
晋西黄土区苹果花生间作系统小气候效应
2018-05-17 04:15蔡智才毕华兴许华森常译方刘京健
干旱地区农业研究 2018年2期
蔡智才,毕华兴,2,3,4,5,6,许华森,王 丹,常译方,刘京健
(1.北京林业大学水土保持学院, 北京 100083; 2.水土保持国家林业局重点实验室(北京林业大学), 北京 100083; 3.北京市水土保持工程技术研究中心(北京林业大学), 北京 100083; 4.林业生态工程教育部工程研究中心(北京林业大学), 北京 100083; 5.山西吉县森林生态系统国家野外科学观测研究站, 北京 100083; 6.北京林果业生态环境功能提升协同创新中心, 北京 102206; 7.中国农业大学资源与环境学院, 北京 100094; 8.吉县林业工作站, 山西 临汾 042200)
农林复合系统能够有效地协调农林用地矛盾、提高自然资源利用效率和增加当地农民收入等。在人口爆炸和农林用地矛盾等问题不断严重的背景下,农林复合系统受到了许多国家和地区的重视,不断发展[1-5]。农林复合系统改变了原来森林系统或农田系统内的辐射、热量和水汽交换等物理基础,形成了具有自身特点的小气候效应[6]。Silva等[7]对西班牙大西洋海岸的林草间作系统研究发现,与单作系统相比,间作系统内光合有效辐射、大气温度和季节性的牧草产量波动均变得更小;Tamang等[8]研究美国佛罗里达防风林+作物间作系统提出,间作系统可以改善小气候(降低风速和增加空气相对湿度等),从而提高当地农民的作物产量;Peng等[9]通过3种林木+大豆(Glycine max)间作系统和大豆单作系统对比研究指出,相较于单作系统,间作系统的PAR降低了39.9%~57.9%,大气温度减少了1.0℃~1.4℃,空气相对湿度增加了5.9%~8.0%,大豆产量减产了58.8%~71.8%。小气候是植物生长和发育最重要的环境因子之一,近年来国内外对间作系统小气候的研究也越来越多[10-14]。然而,这些研究大多均未消除间作系统内林木与作物地下部分的相互影响,以致无法突显间作系统的小气候效应。同时,现有研究基本上只将间作系统的小气候与单作系统进行比较,而很少对间作系统内距树行不同距离处的小气候状况进行分析比较。另外,光合作用作为植物生长的基础,对环境因子变化十分敏感,其反应主要表现为净光合速率大小随环境条件不同而发生变化,因此将果农间作系统内作物净光合速率与间作系统小气候因子结合研究至关重要,而目前关于该方面的研究报道还显得非常不足。
