
干旱胁迫对甘肃地区5种经济林苗木水分利用效率的影响
2018-05-17 04:15种培芳单立山苏世平
干旱地区农业研究 2018年2期
种培芳,单立山,苏世平,李 毅
(甘肃农业大学林学院, 甘肃 兰州 730070)
甘肃位于我国西北部,地域辽阔,自然气候差异大。地形和生态环境的复杂性造就了甘肃丰富的林果树种,苹果、枸杞、大结杏、核桃、花椒、枣等林果品质优良,驰名中外。但甘肃大部分也都为旱区,具有水资源短缺、降雨量少、蒸发量大的特点,尤其纬度的高低变化使得甘肃省各地水分差异性较大[1]。在这种大尺度水分变化条件下,同一物种如何在不同地域适应生存是物种进化的关键所在,而水分则是影响干旱地区经济林发展的主要因素。因此,选择和发展水分利用效率高的树种,对保护甘肃地区水土生态环境安全以及调整旱区农业结构、增加农民收入具有非常重要的意义。
水分是限制植物生长、发育和繁殖最重要的因素之一[2],随着水资源危机和干旱化危害的不断加剧,我国已面临着重大的农林生产问题[3]。水分利用效率(water use efficiency,WUE) 是植物生理活动过程中消耗水形成有机物质的基本效率,能够反映植物碳同化与水分耗散之间的关系,作为植物水分利用状况和抗旱性特征的一个客观评价指标,可为不同生境适宜树种的选择提供科学支持[4]。有研究表明,干旱胁迫下水分利用效率高的植物其生产力也会相对较高[5]。因此,在水分匮乏条件下,选择高水分利用效率的植物种具有重要意义。目前,有些学者采用光合速率与蒸腾速率之比即瞬时水分利用效率(instant water use efficiency, WUEI)来表征,但这个值仅代表某特定时间内植物部分叶片的行为,适于研究引起水分利用效率快速变化的生理生态过程,并不能说明植物本身的水分利用特点[6-7]。所以,更多的研究者利用现代生态学研究的一种新方法即稳定碳同位素比率(δ13C)来代表植物长期生活过程的平均水分利用效率[8-9]。现在,这一方法的可靠性与稳定性已在许多研究结果中得以证实,例如,张怡等对干旱胁迫下四倍体刺槐幼苗水分利用效率及稳定碳同位素比率的研究发现δ13C可以作为筛选高WUE刺槐品种的指标[10];余新晓等利用δ13C值研究北京山区典型树种水分利用效率时发现叶片δ13C值和水分利用效率同向变化[11]。
甘肃作为国家“一带一路”建设战略的重要地区之一在培育优势林果业方面具有重要的地位,特别是在水资源短缺的条件下发展何种经济林树种是值得思考的一个问题。本研究以甘肃地区主要的5种经济林苗木为试材,探讨了这5种苗木在不同水分条件下的水分利用效率特征及其与稳定性碳同位素间的关系,以期为甘肃地区适宜经济林树种的选择及甘肃地区林业可持续发展提供理论依据和方法。
1 研究区概况
研究区位于甘肃省武威市林业高新技术示范园区的良种繁育基地,地处甘肃省河西走廊东北部。该区属典型的温带大陆性荒漠气候,土壤为碱性沙土。具体自然概况见表1。

表1 试验地自然概况
2 材料与方法
2.1 试验材料
试验于2015年4月—2015年10月进行, 试材选用本示范园提供的生长状况相近的核桃(Juglansregia)、枣(Ziziphusjujuba)、枸杞(Lyciumchinense)、沙棘(Hippophaerhamnoides)和花椒(Zanthoxylumbungeanum)2年生实生苗。
2.2 试验设计
