
不同放牧强度对锡林郭勒典型草原生产力及碳储量的影响
2018-05-05 08:53杜宝红高翠萍哈达朝鲁
水土保持研究 2018年1期
杜宝红, 高翠萍, 哈达朝鲁
(1.锡林郭勒职业学院, 内蒙古 锡林郭勒盟 026000; 2 内蒙古农业大学, 呼和浩特 010011)
草原作为世界上分布最广的重要的陆地植被类型之一,在地球表面占据着一定的自然地带,分布面积广泛[1-2]。全球草原面积约5.0×109hm2,约占全球陆地总面积的40%,中国的草原面积为3.55亿hm2,约占国土面积的41.7%,是世界草原总面积的6%~8%,居世界第二,主要类型为温带草原、高寒草原和荒漠草原等[3-4]。内蒙古草原不仅是国家重要的畜牧业生产基地,同时也是重要的绿色生态屏障,在减少沙尘暴和恶劣天气的发生方面发挥着作用,也是研究生态系统对人类干扰和全球气候变化响应机制的典型区域之一和国际地圈—生物圈计划(IGBP)陆地样带—中国东北陆地生态系统样带(NECT)的重要组成部分[5-7],但过度放牧和不合理利用,草原发生了不同程度的退化,甚至发生了大面积的沙化、盐碱化,探讨最大限度地优化草原具有重要的意义。
在全球气候变化成为国际上备受瞩目的问题的同时,我国科学家也开始研究管理模式对土壤碳库和碳循环的影响[8-9]。草原生态系统是陆地碳循环及碳固持的一个重要组成部分,随着全球变暖等环境问题日益突出,生态系统的碳循环成为人们研究的热点[10-12]。我国草原生态系统是陆地生态系统的主体类型,贮存的碳总量约为266.3 Pg,占陆地生态系统的12.7%,其中90%贮存在土壤中。受气候变化及长期不合理的人为活动干扰,我国90%的可利用草原处于不同程度的退化状态,其中20%以上是由于过度放牧造成的,碳正常循环受到影响[10-12]。管理农事对碳循环及其生态过程的效应研究受到了土壤,环境,全球变化的影响,其中放牧导致的土地退化会使草原生态系统功能发生衰退,影响植被生产力及土壤有机质的分解和积累速率,进而影响土壤碳储量和生态系统的碳循环,而适度利用可减少草原碳排放,增加土壤碳贮存,故针对不同草原生态系统类型,科学安排利用方式和强度对草原可持续利用尤为重要,这已成为草原管理研究领域的重要命题之一[13-14]。因此,开展放牧草原植被—土壤碳储量的研究,不仅可以揭示放牧强度对草原植被—土壤碳储量变化的影响,而且可以阐明放牧草原碳储量各组分间的比例及其转化关系,进而从维护碳汇的角度制定可持续利用的放牧制度[15]。典型草原是内蒙古草原的重要组成部分,属于草原区向荒漠区过渡的旱生化草原生态系统,群落结构较单一化,种群密度和群落结构容易发生改变[5-6]。已有研究表明,在不同的放牧强度下或连续过度放牧下,典型草原草原植物种类组成改变,地表植被的生物量、生物多样性和丰富度下降土壤容重增加,孔隙度减小,土壤质量退化[13-14]。土地与植被的退化必然会引起草原有机碳含量的变化,那么,典型草原不同放牧强度下土壤、植物的有机碳含量是否存在显著差异以及如何分配,二者的相互关系如何?加强这些问题的研究有助于揭示放牧对典型草原碳汇的影响机制。有鉴于此,本研究以羊草典型草原为研究对象,研究不同放牧强度下植被—土壤系统有机碳储量,旨在对比不同放牧强度典型草原有机碳含量与储量的变化及其差异性,为科学制定草原保护、利用策略和提高草原生态生产功能的适应性供科学依据。
1 材料与方法
1.1 研究区概况