本文以晋西黄土区具有代表性的苹果+花生间作系统作为试验对象,以花生单作系统作为对照,并在果树与作物间设置根障,以阻断两者根系的交叉延生和土壤水分、养分的运移;将间作系统内每一作物行当做一个整体,分析比较不同作物行的小气候因子和作物生长情况,评价间作系统各小气候因子对花生生长的影响程度,以期为优化果农间作系统的结构,提高整体效益和可持续经营提供理论依据。
1 材料与方法
1.1 研究区概况
研究区地理位置在山西省吉县,坐标为北纬35°53′10″~36°21′02″,东经36°21′02″~36°21′02″,是典型的黄土残塬沟壑区,属暖温带大陆性气候。据当地气象站资料可知,研究区多年平均降水量571 mm,主要集中在6—9月,季节性变化明显,年平均太阳总辐射5 424 MJ·m-2,日照时数2 563.8 h,多年平均气温9.9℃,日较差11.5℃,大于10℃的积温3 358℃,无霜期172 d。土壤类型为黄土母质,土层深厚且质地均一,较适宜果树与作物生长。果农间作系统中的果树树种以苹果为主,截止至2013年,吉县的苹果树栽植面积已占该地区耕地总面积的84.8%;主要农作物为花生、玉米(Zea mays)和大豆等。
1.2 材料
在研究区内选择苹果+花生间作系统与花生单作系统(CK)作为试验对象。苹果树品种为矮化富士,树龄7年,平均株高3.55 m,冠幅半径1.97 m,地径8.16 m,株行距为4 m×5 m,树行东西走向;花生品种为海花1号,株行距0.3 m×0.4 m,行向和树行方向一致,播种于2015年5月上旬。
1.3 方法
1.3.1 试验样区设计 在果农间作系统内,选择6棵果树作为一个试验样区,设置3个重复。种植作物前,在距离苹果树行1.0 m处开挖一条深1.0 m,宽0.2 m的壕沟,并于壕沟内靠树侧布设石棉瓦,以此阻断土壤水分和养分在果树与作物之间的运移以及果树树根和作物根系的交叉,形成根障,作物在距树行1.5 m处开始播种,作物行由北至南依次编号为R1~R6,试验样区的具体情况如图1所示。果农间作系统和作物单作系统的各项农艺管理措施保持一致。
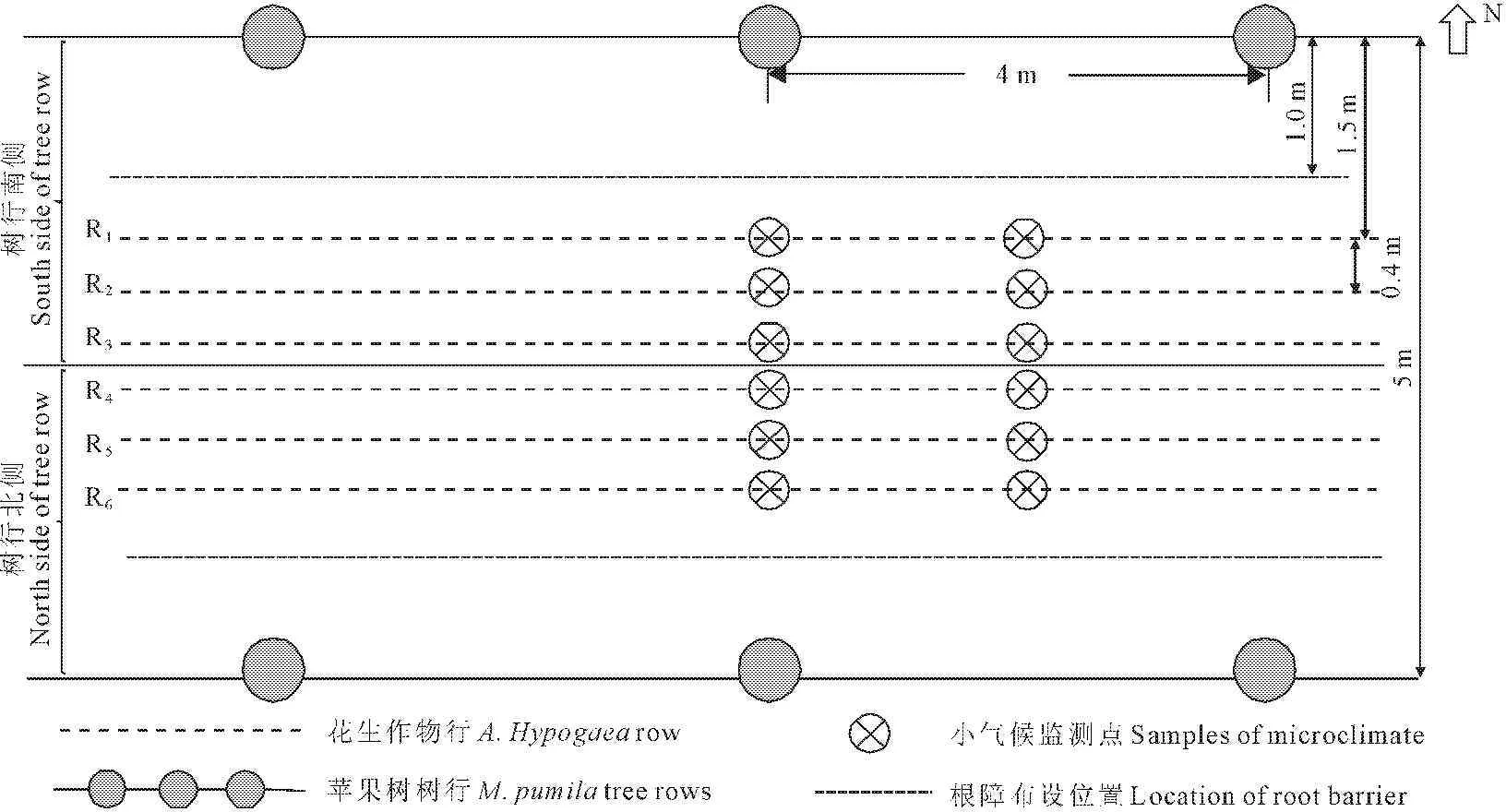
图1样区设计示意图
Fig.1 Sketch of plot design
1.3.2 小气候因子监测 在花生的开花期(7月)和结荚期(8月)选择晴朗无云的天气进行各项数据测定。本试验测定的小气候因子包括光合有效辐射(PAR)、大气温度、空气相对湿度和风速,其中PAR采用棒状光合辐射传感器LI-191SA(美国,LI-COR公司生产)测定,大气温度、空气相对湿度和风速则采用HOBO-U30便携式小型自动气象站(美国,ONSET公司生产)进行测定。将传感器布设在间作系统内的每一作物行,具体情况见图1。测定时间为09∶00—18∶00,测定间隔为1 h·次-1,每个物候期重复3 d取平均值。在作物单作样地内随机选择3个样点,对上述小气候指标采取相同方法进行监测。定义相对某小气候因子:间作系统内某作物行的该小气候因子日均值/单作系统的该因子日均值×100%。
