
60Co-γ辐射诱变处理对无芒雀麦种子萌发及幼苗期抗旱性的影响
2018-05-17 04:15李祥莉
干旱地区农业研究 2018年2期
李 波,李祥莉
(齐齐哈尔大学生命科学与农林学院, 黑龙江 齐齐哈尔 161006)
随着水资源的日益匮乏,干旱早已成为全球内最受关注的环境问题,也是影响植物生长和产量的重要因素之一。近年来我国相继遭遇严重的秋旱和伏旱,其旱情具有持续时间长、发展速度快等特点,对牧草生产造成了很大的影响。因此,筛选具有较强耐干旱的牧草是实现环境绿化和干草饲料最实用的途径。
无芒雀麦(Bromes inermis Leyss)又名光雀麦,属于多年生雀麦属,是水土保持最好的冷季型的禾本科牧草,原产于亚洲、欧洲和北美洲等寒冷地区的一种重要的栽培牧草,多分布于山坡、道旁、河岸。具有叶量丰富,营养价值高,适口性好,消化率高,抗寒抗旱,适度放牧,再生性能力强,各种家畜均喜采食等优良品质,是建立人工草地和优良水保的重要草种[1]。
60Co-γ辐照具有能够提高突变率,后代遗传稳定,缩短育种周期等优点,是获得新种质资源的有效途径[2-3]。近年来辐射诱变技术主要集中在粮食和经济作物上,已成为当代植物改良和作物育种的一种重要手段。目前,对无芒雀麦的研究方向在种间关系、产量、资源培育等方面[4],迄今关于无芒雀麦种子耐旱方面的研究较少。聚乙二醇(PEG)可用于鉴定不同植物种子萌发期的耐旱性研究[5]。种子发育成苗是生活史上最为脆弱的发育时段,开展种子萌发期抗旱性,对干旱环境适应性及响应机制研究尤为重要。本试验采用PEG-6000高渗溶液模拟干旱胁迫,初步确定无芒雀麦适宜的胁迫浓度,再通过不同剂量的60Co-γ射线辐射对无芒雀麦的多项指标进行抗旱性综合分析,为无芒雀麦种子的萌发和幼苗生长的耐旱性研究提供理论依据。
1 材料与方法
1.1 材 料
供试的无芒雀麦种子由黑龙江畜牧研究所提供,利用烘干机烘干后,室温干燥器内保存。
1.2 试验设计
1.2.1 辐照处理 以60Co-γ 射线作为辐射源,2016年5月在中国农业科学院原子能研究所对无芒雀麦干种子进行辐射处理,辐射剂量为 50、100、150、200、250 Gy和300 Gy,剂量15 Gy·min-1,每个处理100 g种子,以未经过处理的种子作对照。
1.2.2 PEG胁迫浓度的设置 采用PEG-6000模拟不同浓度的PEG处理液,共设6个胁迫浓度,质量分数分别为0、5%、10%、15%、20%和25%,与之相对应的溶液水势分别为0、-0.05、-0.20、-0.40、-0.60 MPa和-0.86 MPa[6]。选取均匀、饱满的种子30粒置于发芽盒内浸透胁迫溶液的滤纸上进行发芽,3次重复。待发芽结束,根据发芽率来推断出种子最适宜的胁迫浓度。
1.2.3 种子萌发试验 选取各组的种子若干粒,用35℃蒸馏水浸泡2 h,用吸水纸吸干种子表面的水分,再将种子均匀排布于发芽盒内浸透PEG胁迫浓度为10%的滤纸上,置于光照培养箱中培养,培养条件为:25℃恒温,光照强度120 μmol·m-2·s-1,光照时间12 h·d-1,为了防止发芽盒中水势的变动,每3 d更换一次滤纸,并添加20 ml原液,培养幼苗。
1.3 指标测定及方法
待发芽结束后(10 d),用滤纸吸干10株幼苗上的水分称其鲜质量,用游标卡尺分别测定根长和苗高,采用烘干法测其干质量,取平均值。
胚芽长度(L芽)=(L1+L2+L3+……+L10)/10; 胚根长度(L根)=(L1+L2+L3+……+L10)/10
称取0.1 g鲜样进行抗旱相关指标测定,细胞膜透性采用相对电导率法,可溶性蛋白含量采用考马斯亮蓝法,可溶性糖含量采用硫代巴比妥酸法,游离脯氨酸含量采用酸性茚三酮法;超氧化物歧化酶(SOD)活性采用淡蓝四唑法,过氧化物酶(POD)活性采用愈创木酚氧化法,过氧化氢酶(CAT)活性采用紫外吸收法[7],上述每项指标均做3次重复。