以上述5个树种为试材,每个树种选择生长健康、大小基本一致的苗木,在2015年4月上旬移植到高35 cm,盆口直径为37 cm的花盆中,盆土为90%苗圃土配10%蛭石,每盆1株。每个树种各48盆,充分浇水,确保苗木生长良好。所有实验材料均放在微生境一致的条件下以排除其它因素对水分利用效率的影响。待缓苗为2个月时(约6月初)进行干旱处理,设定4个水分处理,分别为对照(土壤相对含水量为75%~90%)、轻度干旱(土壤相对含水量为60%~70%)、中度干旱(土壤相对含水量为40%~55%)和重度干旱(土壤相对含水量为25%~35%),每个处理12盆;于每天下午六点用铝盒取回土样后置于105℃烘箱中烘干至恒重计算含水量并根据缺水量补充水分使各处理均达到相应的干旱程度。采用遮雨棚防止自然降雨对干旱处理的影响。于8月下旬开始取样,每次随机取3株苗木。
2.3 指标测定
2.3.1 长期水分利用效率(WUEL) 生物量测定分别在6月初和8月底于控水试验开始前和结束后进行,取生长一致的5个树种苗木各3株,洗净后放入烘箱杀青并烘至恒重。用万分之一的电子感应天平称干物质量,用减重法测得控水期间生物量的累计值(单位:g)。水分处理期间的生物量增加值与总浇水量(扣除参照花盆盆土的蒸发量,即以植株的耗水量为基数)的比值即为控水处理期间的长期水分利用效率WUEL[8],单位为mg·mL-1。
2.3.2 瞬时水分利用效率(WUEI) 于控水处理后期(8月上旬),选晴好天气3天,用手提式光合测定系统(Lcpro+Ultra Compact Photosynthesis System,UK)在当地时间9∶00—11∶00分别对标记的3株生长势一致和位置相近的叶片进行净光合速率(Pn,CO2μmol·m-2·s-1)测定,同时获得蒸腾速率(Tr,H2O mmol·m-2·s-1)、气孔导度(Gs,H2O mol·m-2·s-1)。每次测定3个重复。瞬时水分利用效率根据WUE=Pn/Tr[12]计算。
2.3.3 稳定性碳同位素比率(δ13C) 将用于测定生物量的每个树种每个处理的3株叶片80℃烘干至恒重,研磨过100目筛,称6~7 mg装入安踣玻璃管,在铂金丝的催化下与氧化铜在550℃~600℃下放出的氧气燃烧,然后把燃烧生产的气体经真空纯化系统纯化得到纯净的CO2,用MAT-252质谱仪(Finningan MAT Delta S., Bremen,Germany)测定稳定性碳同位素比率,以PDB(Pee Dee Belemnite)为标准,根据下面的公式进行计算:
δ13C={[(13C/12C)sample-
(13C/12C)PDB]/(13C/12C)PDB}×1000
其中,δ13C为样品稳定碳同位素比率(‰),因其绝对值非常小,故以‰表示。δ13C皆为负值,表明样品中13C小于PDB标准中13C。
2.4 统计分析
试验中原始数据的整理采用Microsoft Excel 2003软件完成;用SPSS 19.0软件进行数据分析,用LSD多重比较进行显著性检验。
3 结果与分析
3.1 干旱胁迫对5个树种光合速率、蒸腾速率及气孔导度的影响
5个树种的光合速率(Pn)随干旱胁迫程度的增加而呈逐渐下降趋势(表2),除枣在轻度胁迫下的Pn与CK间无差异外,其它4个树种在3种干旱胁迫处理下(轻度、中度和重度)的Pn均与CK间差异显著(P<0.05)。各树种均在重度干旱胁迫下达到最低值,分别比对照下降了64.73%(花椒),62.04%(核桃),81.01%(枣),49.97%(枸杞)和48.48%(沙棘)。

表2 不同干旱处理下5个树种的光合速率
注:表中数据为平均值±标准差(n=9),不同小写字母表示处理间和树种间在0.05水平差异显著,下同。
Note: Data in table is the average±SD (n=9), different upper cases represent the significant difference between treatments and between species and clones at 0.05 level, respectively; the same as following.