本试验依托内蒙古锡林郭勒草原生态系统国家野外科学观测研究站(内蒙古站,43°38′N,116°42′E,海拔1 224 m)进行。位于内蒙古锡林郭勒盟锡林河流域中段,即中国科学院草原生态系统定位站1979年围封针茅草原附近,该站位于内蒙古锡林河流域,气候属温带半干旱大陆性季风气候,冬季寒冷干燥,夏季受季风影响,较为温暖和湿润。1970—2010年平均气温为0.4℃,其中最冷月(1月)平均气温为-21.4℃,最热月(7月)平均气温为19.0℃。年降水量为337 mm,无霜期约为100 d,6-8月降水量占全年降水量的70%,降水量年际变异较大。地带性土壤类型为暗栗钙土,土层厚度可达1 m以上,地带性植被为大针茅草原。试验样地位于羊草(Leymuschinensis)样地北侧,2004年通过国际合作项目由中德双方共同建设,整个试验区占地面积约300 hm2,群落类型为羊草+大针茅草原。试验处理前,群落物种分布比较均匀,常见植物有45种,其中以广旱生根茎禾草羊草占优势,其次为大针茅、阿尔泰草、冰草、糙隐子草等多年生丛生禾草,这些禾草的地上生物量占群落总生物量的70%以上。常见种和稀有种均以杂类草为主,包括:星毛委陵菜、野韭等[3-5]。
1.2 试验设计
从2011年8月下旬开始,每年进行放牧处理,试验用绵羊统一选择健康的2岁母羊,放牧处理于每年的6月上旬开始,9月中旬结束。采用连续放牧方式,晚上羊群在样地不归牧,不进行补饲。在羊草草原选择4个代表类型,其中对照样地为1979年围栏封育样地(无放牧,NG),另外3个样地为放牧退化样地,选取以自由放牧方式利用的草原为研究对象,以牧户为起点向外辐射状草原形成由重到轻的一个放牧退化梯度,根据草原植物种类组成、地上生物量、群落高度、群落盖度等指标,划分为对照(无放牧,NG)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG),不同重复小区内的4个处理完全随机排列。设定不同的载畜率值分别为0(对照),0.91(轻度放牧),1.82(中度放牧)和2.71(重度放牧)羊单位/hm2/半年[3-5]。
利用随机样方法,2016年8月在NG,LG,MG和HG样地分别设置1 m×1 m样方10个,记录和测定植物种类组成、盖度和高度;筛选其中5个样方采用齐地面刈割牧草的方法测定地上生物量鲜重,带回实验室65℃烘干至恒重;地下生物量与地上生物量在同一样方内测定,采用剖面法,进行10 cm×10 cm×10 cm土块切割,每10 cm为一层进行取样,取到60 cm深度,将取得的样品带回实验室进行筛根、洗根、烘干、称重。土壤样品亦在上述样方处利用土钻取样,取样深度为0—10,10—20,20—30,30—40,40—50,50—60 cm,每样方内取3次后按层混合,带回实验室风干、过筛,测试相关指标,并在每个样方挖取土壤坡面,环刀法测定土壤容重。取回的根样用水洗法将其漂出,装入信封。将根样与草样一起烘干(65℃)称重,得到地上部分、地下部分生物量的值(g/m2)。同时记录群落地形、地貌特征和土壤质地,GPS定位样地经纬度和海拔。将根样与草样分别粉碎并过80目的筛,然后用元素分析仪测出含碳量(%)。
土壤—植物有机碳测定采用重铬酸钾-外加热法,土壤碳储量计算公式如下[11,16]:
式中:P为土壤碳储量(g/m2);di为有机碳含量(%);Hi为土层高度(cm);bi为土壤容重(g/cm3);n为土层深度(cm)。
多样性指数主要用来反映植物的丰富程度,是植物丰富度和均匀程度的综合反映,多样性指数越高,植被的物种多样性越丰富。均匀度是植物分布均匀程度的反映,在一个植被中,各个种的相对密度越均匀,即各物种的个体数很接近,均匀度值越高。丰富度是植被物种多样性丰富程度的反映,当个体数量一定时,物种数越多,计算公式如下[17]:
Patrick丰富度指数(S)
Shannon-Wiener多样性指数(H):
H=-∑PilnPi
Gleason优势度指数(D):
D=1-∑(Pi)2
Pielou均匀度指数(JP):
JP=-∑PilnPi/lnS