1.3.3 作物净光合速率测定 在测定小气候因子的同时,利用LI-6400XT便携式光合测定系统(美国,LI-COR公司)对花生的净光合速率(Pn)进行测定。从09∶00—18∶00每隔1 h,选取间作系统内每一作物行,长势相近、伸展充分且无病虫害的健康叶片3片进行测定,取平均值。单作系统内按“S”形选取5片叶片进行相同测定。
1.3.4 作物产量测定 在2015年生长季末,将试验样区内花生以作物行为单位收取,采集的花生在70℃下烘至恒重,采用精度0.01 g的电子天平称量,取3个重复试验小区的平均值。花生单作系统内,作物产量的取样和测定方法与间作系统相同。
2 结果与分析
2.1 苹果+花生间作系统不同作物行小气候日变化
2.1.1 苹果+花生间作系统不同作物行光合有效辐射日变化 苹果+花生间作系统R1受遮荫影响较大(图2),PAR在13∶00前小幅度上升,之后不断下降;R2的PAR强度在9∶00—11∶00上升明显,11∶00—13∶00之间小幅度波动,之后不断变小;R3和R4的PAR强度日变化趋势具有相似规律,表现为典型的“单峰型”,峰值出现在13∶00左右;R5和R6日变化趋势一致,只是R5的PAR强度略高于R6,两者PAR强度在12∶00前起伏不大,之后快速上升,14∶00达到峰值,之后逐渐下降。太阳辐射强度变化和果树树荫移动使间作系统内不同作物行的PAR日变化表现不同,总体表现为:R1、R2(位于树行南侧)主要在上午受果树遮荫影响;R3、R4(位于树行中间)受果树遮荫影响小;R5、R6(位于树行北侧)主要在下午受影响。整个间作系统的PAR相对指数为70.78%,不同作物行PAR相对指数有所不同,R1~R6的PAR相对指数分别为49.39%、67.69%、88.07%、86.77%、70.51%和62.27%,即距树行越近的作物行全天PAR强度越小(图3)。
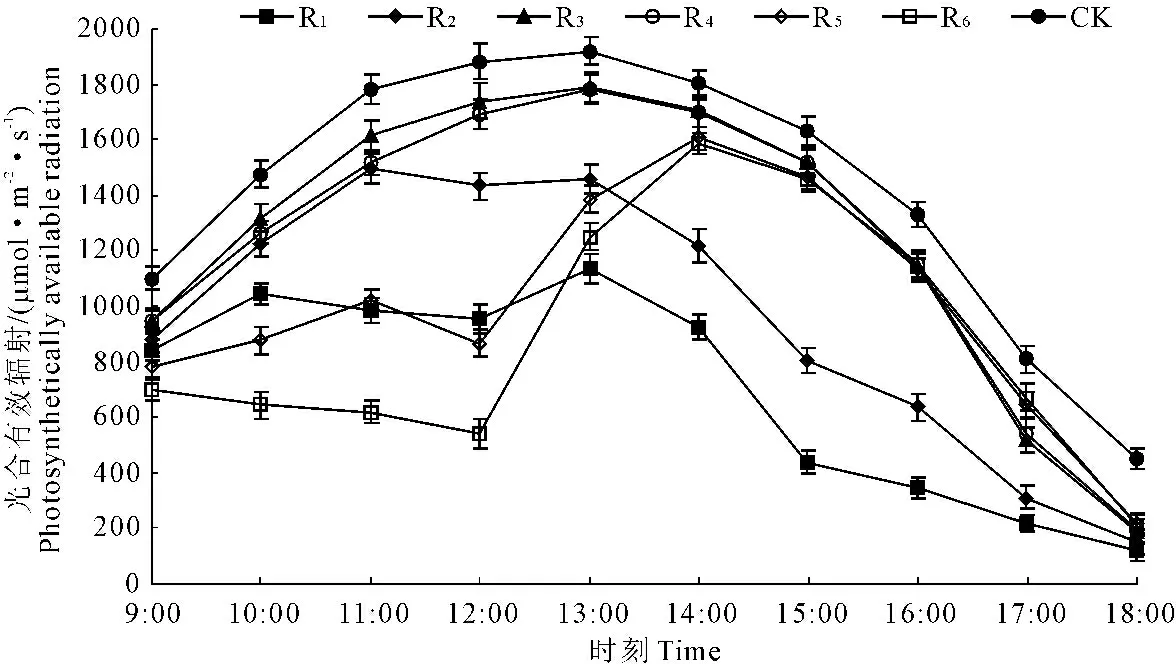

图2 苹果+花生间作系统内不同作物行光合有效辐射日变化
图3苹果+花生间作系统内不同作物行各指标相对指数雷达图
Fig.3 The radar chart of characteristic's relative index of different crop rows inM.pumilaandA.hypogaeaintercropping system