与光合速率相比,干旱胁迫下5个树种的蒸腾速率(Tr)变化较为复杂(表3)。其中花椒、核桃、枸杞和沙棘均逐渐下降,枣则先升高后下降。5个树种的Tr均在重度干旱胁迫下达到最低,分别比对照降低了84.32%(花椒),67.87%(核桃),58.74%(枣),49.71%(枸杞)、52.21%(沙棘),与CK间差异显著(P<0.05)。
5个经济林树种在不同水分条件下的气孔导度(Gs)除了枸杞的呈先下降后升高再下降外,其它4个树种的Gs均随着水分胁迫的增加而逐渐下降(表4)。在重度胁迫时5个树种的Gs均达最低值,分别比CK下降了87.8%(花椒)、73.82%(核桃)、83.54%(枣)、10.28%(枸杞)、83.72%(沙棘)。除枸杞外其它4个树种的各水分处理间差异显著(P<0.05),而枸杞仅中度胁迫与其它处理间差异显著(P<0.05)。
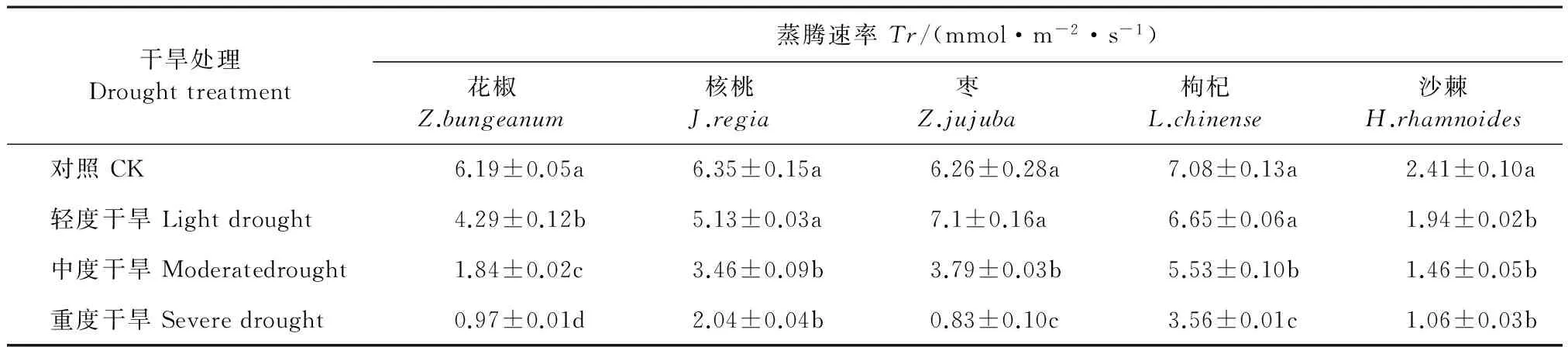
表3 不同干旱处理下5个树种的蒸腾速率
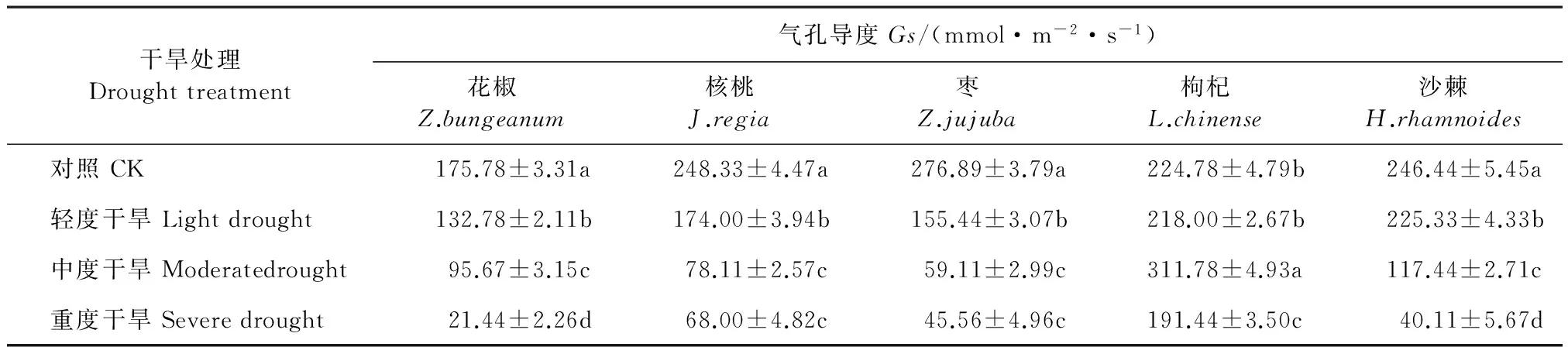
表4 不同干旱处理下5个树种的气孔导度
3.2 干旱胁迫对5个树种瞬时水分利用效率的影响
5个经济林树种的瞬时水分利用效率(WUEI)在不同干旱胁迫下的变化不同(表5)。花椒为升—降—升,在重度干旱胁迫时达到最大值,比CK增加了126.43%。核桃为先升后降,在中度胁迫时达到最大值,比CK增加了23.21%。枣的WUEI在轻度干旱胁迫时开始下降,随着胁迫程度的增加,其值又开始增加,在重度胁迫时达到最大值,比CK增加了43.68%。枸杞的WUEI则随着干旱胁迫程度的增加而先降低后升高的变化,在重度干旱胁迫时比CK增加了4.44%。沙棘的WUEI呈降—升—降的趋势,在中度干旱胁迫时达到最大值,比CK增加了16.18%。核桃各处理间差异不显著,沙棘仅轻度胁迫与其它处理间差异显著(P<0.05),其它3个树种则是重度胁迫与对CK间差异显著(P<0.05)。
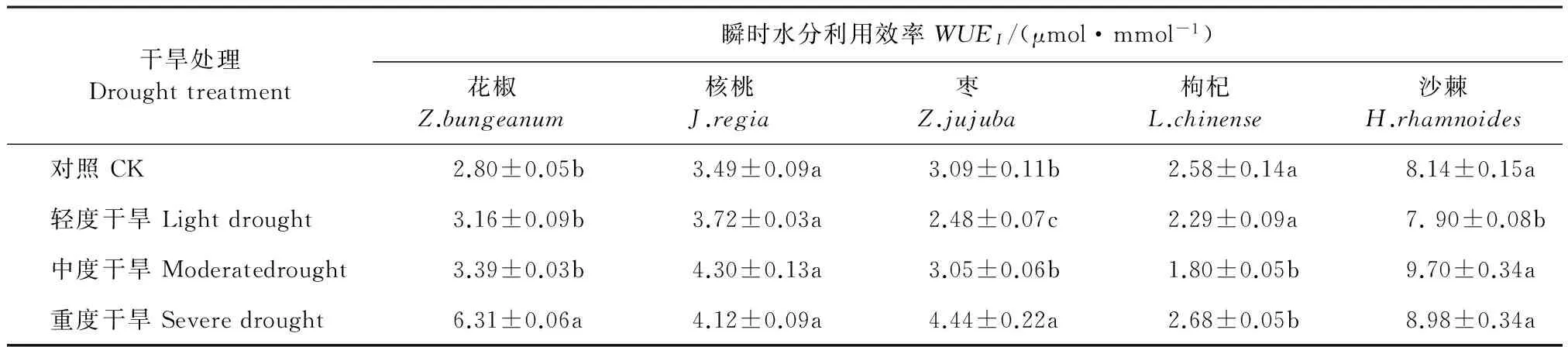
表5 不同干旱处理下5个树种的瞬时水分利用效率
3.3 干旱胁迫对5个树种生物量的影响
随干旱胁迫程度的加剧,花椒、核桃和枣3个树种的生物量呈现逐渐下降的趋势(表6),而枸杞和沙棘的生物量则在轻度胁迫时较CK有所上升,之后又呈下降趋势。5个树种均在重度干旱胁迫下达到最低值,分别比CK下降了73.73%(花椒)、54.52%(核桃)、70.49%(枣)、12.91%(枸杞)、11.81%(沙棘),且花椒、核桃和枣在重度干旱胁迫时与CK间差异显著(P<0.05),而枸杞和沙棘则差异不显著。