式中:S代表样方内物种数目;Pi为样方内种的相对重要值;相对重要值(Pi)=(相对覆盖度+相对高度+相对多度)/3。此外,为了比较不同类型植被多样性指数间的关系,方差分析和多重比较用来检验它们之间的差异显著性。
Excel 2007和SPSS 18.0进行数据统计和方差检验,以平均值±标准误差表示(Mean±SE),单因素方差分析(One-way ANOVA)。
2 结果与分析
2.1 放牧强度对物种特征的影响
放牧是天然草原群落受干扰的主要方式之一。在牧压梯度上,植物群落及其主要植物种群的数量特征发生明显的变化,主要表现为植物群落密度MG>LG>NG>HG,过牧区最低(表1)。优势植物羊草的种群密度在中牧区最大,轻牧区次之,重牧区最低,而其相对密度从轻牧到重牧的梯度上逐渐降低。这些变化是由于不同强度的放牧牲畜践踏,羊草种群大株丛破碎,由一个母株分离出若干个独立小株丛的结果。羊草种群密度表现为MG>LG>NG>HG。表明适度的放牧有利于羊草根茎的生长发育,但超过一定的阈值后,如在过牧区羊草种群也会迅速消退。大针茅的种群密度MG>LG>NG>HG,表现出一定的耐牧性。禾本科草的种群密度从轻牧区到重牧区逐渐降低。综上所述,天然草原在不同强度的放牧干扰下,植物生态适应对策是多样的,这种多样性是由植物自身生物生态学特性决定的。天然草原群落正是由于多样的植物生态适应对策,对放牧干扰产生明显的缓冲作用,表现为一定范围内的草原群落抗干扰能力和系统稳定性。
2.2 放牧强度对物种多样性的影响
多样性指数是植被和生态系统恢复过程最重要的特征之一,是群落生物组成结构的重要指标,不仅能够反映植被群落组织化水平,而且通过结构与功能的关系可间接反映群落功能的稳定性和发育阶段。由图1可知,随着放牧强度的增加,典型草原植被覆盖度、丰富度指数、均匀度指数、多样性指数、优势度指数均降低,其中LG,MG和HG均与NG达到显著差异水平(p<0.05),并且均表现出NG>LG>MG>HG变化趋势;轻度放牧丰富度指数、均匀度指数和多样性指数与中度放牧差异不显著(p>0.05)。HG具有显著较低的群落均匀度水平,MG样地的均匀度水平最高,显著高于3个不同管理方式不同利用程度的羊草群落(p<0.05)(图1)。这反映了较低水平的利用程度使群落内物种重要值的差异逐渐变大,优势种的优势程度被增强,群落的均匀程度降低,而HG降低了物种创造低竞争压力的生存机会,从而群落内各种的重要值和群落地位差异并未充分显露和稳定,使得群落的均匀度呈现较高水平。在轻牧区,群落有较低的植物均匀度和多样性,随着放牧强度增加,群落中的优势植物逐渐被一些耐牧、适牧植物所替代,使中牧区的群落具有较高的植物多样性和均匀度,如果继续增加放牧强度,群落的均匀度和多样性又降低,直至群落结构遭到致命的破坏。

表1 放牧强度对物种特征的影响

注:不同小写字母表示差异显著(p<0.05)。下同。
图1放牧强度对物种多样性的影响
2.3 放牧强度对地上生物量及碳储量的影响
由于植物光合产物的生产和累积能力,以及牲畜的采食方式和喜食程度的差异,不同放牧强度下植物群落地上生物量的差异较大,导致地上植物有机碳储量差异显著(图2)。由图可知,随着放牧强度增加,地上生物量和凋落物量呈递减趋势,NG地上植被和凋落物碳储量最高,其大小依次为NG>LG>MG>HG,放牧显著降低了群落的地上植物有机碳储量(p<0.05),除LG与MG之间差异不显著外,其余放牧梯度间差异显著(p<0.05)。以NG作对照,LG,MG,HG平均群落地上生物量分别下降15.63%,25.98%和46.12%。地表凋落物也表现出同样的规律,与群落地上生物量相比下降的更为明显。放牧干扰降低了植物群落的生产力,直接影响植物对土壤水分和营养元素的吸收,使有机物质生产和地表凋落物累积的减少,归还土壤的有机质降低,回归土壤养分和保持水分的能力下降,给土壤的理化性状造成影响,从而导致土壤贫瘠化和干旱化,进而导致群落结构的简单化和生态功能的下降。
