 μl,正反引物各1 μl,酶1 μl,DEPC水补足至25 μl。重组正反引物分别为5¢-AACATGAGGTCCAT- GATAAACGG-3¢和5¢-TCATGTTGTAGTGTATGTC- TCACTGGTC-3¢。反应条件:94℃ 5 min;94℃ 30 s,62℃ 30 s,72℃ 80 s,35个循环;72℃ 10 min;4℃保存。PCR产物0.5~4.0 μl;PEASY E1 Expression Vector(全式金) 1 μl,混合后室温反应5 min,将连接产物加入Trans-T1(全式金)感受态细胞中,42℃水浴热激60 s;加入LB液体培养基,涂布平板,培养过夜。挑选单菌落由北京诺赛基因组研究有限公司测序。
1.7 SgβInt基因重组蛋白的诱导表达
将测序结果正确的菌株使用EZNA mini plasmid (Omega)试剂盒按照步骤提取质粒转入Transetta(全式金)感受态细胞中,涂布平板,37℃培养过夜。挑取单菌落,加入含氨苄青霉素(AMP)的LB培养基中,200 r/min、37℃震荡培养至OD600 nm约为0.6;加入终浓度为1 mmol/L的IPTG诱导表达目的蛋白,37℃ 200 r/min诱导培养4 h;4℃、5000 r/min离心10 min,收集菌体沉淀。以12%的SDS-PAGE电泳分析诱导表达结果。
2 结果
2.1 SgβInt基因cDNA的克隆及序列特点
测序结果拼接获得的基因cDNA全长,提交至GenBank获得登录号KX925404,其全长为1168 bp,5′和3′分别含61 bp和18 bp的非编码区(UTR),开放阅读框(ORF)为1089 bp,共编码362个氨基酸,其中,在42~74位氨基酸和103~135位氨基酸处分别为EGF like和EGF结构域。同源性分析显示,与家山羊()和小鼠()的相似性最高,分别为37%和36%;利用SignalP软件预测发现,基因编码区前22位氨基酸序列为信号肽序列(图1)。ExPASy预测重组后的基因编码蛋白的理论等电点约为4.98,分子量大小为30.0 kDa。
2.2 SgβInt基因系统进化分析
使用MEGA 4.1软件的邻接法构建系统进化树,所用Integrin的GenBank登录号如下:家蚕(NP_001161754.1)、通讯鳌虾(CAA67357.1)、紫海胆(XP_011679515.1)、双脐螺(AF060203.1)、凡纳滨对虾(ADK56123.1)、中华绒螯蟹(AKJ26284.1)、果蝇(AAC37169.1)、冈比亚按蚊(AAM11657)、仿刺参(AMR60824.1)、鸭嘴海豆芽(XM_013560388.1)、小菜蛾(GQ178290.1)、豌豆蚜(XM_003244399.3)、长牡蛎(EKC27174.1)、瓜实蝇(XP_011182368.1)、地中海实蝇(XP_004521716.1)、意大利蜂(XP_006564503.1)、华支睾吸虫(GAA31131.2)、粘虫(BAJ16205.1)、大乳头水螅(XP_002159375.2)。
用MEGA 4.1软件构建出编码的氨基酸与20个物种的-Integrin基因编码的氨基酸序列的系统进化树,结果显示,编码氨基酸与长牡蛎-Integrin编码氨基酸的亲缘关系最近,二者构成软体动物分支(图2)。

图1 SgβInt基因的核苷酸及其编码的氨基酸序列
黑体加粗表示起始密码子和终止密码子;下划线表示信号肽;阴影部分表示Integrin结构域。
The start and stop codons are marked in bold, the signal peptide sequence is underline and the Integrin is shaded.
