
核相关转录因子2对LOVO细胞裸鼠移植瘤的影响
2018-06-25 07:18史刚刚郝敬鹏
中国中西医结合外科杂志 2018年3期
韩 梅 ,王 晖,东 帅,史刚刚,郝敬鹏 ,何 彬
直肠癌是全世界第三常见的恶性肿瘤[1-2]。直肠癌细胞具有较强的放射抗拒性,且放射治疗还会引起消化道出血、疼痛和穿孔等副作用[3-4]。多项研究显示,人类Runt相关转录因子2(核相关转录因子2,Runx2)在调控骨骼形成和成骨细胞分化进程中发挥着极为重要的作用[5],在前列腺癌、结肠癌等多种癌变组织中均有异常表达[6-7],提示Runx2可能参与癌细胞的增殖活化和进展。研究显示,该蛋白对与癌细胞放射敏感性相关的信号通路具有调控作用[8]。
1 材料与方法
1.1 材料及来源 冻存直肠癌LOVO细胞购买自中国医学科学院。31例直肠癌患者的癌组织及与之相配对的癌旁组织标本由天津医科大学总医院提供,组织样品均较为新鲜,未接受任何抗癌相关治疗。
胰蛋白酶,美国Sigma公司;PBS磷酸缓冲液,Hyclone;细胞培养箱,Heraeus Sepatech 公司。DMEM培养基、胎牛血清和PBS均购自Hyclone,Trizol试剂盒购自Invitrogen;BCA蛋白浓度测试试剂盒(增强型)、5×SDS蛋白上样缓冲液、20×TBS缓冲液等购买于南京建成生物。细胞处理等过程中使用的超净工作台提供自艺斯高上海贸易有限公司。人类Runt相关转录因子2(Runx2)一抗为兔抗人多克隆抗体,Runx2的二抗为辣根过氧化物酶标记的山羊抗兔多克隆抗体;内参GAPDH采用小鼠抗人单克隆抗体,其二抗为HRP标记的山羊抗小鼠多克隆抗体,所有抗体均由艾康生物技术(杭州)有限公司提供。Runx2 siRNA和pcDNA3.1-Runx2由上海生工生物有限公司合成提供。
45只体质量约20 g的雄性成年BALB/C裸鼠(7~8周龄)由天津医科大学动物实验中心提供,生产许可证号SCXK(津)2012-0001,动物使用许可证号SYXK(津)2014-0002。所有大鼠于室温25 ℃,湿度50%~80%,SPF环境下自由饮水和采食。
1.2 RT-PCR检测Runx2 mRNA的表达 癌组织及与之相配对的癌旁组织样品每个检测重复3次。通过Oligo 7.36 Demo软件设计引物,Runx2上游引物:5’-ACCAGCAGCACTCCATATCTCTAC-3’,下游引物:5’-CTTCCATCAGCGTCAACACC ATC-3’;内参GAPDH上游引物:5’-AACATCAT CCCTGCCTCTACTGG-3’,下游引物:5’-CCTCCG ACGCCTGCTTCAC-3’。提取细胞RNA并逆转录为cDNA,采用2-ΔΔCt分析法,以GAPDH基因作为内参,测定Runx2 mRNA相对表达水平。PCR反应条件:95 ℃预变性15 min,95 ℃变性30 s,55 ℃退火 30 s,72 ℃延伸 45 s,共 35个循环,72 ℃min。本组结果统计学分析,采用t检验。
1.3 基因转染 电穿孔法分别将Runx2 siRNA和重组表达载体pcDNA3.1-Runx2转染直肠癌LOVO细胞后,G418筛选得到稳定转染细胞系。将冻存直肠癌LOVO细胞置于60 ℃恒温水浴箱,30 s内进行快速复苏。融化后迅疾将细胞转入EP管并向其中加入DMEM培养基,重复小心吹打细胞使其均匀分布,随后离心弃上清液。将复苏完成的直肠癌LOVO细胞用DMEM完全培养基稀释调整为1×106/mL的密度。将直肠癌LOVO细胞随机平均分为3组,对照组(不实施任何基因转染)、Runx2上调组(将pcDNA3.1-Runx2进行基因转染)、Runx2下调组(将Runx2 siRNA进行基因转染)。
将处于生长旺盛状态的直肠癌LOVO细胞分别用胰蛋白酶消化并收集细胞,用HeBS液(140 mmol/L NaCl,5 mmol/L KCl,0.75mmol/L Na2HPO4,6 mmol/L Glucose,25 mmol/L Hepes)重悬、计数,调整细胞密度至5×106/mL。每次取200μL细胞悬液,分别加入3组电击杯中,向电击杯中分别再加入4μg生理盐水、pcDNA3.1-Runx2、Runx2 siRNA。电击后室温静置2 min。将3组直肠癌LOVO细胞分别加入到3份含有2 mL细胞培养液的六孔细胞培养板和96孔细胞培养板中,置于细胞培养箱培养。将3组细胞分别取出,去培养基并加入含有800 μg/mL G418的培养基,继续培养。定期换液并更换筛选培养基,将G418浓度减半继续筛选培养12~15 d。当看到有稳定的有抗性的克隆出现,及时停药,对其进行扩大培养。
1.4 48 h后检测细胞中Runx2 mRNA的表达 转染约48 h,采用荧光当量RT-PCR法,分别检测3组直肠癌LOVO细胞中Runx2基因相对表达水平。采用Western blotting方法测定其蛋白相对表达水平。每个检测重复3次。通过Oligo 7.36 Demo软件设计引物,Runx2上游引物:5’-ACCAGCAGCACTCCATATCTCTAC-3’,下游引物:5’-CTTCCATCAGCGTCAACACCATC-3’;内参GAPDH上游引物:5’-AACATCATCCCT GCCTCTACTGG-3’,下游引物:5’-CCTCCGA CGCCTGCTTCAC-3’。提取细胞RNA并逆转录为cDNA,凝胶图像分析系统进行光密度分析,以GAPDH基因作为内参,测定Runx2 mRNA相对表达水平。PCR反应条件:95 ℃预变性15 min,95 ℃变性 30 s,55 ℃退火 30 s,72 ℃延伸 45 s,共35个循环,72 ℃ 7 min。