3.4 干旱胁迫对5个树种长期水分利用效率的影响
花椒和枣的WUEL随干旱胁迫加剧表现为先升高后降低,核桃是先降低后升高,枸杞和沙棘则是逐渐上升(表7)。各树种在达到了最大值时分别比CK增加了11.31%(花椒)、12.56%(核桃)、21.15%(枣)、43.06%(枸杞)、41.26%(沙棘),且与CK间差异显著(P<0.05)。
3.5 干旱胁迫对5个树种稳定碳同位素的影响
5个树种的δ13C整体呈随干旱胁迫程度的加剧而增加的趋势(表8),均在重度干旱胁迫时达到最大值。与CK相比,花椒增加了8.14‰,核桃增加了11.59‰,枣增加了7.40‰,枸杞增加了10.29‰,沙棘增加了8.83‰。此外,5个树种间的δ13C也存在差异,尤其是枸杞叶片的δ13C值在对照和重度干旱胁迫均大于其它4个树种,且可以看出干旱胁迫所引起的δ13C差异要比树种间的差异大,可以说水分是引起δ13C的差异的主要因素。
3.6 δ13C与WUEL的关系
5个树种δ13C与WUEL的关系均呈显著正相关性(图1),相关系数分别可达到0.504(花椒)、0.666(核桃)、0.746(枣)、0.839(枸杞)和0.899(沙棘)。而不同水分处理下两者的相关性分别为0.592(CK)、0.597(轻度胁迫)、0.901(中度胁迫)、0.938(重度胁迫)。这表明随着水分胁迫程度的增加,δ13C与WUEL的相关关系也越来越紧密。
4 讨 论
干旱胁迫会导致植物发生一系列生理生态反应,从而适应干旱环境。这种反应包括生物量的调配和水分利用效率的提高等[13]。本研究发现,重度干旱胁迫下5个树种的WUEI均比CK高。这与陈超等[14]研究指出,在干旱条件下植物的水分利用效率会得到提高,水分利用方式会更加有效的报道一致,只是干旱胁迫的程度不同而已。WUE与植物生理因子如叶水势、气孔、光合速率、蒸腾速率等有关[15],其中气孔由于对CO2和水汽扩散的影响不同步,进而影响植物WUE。在本研究中,干旱胁迫导致5个树种气孔导度下降,而水分利用升高,这与Farquhar[16]和龚吉蕊等[17]的研究结果相似。分析5个树种出现这种现象的主要原因是,气孔导度下降,引起光合速率和蒸腾速率的下降,但蒸腾速率下降比光合速率下降快,最终使得水分利用效率升高。

表6 不同干旱处理下5个树种的生物量