2.3 SgβInt编码氨基酸的三级结构预测
使用SWISS-MODEL预测SgβInt中EGF-like、EGF和Integrin-β-tail三个结构域的三级结构,由图3可以看出,编码氨基酸的三维结构包括1个α-螺旋和9个β-折叠,α-螺旋靠近其中3个β-折叠,另外6个β-折叠每2个形成1组。
2.4 SgβInt基因的mRNA在健康大竹蛏各组织中的表达谱
运用荧光定量PCR方法检测了基因在健康大竹蛏不同组织中的表达规律,基因在健康大竹蛏的各个组织中均有表达,在鳃中表达量最高,是性腺的487.5倍;在肌肉、外套膜、血细胞和肝胰腺的表达量分别为性腺表达量的105.7、28.3、21.0、14.9倍;在性腺中表达量最低(图4)。
2.5 SgβInt基因的mRNA在大竹蛏受到PAMPs刺激后血细胞的表达规律
荧光定量PCR法检测LPS、PGN和Glucan刺激后大竹蛏血细胞中基因的表达规律,结果显示,基因的表达量在各种PAMPs刺激后均呈现上升趋势,并且都在刺激后3 h表现出极显著升高。其中,受到LPS刺激后,基因表达量在3 h达到最高(<0.01),为空白对照的4.8倍,随后逐渐降低,在12 h达到最低(<0.01),为空白对照的0.4倍;PGN刺激使基因的表达量上调幅度最大,继在刺激3 h时上升为空白组的10.8倍后(<0.01),在6 h达到最高,是空白对照的35.3倍(<0.01),随后显著降低;Glucan刺激也能诱导基因的表达量出现上调,在刺激3 h时为空白对照的3.2倍(<0.01),随后逐渐降低,但与LPS、PGN刺激后的表达模式不同的是,在24 h时再次出现显著升高(<0.05),并在48 h达到峰值,为空白对照的44.1倍(<0.01)(图5)。
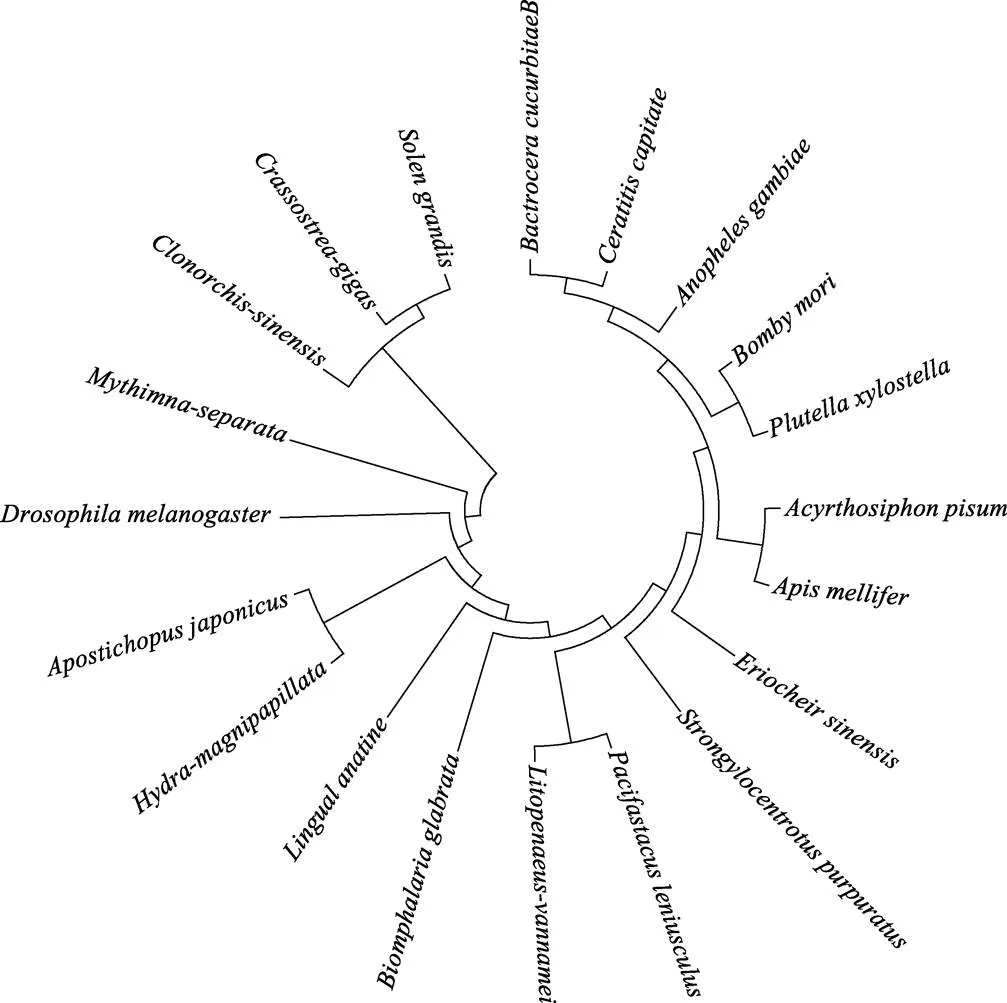
图2 SgβInt与其他物种β-Integrin的系统进化树

图3 SgβInt蛋白的三级结构
蓝色:β-折叠;红色:α-螺旋
Blue: β-sheets; Red: α-helices
2.6 SgβInt基因在大肠杆菌中的重组表达分析
SDS-PAGE分析对比诱导前后的SgβInt蛋白重组菌株,诱导菌株的泳道明显看到诱导蛋白的条带,重组蛋白的分子量大小约为30 kDa,与预测的结果相一致,证明基因编码蛋白被诱导表达(图6)。

图4 SgβInt mRNA在不同组织中的表达谱
3 讨论
整合素是细胞膜上粘附因子的一种,属于具有保守αβ异构体的细胞表面受体大家族(Hagen, 2001)。高等动物的整合素Integrin已被证明可作为人肠道弧菌病毒、腺病毒、手足口病毒、乳头瘤病毒等多种病毒的受体。近几年,陆续有研究发现,无脊椎动物整合素Integrin也参与多种免疫反应。而有关海洋无脊椎动物整合素Integrin的研究还相对较少。本研究以大竹蛏为研究对象,克隆得到了-Integrin基因的cDNA全长。通过Blast分析发现,该基因编码蛋白与家山羊和小鼠等脊椎动物的-Integrin基因编码蛋白相似,而分析基因编码蛋白的序列特点发现,在以邻接法构建的与其他物种-Integrin的系统进化树中,与长牡蛎-Integrin构成了软体动物分支;并且其三级结构包括1个α-螺旋和9个β-折叠,与孔雀鱼()和红鳍东方鲀()的-Integrin基因编码蛋白三维结构相似。综合以上分析结果,SgβInt为-Integrin家族成员。

图5 血细胞中SgβIntmRNA在PAMPs刺激后的表达水平
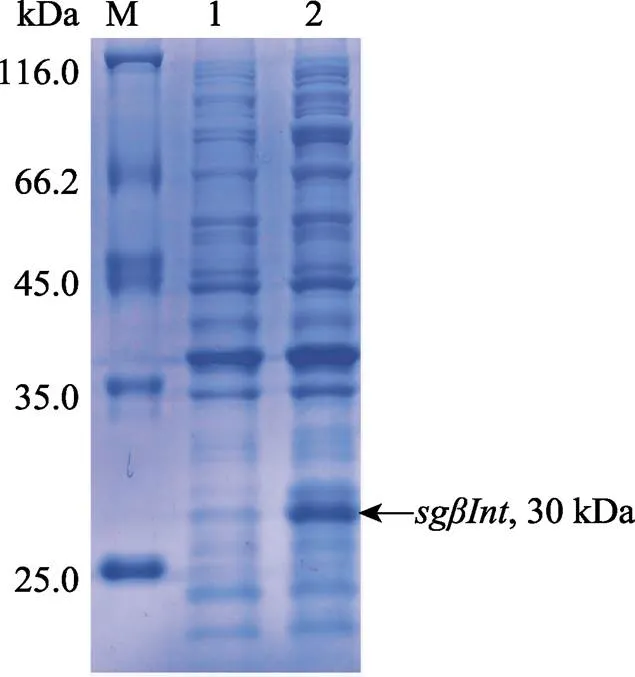
图6 重组SgβInt的SDS-PAGE分析
M: 蛋白质分子量标准;1: 未诱导的重组SgβInt;2: 诱导的重组SgβInt
M: Protein molecular standard; 1: Negative control for recombinant SgβInt (without induction); 2: IPTG induced recombinant SgβInt
仿刺参、长牡蛎、凡纳滨对虾等海洋无脊椎动物的Integrin基因具有组织分布广泛的特点,与以上报道类似,本研究发现,在所有检测组织中均有表达。组织表达谱广与-Integrin在机体的生命活动中参与多种过程密切相关。但是,这些物种的-integrin基因的组织表达谱存在差异,尤其是它们呈现高表达量的组织部位不同,如仿刺参-integrin在肌肉中的表达量最高(Wang, 2016),而长牡蛎和凡纳滨对虾-integrin则分别在性腺和血细胞中表达量最高(Lin, 2013; Jia, 2015)。在鳃的表达量最高,由于鳃是大竹蛏用于滤食和气体交换的器官,与水环境直接接触,考虑到水环境中存在多种外源微生物,基因在鳃中的高表达暗示其可能在大竹蛏抵御外源微生物的过程中发挥重要作用。