2.1.2 苹果+花生间作系统不同作物行大气温度日变化 苹果+花生间作系统所有作物行的气温日变化均呈“单峰型”(图4),峰值出现在14∶00左右;在09∶00—14∶00之间,随着气温的上升不同作物行之间气温的差距不断增大,14∶00—18∶00随着气温的下降不断减小。R1~R6的大气温度相对指数分别为93.98%、94.84%、97.32%、97.27%、93.93%和92.74%,大气温度日均值较CK分别下降了1.87℃、1.60℃、0.83℃、0.85℃、1.88℃、2.25℃,即越靠近树行气温下降越多(图3)。果农间作系统也在一定程度上减小了气温日较差,且距树行越近,气温日较差越小。
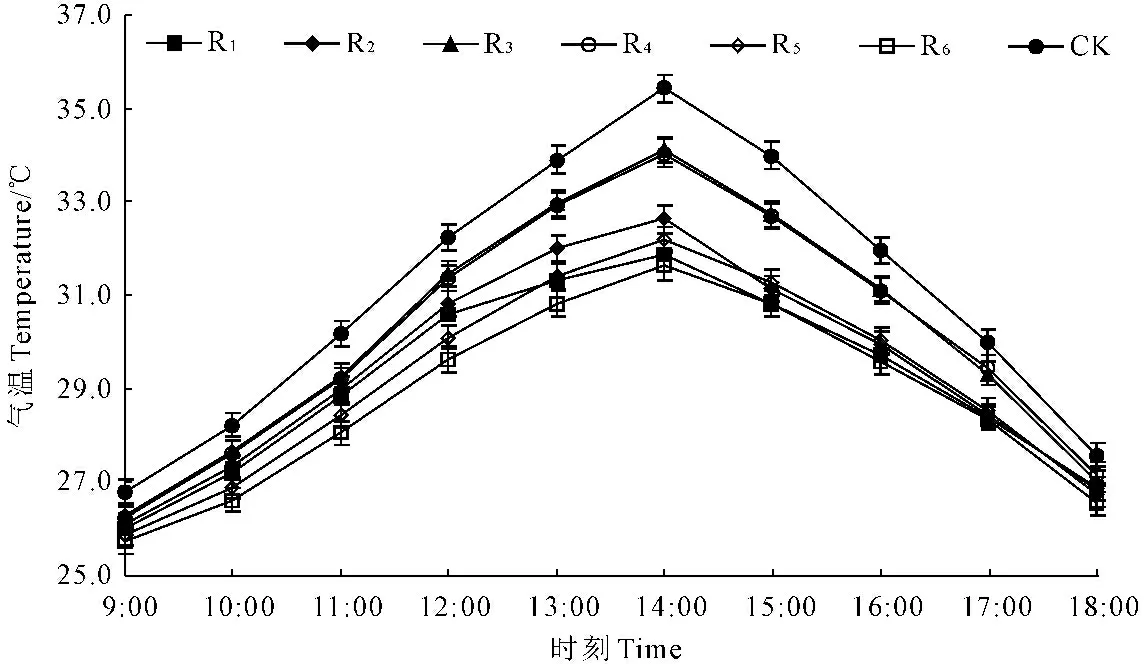
图4苹果+花生间作系统内不同作物行大气温度日变化
Fig.4 Diurnal variation of temperature of the different crop rows inM.pumilaandA.hypogaeaintercropping system
2.1.3 苹果+花生间作系统不同作物行空气相对湿度日变化 与大气温度日变化趋势相反,间作系统所有作物行的空气相对湿度日变化均呈“V”型,早晚空气相对湿度较高,中午低,最低值出现在14∶00左右(图5)。间作系统的所有作物行空气相对湿度均大于CK,间作系统整体空气相对湿度的相对指数为118.50%,R1~R6分别为123.72%、120.52%、112.37%、112.36%、119.37%和122.66%,距离树行南侧最近的R1的空气相对湿度显著大于距树最远的R3(P<0.01);距离树行北侧最近的R6也显著高于R4(P<0.01);不同作物行表现为距树行越近,空气相对湿度提升效果越明显(图3)。

图5苹果+花生间作系统内不同作物行空气相对湿度日变化
Fig.5 Diurnal variation of relativwe humidity of the different crop rows in
M.pumilaandA.hypogaeaintercropping system
2.1.4 苹果+花生间作系统不同作物行风速日变化 风速日变化并没有一定的规律,但是间作系统的风速在一天中不同时刻均不同程度的低于对照(图6)。间作系统整体的风速相对指数为71.88%,R1~R6的风速相对指数分别为71.02%、70.69%、74.22%、73.99%、70.58%和70.58%(图3)。方差分析结果表明,苹果+花生间作系统的风速显著低于花生单作系统(P<0.01)。间作系统内各作物行之间风速并无显著差异,仅处于间作系统中间区域的R3、R4略高于其他作物行。