1.4 数据分析
采用Excel进行数据统计处理,SPSS17.0软件进行单因素方差(ANOVA)检验分析,试验数据取3次数据的平均值,对数据进行方差统计分析。
对其抗逆性的综合评价采用隶属函数法[8],由公式计算与抗逆性的关系的指标的具体隶属值。应用隶属法对不同处理的无芒雀麦幼苗的抗逆性进行综合评价。
隶属函数值计算公式:R(Xi)=(Xi-Xmin) /(Xmax-Xmin),反隶属函数值计算公式:R(Xi)=1-(Xi-Xmin) /(Xmax-Xmin)(式中,Xi为指标测定值,Xmin、Xmax为所有实验材料某项指标的最小值和最大值)。将抗性隶属值进行累加求出平均数,如公式:X=∑U(Xi)/n,X是所求平均抗性的隶属值,其中隶属值越大的其抗逆性越强。
2 结果与分析
2.1 PEG模拟干旱胁迫适宜浓度的确定
在PEG胁迫环境下,适宜胁迫浓度的选择是前提依据。实践中多以LD50(发芽率降低50%的剂量)作为判断作为种子胁迫适应浓度的指标[9]。利用回归方程求得发芽率的半致死胁迫浓度见图1,因此,综合分析初步确定无芒雀麦种子的PEG胁迫浓度为10%。


图1 种子发芽率与胁迫浓度的关系
图2 PEG胁迫10 d后幼苗
Fig.2 Shoot after 10 days under PEG stress
2.2 60Co-γ辐照对PEG胁迫下无芒雀麦苗高和根长的影响
种子萌发后,幼苗和胚根的延伸反映出植物成苗的特性, PEG胁迫浓度为10%时,不同辐射剂量下无芒雀麦幼苗生长状况见图2。由表1可知,PEG胁迫下,种子的苗高、根长与辐射剂量呈负相关关系。在胁迫处理结束(10 d)后,低剂量辐射(50~100 Gy)种子苗高高于对照,分别为对照的0.23和0.11倍,高剂量辐照(150~300 Gy)低于对照,分别为对照的0.14、0.33、0.57和0.77倍,与对照差异显著(P≤0.05),且辐射剂量在50 Gy和300 Gy时达到最大值和最小值。低剂量辐射种子根长高于对照,为对照的0.21和0.12倍,高剂量辐照(150~300 Gy)低于对照,为对照的0.33、0.58和0.78倍,与对照差异显著(P≤0.05)。
2.3 60Co-γ辐照对PEG胁迫下无芒雀麦幼苗生物量及含水量的影响
由表1可知,PEG胁迫下无芒雀麦幼苗,随着辐射剂量的不断增加,幼苗鲜重和干重均有一定程度增加。低剂量辐射(50 Gy和100 Gy)高于对照,与对照相比分别增加了34.18%和14.14%,与对照相比差异显著(P≤0.05),干重增加了4.90%和1.80%,且与对照相比无显著差异(P≥0.05)。含水量直接影响植物的生长、营养状况及产量,很容易受到外界因素的影响。低剂量60Co-γ辐照后,含水量有所增加,分别比对照增加了37.44%和15.5%,且与对照相比差异显著(P≤0.05)。
2.4 60Co-γ辐照对PEG胁迫下无芒雀麦幼苗细胞膜透性的影响
细胞膜是一种选择透过性膜,它能够控制和调节植物内外的交换和运输,其透性是评价植物对逆境反应的指标之一。细胞膜透性与辐射剂量呈正相关(见表1),低剂量辐照(50~100 Gy)低于对照,而高剂量辐照(150~200 Gy)高于对照,分别增加了0.49%和1.47%,说明高剂量辐射对细胞膜产生了一定的损伤,导致细胞内物质的外渗。

表1 60Co-γ辐照对干旱胁迫条件下无芒雀麦苗高、根长、生物量和含水量及细胞膜透性的影响
注:表中数据为3次重复的平均值;同列不同字母表示处理间在P<0.05水平有显著差异。
Note: The data are averages of three replicates; letters within the same column with different letter mean significant differences atP<0.05 below.