表7 不同干旱处理下5个树种的长期水分利用效率

表8 不同干旱处理下5个树种的稳定碳同位素比率比较

图1 5个经济林树种稳定碳同位素组成(δ13C)和WUEL的相关性分析
Fig.1 Analysis of the relationship between δ13C andWUELin five economic forest species
长期水分利用效率能代表一个植物个体的水分状况[18]。本研究结果表明,5种经济林树种在正常供水条件下和重度干旱胁迫WUEL值大小的排序不同,说明5个树种除了在遗传因素上的WUEL差异外,水分条件的改变也会使其WUEL不同。植物降低生长可以提高WUE,适应干旱环境[19]。本研究发现,干旱胁迫导致5个树种的生物量下降,水分利用效率提高,与前人研究结果相似。但纵观5个树种在重度胁迫下WUEL的增幅和生物量的降幅(表6和表7)可以看出,枸杞和沙棘的WUEL的增幅显著大于其它3个树种,但生物量的降幅却小于它们。说明干旱胁迫不但提高了沙棘和枸杞的水分利用效率,也使得它们能利用这些水分来减缓干旱胁迫引起的生物量下降,从而提高对其自身干旱的适应性,这可能是它们响应干旱的一种生理机制。
C3植物13C的自然丰度提供了一个有效测定植物长期碳水平衡的措施[20]。近年来,国内外许多学者对不同树种及其不同基因型的δ13C和WUE的关系进行了研究[21-24],结果表明,水分因子的改变造成植物的WUE差异都能在δ13C值上反映出来,δ13C和WUEL有很强的相关性。尤其是在干旱胁迫下,高δ13C可作为植物高WUE无性系选择的可靠指标[25-26]。但也有学者认为,在正常水分条件下,δ13C与WUEL呈负相关,而在干旱条件下,δ13C则与WUEL没有相关性[27]。本研究结果发现,5个树种之间的δ13C和WUEL之间的相关性各不相同。就树种而言,沙棘δ13C与WUEL间的相关性最大,其它依次为枸杞>核桃>枣>花椒。就不同水分处理间比较,则随水分的减少,δ13C与WUEL间的相关性越来越显著,即重度胁迫>中度胁迫>轻度胁迫>CK。这说明,干旱胁迫下高δ13C是评定这5个树种高WUE的可靠指标。此结论与张怡等在刺槐上的研究结果相似[10],而与赵凤君等在黑杨上的研究结果不同[26]。可能是因为干旱处理的方法不同,也可能是物种间水分利用效率机制不同。