2.2 苹果+花生间作系统不同作物行净光合速率日变化
间作系统各作物行的净光合速率日变化趋势存在差异(图7):CK、R3和R4日变化呈现“双峰型”,峰值分别为12∶00和15∶00左右,均出现了“午休”现象;R1、R2、R5、R6日变化趋势呈“单峰型”,但峰值出现时刻不同,R1、R2为13∶00左右,而R5、R6则是14∶00前后。R1、R2在13∶00达到最大值时,净光合速率比刚进入“午休”的R3、R4略高,之后不断下降;R5、R6在14∶00处于净光合速率峰值,高于其他所有作物行,之后也不断下降。间作系统整体的净光合速率相对指数为77.31%,R1~R6的净光合速率相对指数分别为58.99%、73.85%、90.97%、90.91%、77.57%和71.56%,表现为距树行距离越近的作物行,花生净光合速率越低(图3)。

图6苹果+花生间作系统内不同作物行风速日变化
Fig.6 Diurnal variation of wind speed of the different crop rows inM.pumilaandA.hypogaeaintercropping system
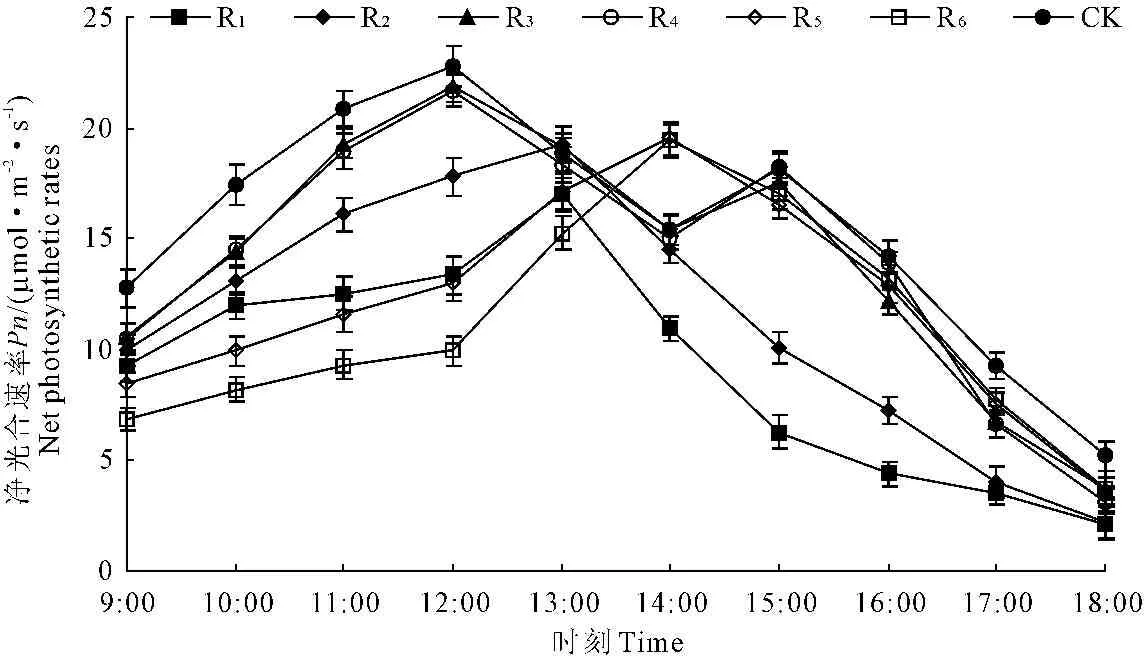
图7苹果+花生间作系统内不同作物行净光合速率日变化
Fig.7 Diurnal variation of net photosynthetic rates of the different crop rows in
M.pumilaandA.hypogaeaintercropping system
2.3 苹果+花生间作系统不同作物行生物学性状
间作系统花生的株高明显低于单作系统(表1),间作系统整体的平均株高较CK降低了25.61%,其中R1~R6比CK分别降低35.85%、26.62%、16.06%、14.96%、28.27%、31.92%;株高从大到小依次为R4>R3>R2>R5>R6>R1,即作物行距树行越近,株高越小。间作系统的产量也明显低于CK,与CK相比,间作系统整体的平均花生产量减产了39.44%,其中R1~R6的花生产量分别减产了55.08%、40.26%、24.34%、26.63%、38.76%、51.59%。
2.4 苹果+花生间作系统花生净光合速率、产量与小气候因子数量关系
2.4.1 苹果+花生间作系统花生净光合速率与小气候因子数量关系 间作系统花生净光合速率(Pn)与光合有效辐射(PAR)、大气温度(T)和空气相对湿度(RH)均在0.01水平上显著相关;而风速与Pn的相关系数仅为0.212,即两者并不具有线性相关性(表2)。