2.5 60Co-γ辐照对PEG胁迫下无芒雀麦幼苗渗透调节物质的影响
图3表明,60Co-γ辐照对PEG胁迫下无芒雀麦幼苗渗透调节物质含量的变化趋势均呈现先升后降,剂量在100 Gy以下均可促进渗透调节物质含量的增加。剂量在100 Gy以下时脯氨酸含量均高于对照,与对照相比上升了24.54%和23.03%,随之下降,剂量在200 Gy时达到最小值,与对照相比下降了34.58%,且与对照差异显著(P≤0.05)。剂量在100 Gy以下均可促进可溶性糖的含量,与对照相比分别提高了7.4%和34.35%。剂量在150~200 Gy时,可溶性糖含量呈下降趋势且均小于对照,与对照相比下降了9.6%和50.34%。蛋白质的代谢受多种因素的影响,包括PEG胁迫在内的非正常环境条件都会影响细胞内的可溶性蛋白质。剂量在150 Gy以下,可溶性蛋白含量均高于对照,与对照相比上升了9.3%、21.67%和20.9%,剂量在200 Gy时降到最小,与对照相比降低了19.64%,且与对照无显著性差异(P≥0.05)。说明高剂量辐射对渗透调节物质含量具有明显的抑制作用。

注:不同处理不同字母表示差异显著(P<0.05)。下同。
Note:Different treatments with the different letters mean s significant different by Duncan’s testP<0.05. The same below.
图360Co-γ辐射对PEG胁迫下无芒雀麦幼苗脯氨酸、可溶性糖和可溶性蛋白含量的影响
Fig.3 The proline、soluble sugar and soluble protein of brome seedling under PEG stress by60Co-γ-rays
2.6 60Co-γ辐照对PEG胁迫下无芒雀麦幼苗保护酶系活性的影响
由图4可以看出,无芒雀麦幼苗CAT活性随着辐射剂量的增加呈现先升后降趋势,剂量在100 Gy时,CAT活性达到最大值,与对照相比提高了11.82%。剂量为200 Gy时显著降低了萌发期的CAT活性,较对照下降了38.83%,与对照间差异显著(P≤0.05)。POD的变化同CAT变化趋势相似,随着辐射剂量的增加,POD活性呈先升后降趋势,剂量在50~100 Gy范围内,POD活性均高于对照,较对照分别提高了9.70%和19.72%,且与对照无显著性差异(P≥0.05)。辐射剂量为200 Gy时,POD活性达到最低,较对照下降了35.45%,且与对照差异显著(P≤0.05)。