1.5 48 h后检测细胞中Runx2蛋白的表达 转染48 h后取3组细胞,根据BCA试剂盒(北京益德益华生物有限公司)提取并测定细胞蛋白浓度[35]:将细胞裂解并稀释至相同的蛋白浓度,100 ℃煮沸5 min使蛋白变性。每组取蛋白40 μg,10% SDSPAGE凝胶电泳(70V,30 min;100 V,90 min);转膜(200 mA,3 h)至PVDF膜;5%脱脂牛奶室温封闭2 h (或4 ℃过夜);一抗1:500,室温孵育2 h (或4 ℃过夜);TPBS洗涤3次,PBS洗涤1次。之后进行蛋白免疫印迹的反应,拍照记录后进行软件的图像分析,测定3组细胞中Runx2蛋白质相对表达水平。
1.6 CCK-8检测细胞增殖 在直肠癌LOVO细胞经过基因转染后继续培养48 h,采用CCK-8法酶标仪下分别测定3组直肠癌细胞的增殖活化能力变化。转染48 h,将3组细胞分别收集接种到细胞培养96孔板上。细胞培养箱中培养至细胞贴壁后继续培养48 h,向各个孔中分别加入20 μL CCK-8试剂,振荡均匀。将3组癌细胞放入细胞培养箱中继续培养72 h,取出细胞,利用酶标仪测定OD490值。
1.1 乳腺癌组织来源 本研究中28例乳腺癌组织标本均来源于南昌市第三人民医院,入选标本为2016年1月至2017年6月间手术切除的乳腺癌组织。入选标准:①所有患者均行病理检查确诊为原发性乳腺癌[2],并经高年资医生根据AJCC乳腺癌分期系统确定患者临床分期[3];②所有患者无严重的肝、肾、心、脑等重要脏器损害;③研究符合伦理学标准,并通过了院伦理学委员会的批准,患者签署知情同意书;④年龄20~81岁,平均年龄(48.75±12.15)岁,体重42~60 kg,平均体重(44.00±3.86)kg。
1.7 克隆形成实验观察Runx2对LOVO的放射敏感性的影响 转染48 h之后将3组细胞消化、计数,调整细胞的浓度,将3组细胞分别接种到不同细胞数于6孔培养板中。培养24~30 h,对3组细胞分别给予0、2、4、6、8、10 Gy的单剂量照射(6MV X射线,30×30 cm2照射野,源皮距100 cm),静置培养14 d。用1%结晶紫固定液固定20 min。显微镜下计数≥50个细胞的克隆。计算不同剂量照射下的细胞存活分数:集落形成率(PE)=集落数/种植细胞数×100%,存活分数(SF)=实验组PE/对照组PE。GraphPad Prism 5软件进行剂量存活曲线拟合,计算相应的D0、N及SF2。Do为曲线指数区下降63%所需剂量, Do越大,放射抗拒性增强; 2Gy时存活分数SF2是代表细胞放射敏感性的重要指标。若将直线部分外推与纵坐标相交点的数值称为外推值N,代表细胞内靶的个数或所需击中靶的次数[9]。
1.8 TUNEL法检测凋亡 转染培养48 h,从细胞培养箱之中取出3组细胞分别测定各组中直肠癌LOVO细胞的凋亡情况。首先轻轻吹打细胞,使之均匀分布;洗涤细胞后滴加原位末端标记反应混合液50μL,在湿盒中37 ℃孵育1 h;滴加POD2转化液50μL,湿盒37 ℃孵育30 min,pH7.4的磷酸缓冲液(PBS)冲洗3次,二氨基联苯胺(DAB)显色。反应过后,用HE进行复染并在显微镜下分析图像,棕黄色细胞核的细胞为凋亡细胞,每张切片高倍光学显微镜下(×400)随机选取10个视野,计算细胞总数和凋亡细胞总数,以凋亡细胞总数/细胞总数计算出细胞凋亡率。
1.9 裸鼠移植瘤实验检测Runx2表达联合X射线照射对细胞的生长抑制作用 选取45只体质量和日龄相近的裸鼠,随机平均分3组(每组15只),即对照组、基因下调组和基因上调组。取转染48 h后的3组细胞胰酶消化重悬,分别接种于相应组裸鼠右腋皮下0.5 cm处。裸鼠在无菌、通风、洁净的SPF级动物房中饲养,接种后1周左右,在裸鼠的接种区出现米粒大小的移植瘤。各组裸鼠麻醉,并固定于解剖板。放疗应用上述6MV X射线,皮源距(SSD)为100 cm,照射野为2 cm×2 cm,吸收剂量率200 cGy/min,2 Gy/次。1次/d,5次/周,总剂量20 Gy。40 d后处死裸鼠,分离肿瘤组织,计算肿瘤瘤块体积。该实验重复进行3次。
1.10 统计学处理 SPSS16.0进行统计分析;并用表示,多组间均数采用单因素方差分析,两组组间比较采用t检验,3组数据间两两比较采用SNK-Q检验,以P<0.05为差异具有统计学意义。
2 结果
2.1 直肠癌及癌旁组织中Runx2 mRNA的表达 图1所示为31例直肠癌患者癌组织和相应癌旁组织中Runx2基因相对表达量。在直肠癌癌组织中的该分子基因相对表达水平为0.83±0.31,癌旁组织中为0.43±0.25(P<0.05)。
2.2 转染48 h后Runx2 mRNA的表达 表1、图2所示为转染48 h后,3组Runx2基因和蛋白相对表达水平,与对照组相比,下调组细胞Runx2的基因相对表达水平显著降低,上调组中细胞中的Runx2 mRNA表达水平则显著升高(P<0.05)。