为分析是否参与大竹蛏抵御外源微生物的过程,本研究用LPS、PGN和Glucan等微生物多糖刺激大竹蛏,并检测血细胞中mRNA的表达规律。微生物多糖LPS、PGN和Glucan是微生物表面某些共有的高度保守的分子结构,宿主细胞不能自主产生这些多糖,为微生物所特有,是微生物生存或致病必需的。LPS、PGN和Glucan刺激都能诱导在大竹蛏血细胞中的表达显著上调,揭示参与大竹蛏抵御多种外源微生物的免疫过程。其中,PGN和Glucan刺激后,的表达量上调最为显著,说明可能更容易被革兰氏阳性菌和真菌诱导。中华绒螯蟹(Huang, 2015)和长牡蛎(Jia, 2015)的-Integrin在受到LPS刺激后6 h时达到最高表达量,而本研究中,一方面的表达量在3 h时达到最高,大竹蛏对于LPS的响应时间早于中华绒螯蟹和长牡蛎Integrin。另一方面,在大竹蛏受到PGN和Glucan刺激后达到最高表达量的时间不一致,其对于不同微生物多糖的响应时间不同,推测其清除作用机制存在差异。
重组的基因编码蛋白是研究基因功能的有利工具。利用制备的重组蛋白,已有研究发现,中华绒螯蟹的-Integrin可以包括凝集金黄色葡萄球菌、藤黄微球菌、大肠杆菌在内的多种细菌,并能够协助机体清除副溶血弧菌等致命菌(Huang, 2015);而长牡蛎和仿刺参的-Integrin都对LPS有较高的亲和力(Lin, 2013; Wang, 2016);构建SgβInt编码蛋白的重组载体,为获得SgβInt重组蛋白,制备抗Integrin重组蛋白抗体,进一步分析SgβInt编码蛋白的功能提供了必要的实验材料。SgβInt编码蛋白对于外源微生物的作用方式还不明确,有待于进一步研究。
本研究中基因的克隆、特性分析及其重组载体的构建,揭示了基因的序列特点及其对于PAMPs的响应规律,为了解大竹蛏对外源微生物的免疫防御机理和进一步分析其作用机制奠定了基础。
Coulson BS, Londrigan SL, Lee DJ. Rotavirus contains integrin ligand sequences and a disintegrin-like domain that are implicated in virus entry into cells. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(10): 5389–5394
Hagen HE, Kläger SL. Integrin-like RGD-dependent cell adhesion mechanism is involved in the rapid killing ofduring early infection ofs.l. Parasitology, 2001, 122(4): 433–438
Huang Y, Zhao LL, Feng JL,. A novel integrin function in innate immunity from Chinese mitten crab (). Developmental and Comparative Immunology, 2015, 52(2): 155–165
Hynes RO. Integrins: Bidirectional, allosteric signaling machines. Cell, 2002, 110(6): 673–687
Jackson T, Sheppard D, Denyer M,. The epithelial integrin alpha-vbeta 6 is a receptor for foot-and-mouth disease virus. Journal of Virology, 2000, 74(11): 4949–4956
Jia ZH, Zhang T, Jiang S,. An integrin from oystermediates the phagocytosistowardthrough LPS binding activity. Developmental and Comparative Immunology, 2015, 53(1): 253–264