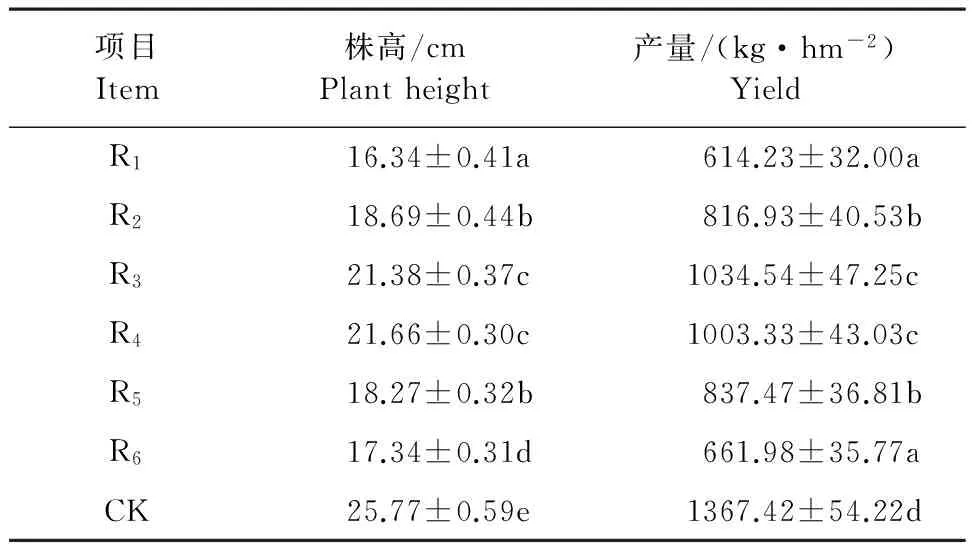
表1 苹果+花生间作系统不同作物行的生物学性状
注:同列不同字母表示差异显著(P<0.05)。
Note:the different letters of same column indicate significant difference(P<0.05).
为了进一步深入了解影响间作系统中花生净光合速率的主要小气候因子及其数量关系,以净光合速率为因变量,间作系统内光合有效辐射、大气温度和相对湿度为自变量,构建花生净光合速率与小气候因子的回归方程如下:
Pn=-26.364+0.00980PAR+0.692T+0.156RH(R2=0.936P<0.01)
(1)
进一步将方程标准化:
Pn=0.886PAR+0.292T+0.232RH
(2)
比较式(2)中各小气候因子前的系数可知,光合有效辐射对花生净光合速率的影响最大,大气温度次之,空气相对湿度影响最小。定义某一小气候因子对花生Pn的贡献率为该小气候因子前的系数占总系数的百分比,则PAR、大气温度和空气相对湿度对花生Pn的贡献率分别为62.84%、20.71%和16.45%。
2.4.2 苹果+花生间作系统花生产量与小气候因子数量关系 花生产量(Yield)与光合有效辐射(PAR)、空气相对湿度均在0.01水平上显著相关,与大气温度在0.05水平上显著相关,与风速不具有线性相关性(表3)。同样,以花生产量为因变量,间作系统内光合有效辐射、大气温度和相对湿度为自变量,建立以下花生产量与小气候各因子的回归方程:
Yield=-7463.795+1.129PAR+124.190T+62.389RH(R2=0.936P=0.19<0.05)
(3)
将方程标准化,得:
Yield=1.382PAR+0.425T+0.777RH
(4)
比较式(4)中各小气候因子的系数可知,在对花生产量的影响程度上,光合有效辐射最大,空气相对湿度次之,大气温度最小。同样,定义某小气候因子对花生产量的贡献率为该因子前的系数占所有因子系数和的百分比,则3者对花生产量的贡献率分别是53.48%、30.07%和16.45%。

表2 苹果+花生间作系统花生净光合速率与小气候各因子的相关系数
注:*表示在0.05水平上显著相关,**表示在0.01水平上显著相关。
Note:*indicates a significant correlation at the 0.05 level, **indicates a significant correlation at the 0.01 level.