2.4 放牧强度对根系碳储量的影响
草原生物量的80%以上集中在地下,地下生物量是土壤有机碳库的最主要输入源,在草原生态系统碳循环中起着关键作用。分析放牧强度对根系碳储量的影响,结果表明,典型草原植物地下部分(根系)有机碳储量明显高于地上部分有机碳储量(表2),随着土层深度的增加,根系有机碳储量呈现减少的趋势,均以0—10 cm到10—20 cm下降的量最多。从地下0—60 cm根系有机碳储量来看,大小序列仍为NG>LG>MG>HG,NG和LG差异不明显,但显著高于MG和HG(p<0.05)。0—10 cm根系有机碳储量占到0—60 cm根系总有机碳储量的50%以上,0—20 cm根系有机碳储量占到0—60 cm根系总有机碳储量的20%以上,因此根系总有机碳储量在不同放牧强度间的变化主要受0—20 cm根系有机碳储量的影响,信息量可达70%以上,0—20 cm根系有机碳储量仍以NG最高,HG最低。

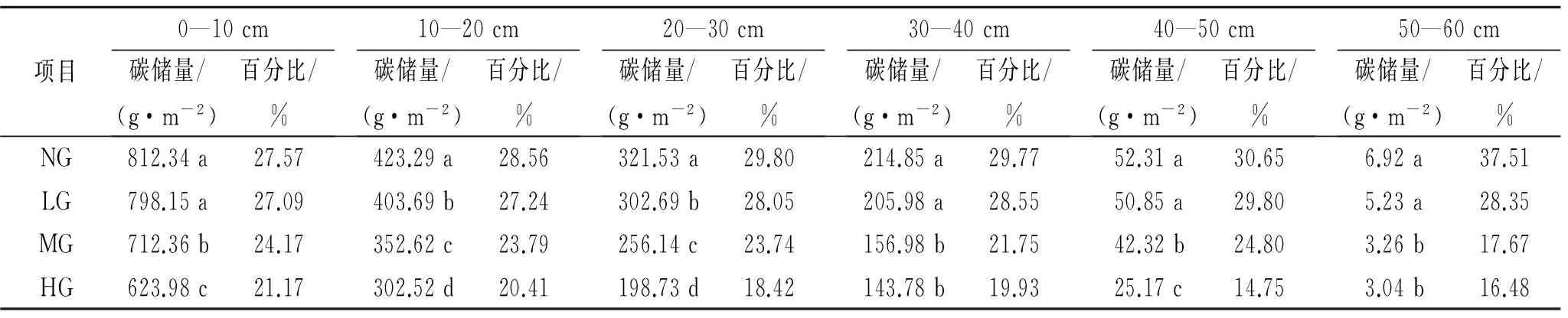
图2 放牧强度对地上生物量及碳储量的影响
2.5 放牧强度对土壤有机碳储量的影响
土壤是草原生态系统中碳的主要存储库,占系统的80%以上。放牧利用在明显影响植被生物量的同时,也会对土壤的碳储量产生影响,致使草原土壤碳储量产生差异。本研究比较了不同放牧强度下草原的土壤碳储量(表3),结果显示,土壤有机碳含量随放牧强度的增加呈先增加后减少趋势,并在MG和LG样地达到最大。不同放牧强度只对0—40 cm土层有机碳含量影响差异显著,在40 cm以下土层,其影响不显著(p>0.05)。土壤有机碳含量均随土层深度的增加呈降低趋势,其中40—50 cm,50—60 cm土层有机碳含量及储量差异不显著(p>0.05)。由表3还可知,表层0—10 cm土壤有机碳储量最高,达到2.19~2.89 kg/m2,已远远大于植被有机碳储量,0—10 cm的浅层土壤中,NG和HG土壤碳储量差异不明显(p<0.05),MG和LG表现出较强的储碳能力,10—20 cm呈现出显著降低的特征;从0—20 cm的土壤总碳储量来看,该深度范围内的碳储量占到0—60 cm碳储量的30%~40%,碳储量大小序列为MG>LG>NG>HG,MG和LG的土壤储碳优势逐渐表现出来,MG表现出强劲的固碳能力,其碳储量显著高于其他,而在这一深度范围,HG碳储量亦较低;从0—60 cm土层深度来看,碳储量表现出随着退化程度的加重而增加的趋势,NG土壤碳储量显著下降,不同退化程度之间均存在显著性差异(p<0.05),其中MG的固碳能力最强,其余依次为LG,NG和HG,HG相对于NG土壤有机碳储量显著减少,降幅为6.23%。由此可以看出,适度放牧有助于土壤有机碳的积累。
2.6 放牧强度对生态系统有机碳储量的影响
由图3可知,不同放牧强度典型草原土壤有机碳储量大小序列为MG>LG>NG>HG,植被有机碳储量大小序列为NG>LG>MG>HG,生态系统有机碳储量大小序列为MG>LG>NG>HG,与土壤有机碳储量保持一致的变化规律,由此说明放牧对植被—土壤系统有机碳储量的影响具有非同步性,轻度和中度放牧有助于生态系统有机碳储量的积累,而重度放牧草原生态系统有机碳储量显著降低。随着放牧强度的增加,地上植物碳储量占生态系统有机碳储量比例分别为17.85%,15.68%,13.01%,15.29%,土壤碳储量所占比例分别为82.15%,84.32%,86.99%,84.71%。土壤碳贮量占总碳贮量的80%以上,是草原碳储量的主体部分,影响着整个生态系统有机碳储量的变化。各样地各部分所占生态系统有机碳储量比例均表现为土壤>根系>地上植物>凋落物。