图1 直肠癌及癌旁组织中Runx2 mRNA的表达()
表1 转染后Runx2 mRNA与蛋白的表达(,n=3)

表1 转染后Runx2 mRNA与蛋白的表达(,n=3)
注:与对照组相比,aP<0.05
Runx2蛋白相对表达水平对照组 0.88±0.09 1.09±0.08 Runx2下调组 0.23±0.04a 0.21±0.06a Runx2上调组 1.47±0.09a 1.56±0.09a组别 Runx2基因相对表达水平
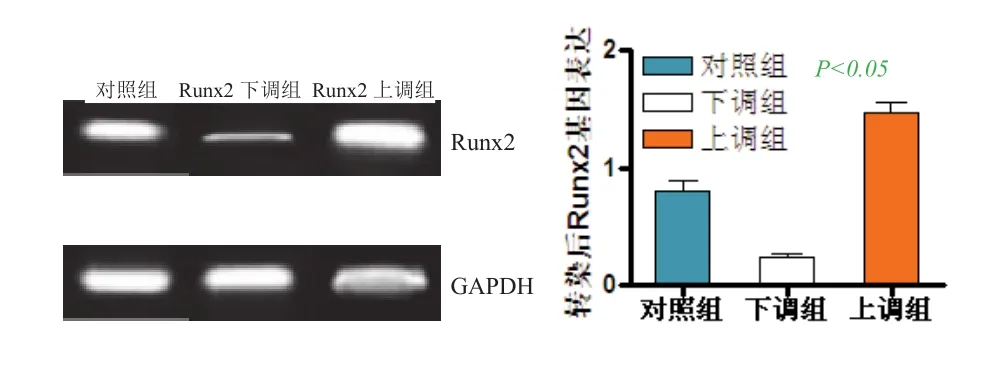
图2 转染后Runx2 mRNA的表达()