各辐射剂量的SOD活性的变化趋势呈现先升后降,剂量在50~150 Gy范围内,SOD活性均高于对照,较对照分别提高了44.49%、25.16%和12.63%,与对照差异显著(P≤0.05),且剂量在50 Gy时SOD活性升到最大。辐射剂量为200 Gy时,SOD活性降低到最小,较对照下降了17.83%,且与对照无显著性差异(P≥0.05)。由此可见,PEG模拟水分胁迫下,60Co-γ射线辐射的无芒雀麦种子可以通过增加体内保护酶活性来提高抵抗干旱环境的能力。
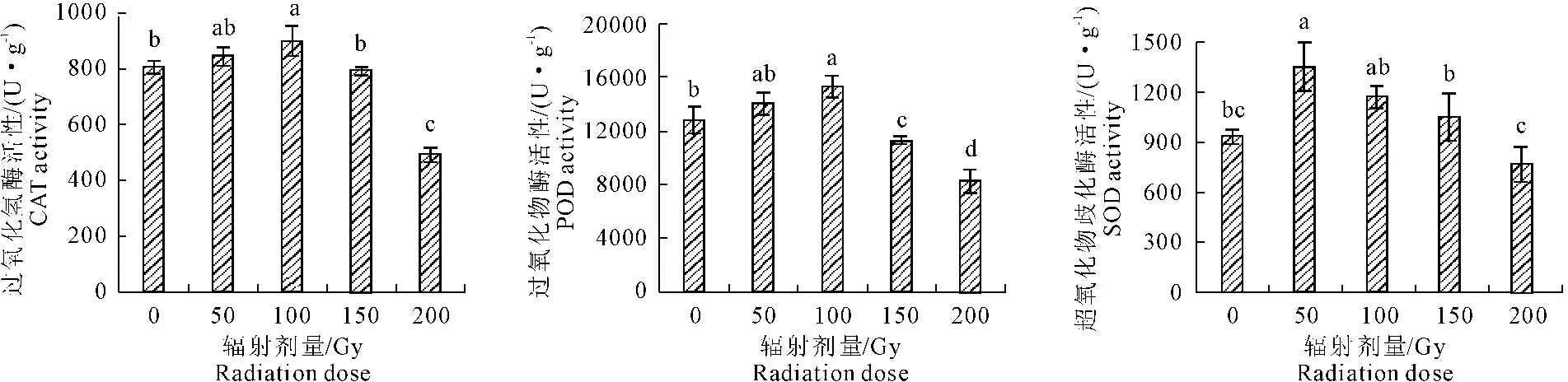
图460Co-γ辐射对PEG胁迫下无芒雀麦幼苗CAT、 POD和SOD活性的影响
Fig.4 The CAT、POD and SOD of brome seedling under PEG stress by60Co-γ radiation
2.7 60Co-γ辐照对PEG胁迫下无芒雀麦幼苗抗旱性综合分析
60Co-γ辐照对PEG胁迫下无芒雀麦幼苗抗旱性的影响反映在幼苗的生长、渗透调节物质、膜脂氧化和抗氧化酶的变化。各指标中存在着一定的相关性和差异性,在具体进行植物耐旱性评价时需要综合考虑。辐照对PEG胁迫下无芒雀麦幼苗在苗高、根长、鲜质量、干质量、含水量、细胞膜透性、可溶性糖、可溶性蛋白、脯氨酸、过氧化物酶、过氧化氢酶和 超氧化物歧化酶的变化不同。因此应用单一指标不能准确反映不同60Co-γ辐照剂量对无芒雀麦幼苗抗旱能力的影响,需要用多种指标进行综合评价,将12项指标的数据代入隶属函数公式中计算出隶属函数值,其隶属函数值见表2,隶属函数综合评价值越大说明其抗旱能力越强。4种辐射剂量下抗旱的强弱顺序为:50 Gy>10 Gy>150 Gy>200 Gy,50~100 Gy辐射剂量能有效提高无芒雀麦的抗旱能力。
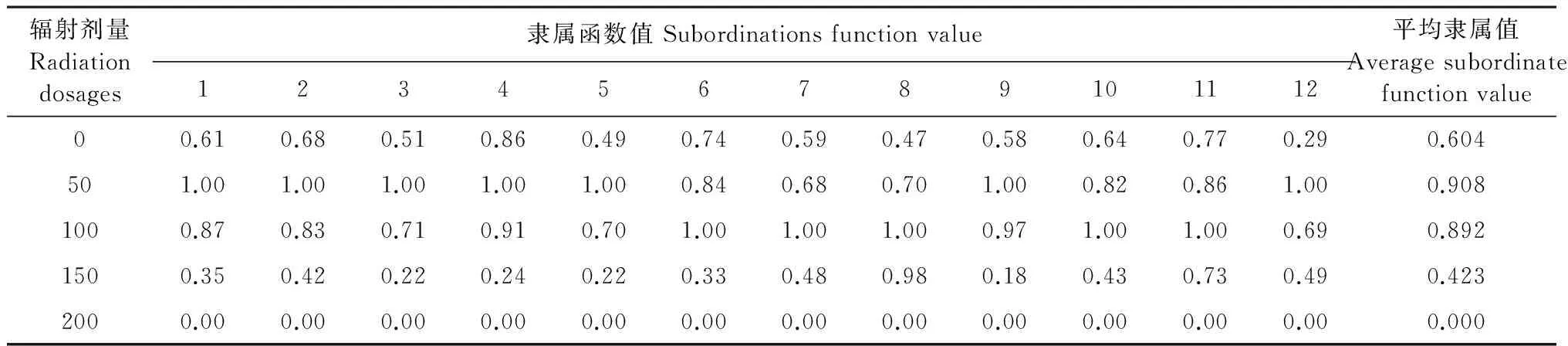
表2 60Co-γ辐照对干旱胁迫条件下无芒雀麦幼苗12项指标的隶属值
注:1,2,3,4,5,6,7,8,9,10,11,12为各指标隶属值,包括有苗高,根长,鲜质量,干质量,含水量,细胞膜透性,可溶性糖,可溶性蛋白,脯氨酸,过氧化物酶,过氧化氢酶,超氧化物歧化酶的隶属值。
Note: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 is the subordinate value of each index,including shoot length, root length, fresh weight, dry weight, water content, cell membrane, soluble sugar, soluble protein, proline, POD, CAT and SOD.