表3 放牧强度对根系碳储量的影响

图3放牧强度对生态系统有机碳储量的影响
2.7 土壤碳储量与植被碳储量及土层深度的关系
本研究分析了植被地上生物量、地下生物量(0—60 cm)与土壤有机碳储量的相关性,以及不同放牧强度下,草原土壤不同深度的碳储量与相应深度的植被地下生物量关系(表4)。植被地上碳储量与0—20 cm土壤碳储量存在着显著的相关性(p<0.05),其相关系数随着土层深度的增加而降低,以MG的相关系数绝对值最高,而20—40 cm以下,植被地上碳储量与土壤碳储量没有显著的相关性(p>0.05)。
上述研究表明,不同放牧强度下草原土壤有机碳的垂直分布呈现出了较好的一致性变化,即随着土层深度的增加土壤有机碳储量呈递减趋势。为了定量描述这种趋势,本研究利用回归分析方法,建立土层深度与土层有机碳储量的回归方程(表4)。对于不同放牧强度下,线性方程、对数方程与一元二次方程的效果都比较好,且对数方程和二次方程效果略优于线性方程。另外,研究还将4组数据综合,建立通用方程,发现3类方程的效果也是可以接受的。总之,回归分析结果表明土壤有机碳储量与土层深度具有较好的相关关系。

表4 放牧强度对根系碳储量的影响
注:*表示显著相关(p<0.05),**表示极显著相关(p<0.01)。

表5 土壤有机碳储量与土层深度的回归分析
注:y为土壤有机碳储量,x为土层深度。
3 讨论与结论
天然草原的生产从大尺度上来讲,主要取决于当地的气候条件、地形特征和土壤特性等环境因子,但对同一块草原而言,这些因素的影响是恒定不变的,那么放牧是影响草原群落特征最主要的因子。放牧对草原的影响,首先表现在草原植物的变化,地上生物量是反映草原生态系统、放牧系统稳定性的重要指标,其大小可判断草原状况、生产潜力和载畜能力等,在放牧胁迫下,群落组分在放牧中有直接的损失,影响植物群落的生产力[18]。本研究表明,随放牧强度的增加,群落地上生物量呈现出降低的变化趋势,由此看来,随着放牧强度的增强,家畜的频繁啃食降低了牧草的生物量,影响其物质能量的积累,进而影响植物的生长发育和繁殖,造成群落中物种个体普遍减少。另外,家畜的踩踏也影响了草原的土壤特性,这也引起草原植物群落生产力的降低。这与段敏杰[19]、董全民[20]等在天然草原的研究结果一致。不同功能群植物地上生物量和占群落生物量的比例变化主要受放牧过程中放牧家畜选择性采食的影响。放牧过程中,家畜优先选择采食优良牧草(禾草和莎草),尤其是禾草,使禾草原上生物量减小。主要是因为禾草相对高度较高,处于草群上层,家畜优先采食,同时抑制了其生长发育。上层高大植物的减少,使草层下部能接受到阳光照射,光合作用增强,促进了物质能量的积累。且受到上层禾草郁蔽环境的影响减小,增强了自身的补偿作用和放牧抵抗力,比例增加;杂类草和毒草是适口性差或不能食用的,同时上层高大草类的减少降低了对其的抑制作用,故随放牧强度的增加,比例增加[21]。
植物多样性对植物群落稳定性的影响是通过不同功能群间的补偿作用实现的,在牧压梯度下,物种和植物功能群间的这种补偿作用尽管有时是十分有限的,但对维持生态系统结构与功能过程的相对稳定性却具有极其重要的作用[22]。本研究表明,随着放牧强度的增加,物种丰富度和多样性指数基本都呈现出单峰变化,即在中度放牧区多样性最高,虽然均匀度指数的变化趋势较复杂,但在中度放牧的均匀度指数高于重度放牧。这是因为通过适度的放牧干扰,家畜对植物的采食作用,抑制了优势种的竞争能力,使弱势物种的入侵和定居成为可能,一些下繁草的物种数量增加,同时家畜不喜食和不采食的杂类草和毒草类的数量也增加,增加了群落结构的复杂性,群落内物种的多样性出现一定程度的增加。