图3 转染后Runx2蛋白的表达()
2.4 CCK-8检测细胞增殖 如表2所示为癌细胞增殖活性的变化。CCK-8结果显示,与对照组相比,Runx2基因沉默组直肠癌LOVO细胞增殖水平显著降低(P<0.05),而上调组中直肠癌细胞的增殖水平则明显升高。与Runx2上调组相比,Runx2下调组细胞增殖水平显著降低。见图4。

表2 各组直肠癌LOVO细胞增殖情况(,n=3)

图4 各组直肠癌LOVO细胞增殖情况()
2.5 Runx2表达对LOVO的放射敏感性的影响 表3、图5所示为基因转染后直肠癌LOVO细胞经过放疗后。与对照组相比,Runx2下调组细胞平均致死剂量D0、SF2和N值明显降低,Runx2上调组细胞的D0、SF2和N值显著升高(P<0.05)。
组别 D0 N SF22.6 细胞的凋亡 与对照组(24.12±1.18)%比 较, 上 调 组(13.23±0.37)% 和 下 调 组(58.16±0.92)%癌细胞中Runx2的表达水平对直肠癌LOVO细胞凋亡率的影响。Runx2基因沉默显著促进了直肠癌LOVO细胞的凋亡,上调组则明显降低了直肠癌LOVO细胞的增殖活化能力。见图6。
表3 各组直肠癌细胞株LOVO的放射敏感性(,n=3)

表3 各组直肠癌细胞株LOVO的放射敏感性(,n=3)
注:与对照组相比,aP<0.05
对照组 0.58±0.08 1.36±0.13 0.65±0.08 Runx2下调组 0.43±0.05a 0.98±0.07a 0.47±0.06a Runx2上调组 0.65±0.09a 2.67±0.29a 0.93±0.12a

图5 各组直肠癌细胞株LOVO的放射敏感性()

图6 各组直肠癌LOVO细胞的凋亡率()
2.7 Runx2表达联合X射线照射对细胞的生长抑制作用 Runx2基因沉默组裸鼠移植瘤的平均体积(2.53±0.23)cm3明显小于对照组(3.82±0.31)cm3,其抑瘤率显著升高(P<0.05);上调直肠癌细胞中Runx2相对表达之后,裸鼠皮下种植瘤平均体积明显变大(5.21±0.21)cm3,抑瘤率显著降低。见图7。