Brooks等[28]认为,落叶乔木δ13C大于落叶灌木的δ13C。本研究的5个树种中,核桃、花椒和枣是属于落叶乔木或小乔木,但它们的δ13C小于灌木枸杞和沙棘δ13C。这和以往的研究的结果不同,可能是分析方法或植物的生态适应性不同,或者是因为所选择的核桃、花椒和枣较小的苗木,所以其本身的δ13C较小的缘故。当然,δ13C还和其它很多因素有关,究其和水分利用效率的关系,也会和其它因素如物种、水分条件、气候环境等有关,需要进行进一步的研究。
综上所述,干旱胁迫下枸杞和沙棘因其高水分利用效率和高δ13C值而比其它几个树种拥有更高的抗旱性。所以在甘肃省水分条件较差的地区可以选择这2个经济林树种,以发挥其水分利用高的特点来适应当地生境,而在水分较好的地区则可以发展其它3个树种。
参考文献:
[1] 闫业庆,胡雅杰.甘肃省降水量时空分布特征浅析[J].甘肃科技,2011,27(1):63-66.
[2] 肖春旺,周广胜.毛乌素沙地中间锦鸡儿幼苗生长、气体交换和叶绿素荧光对模拟降水量变化的响应[J].应用生态学报,2001,21(5):692-696.
[3] 张光灿,刘 霞,贺康宁,等.金矮生苹果叶片气体交换参数对土壤水分的响应[J].植物生态学报,2004,28(1):66-72.
[4] 严昌荣,韩兴国,陈灵芝,等.暖温带落叶阔叶林主要植物叶片中δ13C值的种间差异及时空变化[J].植物学报,1998,40(9):553-859.
[5] Lucero D W, Grieu P, Guckert A. Water deficit and plant competition effects on growth and water use efficiency of white clover(TrifoliumrepensL.) and ryegrass (LoliumperenneL.)[J]. Plant and Soil, 2000,227:1-15.
[6] 刘 冰,赵文智.荒漠绿洲过渡带柽柳和泡泡刺光合作用及水分代谢的生态适应性[J].中国沙漠,2009,29(1):101-107.
[7] 闫海龙,张希明,许 浩,等.塔里木沙漠公路防护林植物沙拐枣气体交换特性对干旱胁迫的响应[J].中国沙漠,2007,27(3):460-465.
[8] 赵凤君,沈应柏,高荣孚,等.叶片δ13C与长期水分利用效率的关系[J].北京林业大学学报,2006,28(6):40-45.
[9] 宋锋惠,吴正保,俞 涛,等.新疆5个枣品种叶片碳同位素组成、瞬时水分利用效率的季节变化及与气象因子的关系[J].果树学报,2012,29(1):66-70.