这在一定程度上支持了“中度干扰假说”[23]。即干扰对资源和环境异质性的作用表现为非线性,适中水平的干扰增加物种多样性,而超过一定阈值的干扰又会降低物种多样性。重度放牧下群落物种丰富度、多样性指数和均匀度指数都表现为最低,当放牧压力过大时,群落内的可食性牧草因家畜啃食过度而失去再生能力,以致在群落中消失,加之不具可食性的毒杂草在群落中所占的比例增加,而且由于家畜的过度采食,降低了有机质向土壤中的输入,土壤营养过度消耗,降低了植物的竞争能力,使草原植物群落中的物种多样性下降。
植被是土壤有机碳的主要输入来源,在放牧退化演替过程中,植被演替的短期效应较为明显,而土壤作为一个相对稳定的系统,其变化较为缓慢,尤其是土壤有机碳的积累是长期结果,这就致使土壤有机碳储量与植被状况的关系变得复杂而不确定[24]。不同干扰强度主要是通过家畜采食改变地上部分,降低植物地上生物量(图2);过度放牧,使植物叶面积降低,草原初级生产固定碳素的能力下降,光合作用向地下的能量输入减弱,导致植物地下根系的生产力和现存量降低;草原生物量降低使凋落物的积累和输入降低,减少了碳素由植物凋落物向土壤的输入,降低土壤有机碳含量,使土壤碳储量降低,而适度放牧有利于草原有机碳的蓄积,因为它可以减少凋落物的积累,促进其分解并加速碳向土壤的输送[24]。不同放牧强度草原土壤有机碳储量大小序列为MG>LG>NG>HG,轻度和中度放牧提高了生态系统有机碳水平。导致这一变化的原因可能归因于不同放牧强度下根冠比、凋落物分解及地上生物量的差异;一方面,放牧强度的加大导致植物地下生物量增加,使得植物根冠比率增加,增加了有机碳向地下的分配量[25];另一方面,动物的践踏使凋落物破碎并与土壤充分接触,促进凋落物的分解,有助于碳和养分元素转移到土壤中而重度放牧促使草原植被叶面积和储藏物质严重减少,净初级生产力降低,物质的输入小于输出,导致生态系统碳储量降低。Frank等[26]指出放牧会抑制土壤侵蚀的发生,从而保护了土壤有机碳含量。食草动物排泄物的归还也使土壤表层速效养分增加,土壤的矿化作用加强。此外,放牧使植物的组成发生变化,导致了低的产草量,增加了碳向地下的分配量。放牧还使凋落物积累量减少,这是由于动物的践踏使凋落物破碎并与土壤充分接触,有助于凋落物的分解,也有助于碳和养分元素转移到土壤中。
在生态系统碳储量中,有机碳储量从大到小依次为:土壤有机碳储量、根系有机碳储量、地上植物有机碳储量和凋落物有机碳储量,由此可以看出,不同放牧强度羊草生态系统有机碳储量中土壤有机碳储量占居主导地位,这一结果与对典型草原和草甸草原生态系统碳储量研究所得到的结果相似[3-5]。不同放牧强度草原0—10 cm土壤有机碳储量均为最高,这一方面是由于地上植物的枯萎、凋落物沉积在土壤表层,促进了表层土壤有机碳的输入,另一方面归因于草原地下生物量大部分集中于表层土壤中,随着深度增加,数量急剧降低[25,27]。随着放牧强度的增加,地上植物、凋落物、根系有机碳储量所占生态系统有机碳储量比例呈现降低趋势,而土壤有机碳储量所占比例呈递增趋势,0—40 cm土壤碳储量占到系统总量的80%以上,说明草原碳储量主要储存在土壤当中,如李怡[27]指出,针茅草原地上植被碳储量仅占总碳储量的2%~5%,其余大部分碳储存在土壤中,土壤表现出了碳汇的特征,如果大范围进行科学保护与适当利用,其碳固持能力,也即草原的碳汇功能将得到充分的发挥,其固碳潜力巨大。另外,很多研究对土壤碳垂直分布做出了定性判断,认为草原生态系统土壤有机碳随着深度的增加而减少,土壤有机碳的垂直分布格局受凋落物量、淋溶作用、植物根系分布及活动特征和微生物活动等多种因素的影响,在典型草原地区随着土层深度的增加,土壤温度、含水量和质地条件变差,有机质来源减少,因此出现了有机碳随土层深度的增加而表现出逐渐降低的趋势[23,27]。本研究发现,土壤有机碳储量与深度之间存在着显著的相关性,而且这一关系可以用线性、对数和二次函数来定量描述。通过本研究认为,面临当前草原退化严重的现状,实施围封禁牧具有重要意义,但是,在退化草原的恢复过程中要加强对围封不干扰生态系统的监测,同时,要科学认识放牧利用对维持草原生态系统健康的重要性及特殊意义,适时采取相应措施,发挥适度放牧利用对退化草原碳积累和恢复进程的优化调控作用。