Li DF, Zhang MC, Yang HJ,. β-integrin mediates WSSV infection. Virology, 2007, 368(1): 122–132
Lin YC, Chen JC, Chen YY,. Characterization of whiteshrimpintegrin β and its role in immunomodulation by dsRNA-mediated gene silencing. Developmental and Comparative Immunology, 2013, 40(2): 167–179
Mamali I, Lamprou I, Karagiannis F,. A β integrin subunit regulates bacterial phagocytosis in medfly haemocytes. Developmental and Comparative Immunology, 2009, 33(7): 858–866
Moita LF, Vriend G, Mahairaki V,. Integrins ofand a putative role of a new β integrin, BINT2, in phagocytosis of. Insect Biochemistry and Molecular Biology, 2006, 36(4): 282–290
Myat MM, Rashmi RN, Manna D,KASH- domain protein Klarsicht regulates microtubulestability and integrin receptor localization during collective cellmigration. Developmental Biology, 2015, 407(1): 103–114
Nelsen-Salz B, Eggers HJ, Zimmermann H. Integrin alpha (v) beta3 (vitronectin receptor) is a candidate receptor for the virulent echovirus 9 strain Barty. Journal of General Virology, 1999, 80(9): 2311–2313
Relja B, Meder F, Wang M,. Simvastatin modulates the adhesion and growth of hepatocellular carcinoma cells via decrease of integrin expression and ROCK. International Journal of Oncology, 2011, 38(3): 879
Rose DM, Han J, Ginsberg MH. Alpha4 integrins and the immune response. Immunological Reviews, 2002, 186(1): 118–124
Schwartz MA, Schaller MD, Ginsberg MH. Integrins: Emerging paradigms of signal transduction. Annual Review of Cell Developmental Biololgy, 1995, 11: 549–599
Shahidul A, Yamazaki M, Maruyama S,Differential expression of perlecan receptors, α-dystroglycan and integrin β1, before and after invasion of oral squamous cell carcinoma. Journal of Oral Pathology and Medicine, 2011, 40(7): 552–559
Stark KA, Yee GH, Roote CE,. A novel alpha integrin subunit associates with betaPS and functions in tissue morphogenesisand movement duringdevelopment. Development, 1997, 124(22): 4583–4594