3 讨 论
种子萌发是旱生植物生活史中对土壤干旱十分敏感而关键的阶段。在PEG环境中,种子能够萌发是植物生长发育的前提。一般而言,随着辐射剂量的增加,突变频率也会随之提高,但过高的辐射剂量会干扰生物体正常生长代谢能力,从而增加畸变率。辐射诱变首先要确定适宜的辐射剂量[10]。对于不同植物来讲,在PEG胁迫下60Co-γ辐照的反应机理不同,从而出现不同的作用机制。PEG为10%胁迫下,无芒雀麦在不同剂量的抗旱性不同,综合分析最适宜的辐射剂量为50~100 Gy。
根系是水分胁迫下最敏感的部位,根系的生长发育状况直接影响着其生长与产量的形成。PEG胁迫下,低剂量辐射促进种子的根系、幼苗生长,从而更好的维持植物中的水分,可能适宜的辐射剂量迫使同化物更多的配合根系的分配,促进了根系的分枝和下扎,这与李文鹤[11]对野菊的研究结果相似。而高剂量辐射严重抑制植物的生根及生长,导致植物幼苗几乎不生长。
60Co-γ辐照对PEG胁迫下无芒雀麦幼苗的生物量和含水量产生不同影响。低剂量辐射提高了植物体内的水分,减缓胁迫对植物造成的负面影响,促进了植物对水分和无机养料的吸收,使其鲜重和干重增加。随着辐射剂量的逐渐增加,无芒雀麦幼苗的鲜重、干重及含水量均呈现下降趋势,说明高辐射抑制植物对水分和无机养料的吸收,导致鲜重和干重均下降。一般认为在干旱环境下能维持较高含水量的品种具有较强的抗旱性[12],Kaya[13]等也有类似结果,即能增加玉米叶片的含水量。
当植物受到辐照和胁迫时细胞内活性氧量增加,导致细胞结构改变,膜系统紊乱,进而影响植物的生长发育。渗透调节在植物抗旱中起着至关重要的作用,可通过提高细胞液浓度、降低植物体的渗透势等方法来维持植物正常的生理代谢[14-15]。可溶性蛋白能够维持细胞内的渗透势、脯氨酸、可溶性糖含量在一定程度上反映了植物的抗逆性,抗性强的品种往往积累较多的渗透调节物质。研究结果表明,渗透调节物质含量的变化趋势与辐射剂量呈负相关,与张玉[16]、滕娟[17]等人研究结果相似。在逆境条件下,积累这些物质以适应外界环境的变化,对植物具有保护作用,从而提高植物的抗逆适应性。高剂量60Co-γ射线辐照对PEG胁迫下无芒雀麦幼苗细胞膜透性有一定的破环作用,导致细胞膜透性增加。由此可见,辐射剂量在100 Gy以下时无芒雀麦能够积累较多的渗透调节物质,且膜未受到损伤,可抵御干旱环境维持正常的生理代谢,而高剂量辐射不能维持正常的代谢,甚至破坏了膜结构的完整性。
植物在辐射条件下,活性氧自由基的产生能力增强,酶促活性氧自由基清除系统开启[18]。本研究中,无芒雀麦种子3种保护酶(SOD、POD、CAT)活性在干旱环境下辐射响应不同,随辐射剂量的增加整体变化趋势呈先升后降。这与郑宾国[19]的研究结果一致,辐射剂量在100 Gy时,SOD、POD达到最大值,随之下降,说明低剂量辐射未使得SOD、POD酶活性系统受到损坏,可以清除体内过多的氧化性物质,因此酶活性上升。但辐射剂量过大时,SOD、POD酶活性系统受到损害,逐渐失去抵御氧化性物质的能力。这与何玮对甜高粱的研究结果相符。CAT活性可作为植物在其的指示性指标,其变化较为敏感。结果显示,低剂量辐射条件下,其幼苗主要靠3种保护酶协同作用下,共同消除自由基。但随着剂量的增加,活性氧积累量的增加致使部分酶失活,导致三种酶活性同时出现下降的趋势。
本研究从多项指标综合分析无芒雀麦种子萌发期抗旱性,但由于植物在不同生长阶段抗旱性不尽相同,干旱对60Co-γ辐射无芒雀麦萌发期研究,不能完全代表田间育苗实际情况,还要进一步进行田间苗期耐旱实验。本试验为利用60Co-γ射线辐射来提高无芒雀麦的耐旱性提供了理论依据。
参考文献:
[1] 杨慧玲,曹志平,董 鸣,等.沙埋对无芒雀麦种子萌发和幼苗生长的影响[J].应用生态学报,2007,18(11):2438-2443.