致谢:特别感谢中科院内蒙古锡林郭勒草原生态系统国家野外科学观测研究站所有工作人员和研究人员在本研究中提供的帮助和支持。
参考文献:
[1] Dick M, Silva M A D, Dewes H. Life cycle assessment of beef cattle production in two typical grassland systems of southern Brazil[J]. Journal of Cleaner Production, 2015,96:426-434.
[2] Zhu X, Yu G, Wang Q, et al. Seasonal dynamics of water use efficiency of typical forest and grassland ecosystems in China[J]. Journal of Forest Research, 2014,19(1):70-76.
[3] Shi Y, Wang Z Q, Zhang X Y, et al. Effects of nitrogen and phosphorus addition on soil microbial community composition in temperate typical grassland in Inner Mongolia[J]. Acta Ecologica Sinica, 2014,34:4943-4949.
[4] Han D, Wu P, Zhang Q, et al. Feature extraction and image recognition of typical grassland forage based on color moment[J]. Transactions of the Chinese Society of Agricultural Engineering, 2016,32(23):168-175.
[5] 柴曦,梁存柱,梁茂伟,等.内蒙古草甸草原与典型草原地下生物量与生产力季节动态及其碳库潜力[J].生态学报,2014,34(19):5530-5540.
[6] 王晶,王珊珊,乔鲜果,等.氮素添加对内蒙古退化草原生产力的短期影响[J].植物生态学报,2016,40(10):980-990.
[7] 于丽,赵建宁,王慧,等.养分添加对内蒙古贝加尔针茅草原植物多样性与生产力的影响[J].生态学报,2015,35(24):8165-8173.
[8] Genet H, Mcguire A D, Barrett K, et al. Modeling the effects of fire severity and climate warming on active layer thickness and soil carbon storage of black spruce forests across the landscape in interior Alaska[J]. Environmental Research Letters, 2013,8(4):045016.