[10] 张 怡,罗晓芳,沈应柏.干旱胁迫下四倍体刺槐幼苗水分利用效率及稳定碳同位素组成的研究[J].西北植物学报,2009,29(7):1460-1464.
[11] 余新晓,杨芝歌,白艳婧,等.基于δ13C值的北京山区典型树种水分利用效率研究[J].应用基础与工程科学学报,2013,21(4):593-599.
[12] 李荣生,许煌灿,尹光天,等.植物水分利用效率的研究进展[J].林业科学研究,2003,16(3):366-371.
[13] 陈 平,孟 平,张劲松,等.两种药用植物生长和水分利用效率对干旱胁迫的响应[J].应用生态学报,2014,25(5):1300-1306.
[14] 陈 超,赵丽丽,王普昶,等.百脉根对干旱胁迫的生长、生理生态响应及其抗旱性评价[J].水土保持学报,2014,28(3):300-306.
[15] 蔡丽平,吴鹏飞,侯晓龙,等.干旱胁迫对水土保持先锋植物类芦光合特性的影响[J].水土保持学报,2011,25(6):237-214,259.
[16] Farquhar G,张玉翠译.水分利用效率与用水有效性:基于气孔视角的稳定同位素应用研究[J].中国生态农业学报,2014,22(8):886-889.
[17] 龚吉蕊,黄永梅,葛之葳,等.4种杂交杨对土壤水分变化的生态学响应[J].植物生态学报,2009,33(2):387-396.
[18] 陈 平,张劲松,孟 平,等.稳定碳同位素测定水分利用效率的可行性分析—以决明子为例[J].生态学报,2014,34(19):5453-5459.
[19] Saraswathi S G, Paliwal K. Drought induced changes in growth, leaf gas exchange and biomass production inAlbizialebbeckandCassiasiameaseedlings[J]. Journal of Environmental Biology, 2011,32:173-178.
[20] 刘海燕,李吉跃.稳定碳同位素在植物水分利用效率研究中的应用[J].西北林学院学报,2008,23(1):54-58.
[21] 苏 波,韩兴国,李凌浩,等.中国东北样带草原区植物δ13C值及水分利用效率对环境梯度的响应[J].植物生态学报,2000,24(6):648-655.
[22] 孙守家,孟 平,张劲松,等.华北石质山区核桃-绿豆复合系统氘同位素变化及其水分利用[J].生态学报,2010,30(14):3718-3726.
[23] 方晓娟,李吉跃,聂立水,等.毛白杨杂种无性系稳定碳同位素值的特征及其水分利用效率[J].生态环境学报,2009,18(6):2267-2271.
[24] 严昌荣,韩兴国,陈灵芝.六种木本植物水分利用效率和其小生境关系研究[J].生态学报,2001,21(11):1952-1956.
[25] Osorio J, Pereira J S. Genotypic differences in water use efficiency and 13C discrimination in Eucalyptus globules[J]. Tree Physiology, 1994,(4):871-882.
[26] 赵凤君,高荣孚,沈应柏,等.水分胁迫下美洲黑杨不同无性系间叶片δ13C和水分利用效率的研究[J].林业科学,2005,41(1):36-41.
[27] Ehleringer J R, Hall A E, Farquha R G D. Stable Isotopes and Plant Carbon-water Relations[M]. New York: Academic Press, 1993:247-267.
[28] Brooks J R, Flanagan B, Buchmann N, et al. Carbon isotope composition of boreal plants: functional grouping of life forms[J]. Oecologia, 1997,110:301-311.
猜你喜欢
今日农业(2022年13期)2022-09-15
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
中国水土保持(2022年3期)2022-03-24
河南水利与南水北调(2022年1期)2022-03-16
山西林业(2021年2期)2021-07-21
今日农业(2020年15期)2020-12-15
今日农业(2020年17期)2020-10-27
今日农业(2019年14期)2019-01-04
现代农村科技(2018年6期)2018-01-18