von Andrian UH, Engelhardt B. Alpha4 integrins as therapeutic targets in autoimmune disease. New England Journal Medicine, 2003, 348(1): 68–72
Wang ZH, Shao Y, Li CH. A β-integrin from sea cucumberexhibits LPS binding activity and negatively regulates coelomocyte apoptosis. Fish and Shellfish Immunology, 2016, 52: 103–110
Wickham TJ, Mathias P, Cheresh DA,. Integrins-alpha- v-beta-3 and alpha-v-beta-5 promote adenovirus internalization but not virus attachment. Cell, 1993, 73(2): 309–319
Yang JL, Wang LL, Zhang Huan,. C-type lectin in(CfLec-1) mediating immune recognition and opsonization. PLoS One, 2011, 6(2): e17089
Yee GH, Hynes RO. A novel, tissue-specific integrin subunit, beta nu, expressed in the midgut of. Development, 1993, 118(3): 845–858
Zhuang SF, Kelo L, Nardi JB,. Multiple α subunits of integrin are involved in cell-mediated responses of the Manduca immune system. Developmental and Comparative Immunology, 2008, 32(4): 365–379
(编辑 冯小花)
Recombinant Expression and Characterization of the-Integrin Gene of
LIU Jiale1,2, WEI Xiumei3, YANG Dinglong3, TONG Tong1, LIU Xiangquan3①
(1. Guangxi Institute of Oceanology, Key Laboratory of Marine Biotechnology of Guangxi, Beihai 536000; 2. College of Fisheries and Science, Shanghai Ocean University, Shanghai 201306; 3. Key Laboratory of Marine Ecological restoration, Shandong Marine Resource and Environment Research Institute, Yantai 264006)
Integrins are heterodimeric cell surface receptors that consist of α and β subunits to regulate cell adhesion, migration, proliferation, apoptosis and phagocytosis. The present study identified the-integrin gene from() and analyzed the characterization of its encoded protein. The phylogenetic tree was constructed by the neighbor-joining method and the three-dimensional structure was predicted with SWISS-MODEL. The full-length cDNA ofwas 1168 bp, containing a 61 bp 5´UTR, 18 bp 3´UTR and an open reading frame (ORF) of 1089 bp that encodes a polypeptide of 362 amino acids with an estimated molecular mass of 30.0 kDa. The encoded protein ofshared significant similarities with that inand. The phylogenetic tree indicated thathad a close genetic relationship withto form a mollusk branch. The three-dimensional structure of SgβInt consisted of one α-helice and nine β-sheets as a member of integrin superfamily.expressed in all tested tissues, including mantle, gill, hemocyte, gonad, muscle and hepatopancreas, with the highest expression in gill which was 487.5 times higher than gonad.was induced by all the three pathogen associated molecular patterns (PAMPs) including LPS, PGN and glucan with the peak level at 3 and 48 h post LPS and glucan stimulation, respectively.expression reached the maximal level at 6 h post PGN stimulation with 53.5-fold induction. All the results revealed thatmight regulate the immune response ofto microorganism. This study shed new light on the research of-integrin in mollusk and immune defense mechanism of.
; Integrin; Immune response; Real-time PCR
LIU Xiangquan, E-mail: lxq6808@163.com
2016-12-26,
2017-01-14
S968.3
A
2095-9869(2018)01-0120-08
10.11758/yykxjz.20161226001
http://www.yykxjz.cn/
* 广西海洋生物技术重点实验室开放基金资助课题资助[This work was supported by Open Research Fund Program of Guangxi Key Laboratory of Marine Biotechnology (GLMBT-201504)]. 刘佳乐, E-mail: liujlwz@163.com
刘相全,副研究员,E-mail: lxq6808@163.com
刘佳乐, 韦秀梅, 杨顶珑, 童潼, 刘相全. 大竹蛏整合素基因的特性分析和重组表达. 渔业科学进展, 2018, 39(1): 120–127
Liu JL, Wei XM, Yang DL, Tong T, Liu XQ. Recombinant expression and characterization of the-integrin gene of. Progress in Fishery Sciences, 2018, 39(1): 120–127
猜你喜欢
医学综述(2022年10期)2022-11-28
传染病信息(2022年2期)2022-07-15
中国药学药品知识仓库(2022年9期)2022-05-23
锦州医科大学报(2022年2期)2022-05-07
现代临床医学(2021年2期)2021-03-29
口腔医学(2020年6期)2020-12-19
癌变·畸变·突变(2020年4期)2020-08-05
当代县域经济(2018年9期)2018-09-05
文苑·经典美文(2017年6期)2017-06-12
当代县域经济(2015年12期)2015-03-20