[2] 张 雪,刘青松,师文贵,等.25份无芒雀麦种质资源遗传多样性的ISSR分析[J].中国草地学报,2014,36(3):104-107.
[3] Song J, Ren M, Song C, et al. The effect of60Coγ-rays on the crystal structure, melting and crystallization behavior of poly(butylene succinate[J]. Polymer International, 2011,53(11):1773-1779.
[4] 陈积山,朱瑞芬,高 超,等.苜蓿和无芒雀麦混播草地种间竞争研究[J].草地学报,2013,21(6):1157-1161.
[5] Shuya Wei, Wei Hu, Xiaomin Deng. A rice calciun-dependent protein60Co-γ positively regulates drought stress tolerance and spikelet fertility[J]. BMC Plant Biology, 2014,(14):133-136.
[6] 张 娜,赵宝平,张艳丽,等.干旱胁迫下燕麦叶片抗氧化酶活性等生理特性变化及抗旱性比较[J].干旱地区农业研究,2013,31(1):166-171.
[7] 王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[8] 王婧泽,高树仁,孙丽芳,等.3个玉米自交系对盐胁迫的生理响应及耐盐性评价[J].干旱地区农业研究,2017,35(2):89-95.
[9] 熊秋芳,陈玉霞,张雪清,等.60Co-γ辐射对萝卜种子萌发和幼苗生长的影响[J].浙江农业科学,2014,(3):356-359.
[10] Ansari O, Chogazardi H R, Sharifzadeh F, et al. Seed reserve utilization and seedling growth of treated seeds of mountain rye (Secale montanum) as affected by drought stress[J]. Cercetari Agronomice in Moldova, 2012,45(2):43-48.
[11] 李文鹤,何 淼,卓丽环.PEG-6000处理对野菊种子萌发期抗旱性的影响[J].种子,2010,29(11):51-54.
[13] Kaya C, Tuna L, H. Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress condition[J]. Journal of Plant Nutrition, 2006,29(8):1469-1480.
[14] 高 睿,马 淼,赵红艳.60Co-γ辐射对光果甘草种子耐盐性影响的研究[J].农学学报,2016,6(7):77-82.
[15] Munns R. Comparative physiology of salt and water stress[J]. Plant Cell and Environment, 2002,25(2):239-250.
[16] 张 玉,白史且,李达旭,等.60Co-γ辐射对菊苣种子发芽及幼苗生理的影响[J].草地学报,2013,21(1):147-151.
[17] 滕 娟,熊俊芬,何忠俊,等.60Co-γ辐射对三七幼苗生理特性的影响[J].云南农业大学学报自然科学,2015,30(3):445-449.
[18] 李 波,徐婉玉,彭 丹,等.卫星搭载苜蓿种子叶片愈伤组织对逆境胁迫的响应[J].干旱地区农业研究,2015,33(6):167-171.
[19] 郑宾国,崔节虎,彭伟功,等.不同γ-射线辐照剂量对鱼腥藻生长及生理特性的影响[J].西北农林科技大学学报自然科学版,2013,41(8):182-188.
猜你喜欢
作物研究(2022年4期)2022-09-21
中国临床医学影像杂志(2022年6期)2022-07-26
昆明医科大学学报(2021年8期)2021-08-13
中国糖料(2021年3期)2021-07-13
种子(2020年11期)2020-12-05
中国临床医学影像杂志(2019年1期)2019-04-25
天然产物研究与开发(2019年1期)2019-03-01
中国医药指南(2017年3期)2017-11-13
安徽医科大学学报(2015年9期)2015-12-16
医学研究杂志(2015年9期)2015-07-01