[9] Torn M S, Trumbore S E, Chadwick O A, et al. Mineral control of soil organic carbon storage and turnover[J]. Nature, 1997,389(6647):170-173.
[10] Poeplau C, Marstorp H, Thored K, et al. Effect of grassland cutting frequency on soil carbon storage:A case study on public lawns in three Swedish cities[J]. Soil, 2016, 2(2):1-23.
[11] Schwendenmann L, Pendall E. Effects of forest conversion into grassland on soil aggregate structure and carbon storage in Panama: Evidence from soil carbon fractionation and stable isotopes[J]. Plant and Soil, 2006,288(1):217-232.
[12] Reid J P, Adair E C, Hobbie S E, et al. Biodiversity, Nitrogen Deposition, and CO2Affect Grassland Soil Carbon Cycling but not Storage[J]. Ecosystems, 2012,15(4):580-590.
[13] Milchunas D G, Lauenroth W K, Chapman P L, et al. Effects of grazing, topography, and precipitation on the structure of a semiarid grassland[J]. Plant Ecology, 1989,80(1):11-23.
[14] Harrison S, Inouye B D, Safford H D. Ecological Heterogeneity in the Effects of Grazing and Fire on Grassland Diversity[J]. Conservation Biology, 2003,17(3):837-845.
[15] Bullock J M, Hill B C, Dale M P, et al. An experimental study of the effects of sheep grazing on vegetation change in a species-Poor grassland and the role of seedling recruitment into gaps[J]. Journal of Applied Ecology, 1994,31(3):493-507.
[16] Masiello C A, Chadwick O A, Southon J, et al. Weathering controls on mechanisms of carbon storage in grassland soils[J]. Global Biogeochemical Cycles, 2004,18(4):449-460.
[17] Bakker E S, Ritchie M E, Olff H, et al. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. [J]. Ecology Letters, 2006,9(7):780-789.
[18] Gustavsson E, Lennartsson T, Emanuelsson M. Land use more than 200 years ago explains current grassland plant diversity in a Swedish agricultural landscape[J]. Biological Conservation, 2007,138(1/2):47-59.
[19] 段敏杰.放牧干扰下藏北紫花针茅高寒草地生物量遥感监测[D].北京:中国农业科学院,2011.
[20] 董全民,赵新全,马玉寿,等.放牧强度对高寒混播人工草地群落特征及地上现存量的影响[J].草地学报,2012,20(1):10-16.
[21] An Hui, Li Guoqi. Effects of grazing on plant biomass and soil nutrient in desert steppe[J]. Journal of Plant Nutrition & Fertilizer, 2013,19(3):705-712.
[22] May F, Grimm V, Jeltsch F. Reversed effects of grazing on plant diversity: The role of below-ground competition and size symmetry [J]. Oikos, 2009,118(12):1830-1843.
[23] 刘艳红,赵惠勋.干扰与物种多样性维持理论研究进展[J].北京林业大学学报,2000,22(4):101-105.
[24] Gang C, Zhang J, Li J. The Advances in the carbon source/sink researches of typical grassland ecosystem in china[J]. Procedia Environmental Sciences, 2011,10:1646-1653.
[25] Xiong D, Shi P, Sun Y, et al. Effects of grazing exclusion on plant productivity and soil carbon, nitrogen storage in alpine meadows in northern tibet, china[J]. Chinese Geographical Science, 2014,24(4):488-498.
[26] Frank D A, Mcnaughton S J, Tracy B F. The Ecology of the Earth′s Grazing Ecosystems[J]. BioScience, 1998,48(7):513-521.
[27] 李怡,韩国栋.放牧强度对内蒙古大针茅典型草原地下生物量及其垂直分布的影响[J].内蒙古农业大学学报:自然科学版,2011,32(2):89-92.
猜你喜欢
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
矿产勘查(2020年4期)2020-12-28
矿产勘查(2020年2期)2020-12-28
今日农业(2020年19期)2020-12-14
石油与天然气地质(2020年4期)2020-08-14
安徽农学通报(2020年7期)2020-05-26
中国矿业(2019年8期)2019-08-16
中国农业文摘-农业工程(2016年5期)2016-04-12
中国科技信息(2015年2期)2015-11-16