表3 苹果+花生间作系统花生产量与小气候各因子的相关系数
注:*表示在0.05水平上显著相关,**表示在0.01水平上显著相关。
Note:* indicates a significant correlation at the 0.05 level, ** indicates a significant correlation at the 0.01 level.
3 讨 论
果树的出现打破了原来单一的农田种植结构,将原来作物单作的平面结构变为乔木与作物相结合的立体结构,塑造了新的小气候条件。本研究发现,果树在上、作物在下的双层结构造成果树对作物的存在一定程度遮荫作用,导致林下光合有效辐射减弱。果树树冠的遮荫及叶片的蒸腾作用,致使间作系统的大气温度降低,且气温日较差也变小了。果树的存在,导致间作系统内蒸发和蒸腾的水汽会在空气中停留较长时间,间作系统的空气相对湿度有一定程度提高。进入间作系统内的气流会受到果树的阻挡和摩擦作用,迫使气流分散,造成风速渐弱[15]。这与许华森等[16]研究结果相似,即间作果树一方面会对作物冠层的光合有效辐射产生负面效应;另一方面又对其他小气候因子产生正面效应,在一定程度上改善小气候。笔者还发现,距树行不同距离处的作物行小气候因子表现出差异性,表现为距树行越近小气候效应越明显,这是由于距树行越近,受到果树的影响程度越深。晁海等[17]对新疆轮台县杏棉间作系统的小气候分布特征进行研究,也得出类似结果。
作物的生长发育与周围的小气候状况密切相关,果农间作系统小气候条件的改变会影响间作作物的生长。在未超过作物光饱和点情况下,较高的PAR会增强作物的净光合速率(Pn);大气温度升高通常有利于作物进行光合作用,积累干物质,但是高温会在一定程度上损伤作物,抑制光合作用;在较干旱的黄土区,提高空气相对湿度有利于作物的生长;间作系统能在一定程度上防止风力过大引起的作物倒伏现象。本研究发现,花生单作系统和间作系统中间位置作物行均出现“午休”现象;而其他作物行由于果树影响,避免了“午休”,但是在一定程度上抑制了作物净光合速率;间作系统小气候对作物生长发育的综合效应表现为负效应,造成花生净光合速率和产量降低。这与彭晓邦等[18]对渭北黄土区核桃与作物间作系统的研究结果相似。统计分析表明,光合有效辐射对花生生长发育和产量的影响最大,PAR是作物进行光合作用和产量形成的能量来源,其质和量是决定作物Pn和产量的主要因子。
在认识到果农间作系统小气候效应对作物影响的基础上,如何采取适当的措施,使系统的整体效益最大化,一直是学者们研究的重点。孙尚伟等[19]提出对杨树玉米复合系统进行修枝处理,可提高林下PAR 39.7%~98.9%,同时提升玉米产量。因此,合理及时的修枝能在一定程度上降低间作系统小气候对作物的负效应,建议将苹果树冠层的修剪工作与花生物候期结合考虑,从而达到既不影响果树生长又增加作物产量的目的。包宛鑫[20]通过研究5种不同株行距的杏与粮棉间作系统发现,影响间作系统透光性最直接的因素是株行距,表现为株行距越大,透光性越好。本研究中,离树行越近的作物行所受影响越大,因此适当增加果树与作物的间作距离可有效减轻果树造成的负面效应。同时,为了更全面地揭示间作系统的小气候效应,还应结合大气CO2浓度、土壤表层温度等进行研究,得出更为准确的结论。
4 结 论
1) 苹果+花生间作系统小气候效应明显,与花生单作系统相比,间作系统的光合有效辐射明显减弱,大气温度降低,空气相对湿度提高,风速变小;且距树行越近,作物行的这种小气候效应越明显。
2) 与花生单作系统相比,苹果+花生间作系统内不同作物行的花生净光合速率、株高和产量均出现了不同程度的降低,且表现为距树行越近,花生所受影响越大。
3) 苹果+花生间作系统各小气候因子对花生生长和产量的影响程度不同,对花生净光合速率的贡献率为光合有效辐射最大,大气温度次之,空气相对湿度最小;对花生产量的贡献率为光合有效辐射最大,空气相对湿度次之,大气温度最小。
参考文献:
[1] Vrahnakis M, Nasiakou S, Kazoglou Y, et al. A conceptual business model for an agroforestry consulting company[J]. Agroforestry Systems, 2016,90(2):1-18.
[2] Betencourt E, Duputel M, Colomb B, et al. Intercropping promotes the ability of durum wheat and chickpea to increase rhizosphere phosphorus availability in a low P soil[J]. Soil Biology & Biochemistry, 2012,46(1):181-190.
[3] Mcdermott M E, Rodewald A D, Matthews S N. Managing tropical agroforestry for conservation of flocking migratory birds[J]. Agroforestry Systems, 2015,89(3):383-396.
[4] 解婷婷,苏培玺,周紫鹃,等.气候变化背景下农林复合系统碳汇功能研究进展[J].应用生态学报,2014,25(10):3039-3046.
[5] 李岩泉,何春霞.我国农林复合系统自然资源利用率研究进展[J].林业科学,2014,50(8):141-145.
[6] Sánchez i A, Lassaletta L, Mccollin D, et al. The effect of hedgerow loss on microclimate in the Mediterranean region: an investigation in Central Spain[J]. Agroforestry Systems, 2010,78(1):13-25.
[7] Silva-Pando F J, González-Hernández M P, Rozados-Lorenzo M J. Pasture production in a silvopastoral system in relation with microclimate variables in the atlantic coast of Spain[J]. Agroforestry Systems, 2002,56(3):203-211.
[8] Tamang B, Andreu M G, Rockwood D L. Microclimate patterns on the leeside of single-row tree windbreaks during different weather conditions in Florida farms: implications for improved crop production[J]. Agroforestry Systems, 2010,79(1):111-122.
[9] Peng X, Thevathasan N V, Gordon A M, et al. Photosynthetic response of soybean to microclimate in 26-Year-Old tree-based intercropping systems in southern ontario, canada[J]. Plos One, 2015,10(6):e0129467.
[10] H. Knørzer, H. Grøzinger, S. Graeff-Hønninger, et al. Integrating a simple shading algorithm into CERES-wheat and CERES-maize with particular regard to a changing microclimate within a relay-intercropping system[J]. Fuel & Energy Abstracts, 2011,121(2):274-285.
[11] Siles P, Harmand J M, Vaast P. Effects of Inga densiflora on the microclimate of coffee (Coffeaarabica L.) and overall biomass under optimal growing conditions in Costa Rica[J]. Agroforestry Systems, 2010,78(3):269-286.
[12] Menezes R S C, Salcedo I H, Elliott E T. Microclimate and nutrient dynamics in a silvopastoral system of semiarid northeastern Brazil[J]. Agroforestry Systems, 2002,56(1):27-38.
[13] 史彦江,卓热木·塔西,宋锋惠,等.枣农间作系统小气候水平分布特征研究[J].新疆农业科学,2010,47(5):888-892.
[14] 黄国勤,张明林,刘隆旺,等.秸秆覆盖对红壤旱地间作生态系统小气候的影响[J].中国生态农业学报,2006,14(1):76-78.
[15] 张俊娥,黄大庄,贾玉彬,等.条墩桑对间作田小气候影响的初步研究[J].蚕业科学,2007,33(1):102-105.
[16] 许华森,毕华兴,高路博,等.晋西黄土区苹果+大豆间作系统小气候及其对作物生产力的影响[J].中国水土保持科学,2014,2(2):9-15.
[17] 晁 海,张大海,徐 林,等.杏棉间作系统小气候水平分布特征研究[J].新疆农业大学学报,2007,30(1):35-39.
[18] 彭晓邦,仲崇高,沈 平,等.玉米大豆对农林复合系统小气候的光合响应[J].生态学报,2010,30(3):710-716.
[19] 孙尚伟,尹伟伦,夏新莉,等.修枝对复合农林系统内小气候及作物生长的影响[J].北京林业大学学报,2009,31(1):25-30.
[20] 包宛鑫.杏与粮棉间作系统内的光分布特征研究[D].乌鲁木齐:新疆农业大学,2011.
猜你喜欢
磷肥与复肥(2022年6期)2022-08-02
广西林业科学(2022年2期)2022-05-09
农业灾害研究(2022年1期)2022-05-07
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
南方农业·下旬(2016年9期)2017-04-27
江苏农业科学(2017年5期)2017-04-15
科技与创新(2016年18期)2016-11-04
湖北农业科学(2014年21期)2014-12-22
青年文摘·上半月(1988年2期)1988-11-01