图7 各组移植瘤平均体积比较
3 讨论
Runx2可能会增加直肠癌细胞的放射敏感性。目前关于Runx2在直肠癌细胞中的表达水平变化方面的研究还较为缺乏,本研究首先探讨Runx2在直肠癌LOVO细胞中是否存在异常表达,并通过基因转染双向调控直肠癌细胞中的Runx2的表达,研究上调和下调Runx2在直肠癌细胞中的相对表达水平对移植瘤放射敏感性的影响,以期为直肠癌的治疗提供借鉴和依据。
Runx2是近年来医学界研究的一大热点,其可以通过调控多种基因转录进而影响骨骼的形成、骨细胞分化等[10]。近年来研究表明,Runx2在肿瘤细胞的进展中也有重要的作用,给癌症的防治又打开了一扇大门。Runx2在调控肿瘤细胞的侵袭和转移能力方面有特殊作用[11-12],尤其在促进肿瘤细胞向骨中转移进程中作用显著[13]。这可能因为Runx2可以介导与癌细胞浸润转移相关的金属基质蛋白酶异常表达、肿瘤血管生成和与癌细胞侵袭相关的MAPK蛋白表达水平升高[14-15]。就通过基因转染Runx2研究其与直肠癌细胞存活与凋亡关系研究尚少,而且就其与直肠癌移植瘤放射敏感性的关系也比较缺乏。
本研究通过荧光定量,探讨了Runx2 在直肠癌细胞组织的基因相对表达水平情况。结果显示,其在癌组织中的相对表达水平明显高于癌旁组织。该结果与张海元等[16]就Runx2蛋白与结直肠癌和大肠癌关系的研究较为一致。通过电穿孔法基因转染Runx2发现,Runx2下调组直肠癌LOVO细胞内Runx2基因表达明显降低,Runx2上调组明显升高,这表明基因转染成功。Runx2基因下调组直肠癌细胞增殖活性被抑制,而细胞凋亡率增加;Runx2上调组中则有相反变化出现。研究显示,Runx2介导的癌细胞的增殖与凋亡可能与调控细胞增殖的相关基因有关[17]。本研究结果显示,Runx2下调组直肠癌LOVO细胞放射敏感性升高,相较而言,Runx2上调组直肠癌LOVO细胞放射敏感性降低。下调Runx2表达后,经过射线放疗处理,癌细胞的移植瘤瘤体体积显著降低,该瘤体得到了显著的抑制;上调Runx2的表达则与之效应相反。提示直肠癌LOVO细胞的放射敏感性与Runx2的表达水平关系密切,通过基因沉默可以提高癌细胞的放射敏感性,增强放射治疗的疗效。
Runx2基因调控对直肠癌放射敏感性的影响机制尚不可知。刘勇等[18]应用免疫组化法检测局部进展期直肠癌患者术前同步放化疗前、后的凋亡相关蛋白,结果表明放化疗组中Bax表达变化与肿瘤肌层浸润和放疗后肿瘤细胞退缩之间差异有显著性意义。提示放射治疗直肠癌与癌细胞中的Bax的表达水平有关。已有研究表明,Runx2参与Bax、Bcl-2等信号通路调控[19-20]。由此可知,Runx2可能通过调节这些信号通路从而对直肠癌细胞的放射敏感性发挥影响,有待进一步研究探讨。
本研究通过基因转染技术分别上调和下调调控直肠癌细胞中的Runx2基因和蛋白的表达,提高了直肠癌细胞的放射敏感性。基因手段和放疗手段相结合,显著抑制了移植瘤的生长,但其机制还有待进一步探讨研究。
[1] Zuo B,Zhu JF,Li J,et al.microRNA-103a functions as a mechanosensitive microRNA to inhibit bone formation through targeting Runx2[J].J Bone Miner Res, 2015,30(2):330-345.
[2] Weiser MR,Fichera A,Schrag D,et al.Progress in the PROSPECT trial: precision treatment for rectal cancer [J].Bull Am Coll Surg,2015,100(4):51-52.
[3] Qiu J,Yong T,Yang G,et al.Effect of a chemical inhibitor of human phosphatidylethanolamine-binding protein 4 on radiosensitivity of rectal cancer cells[J].World J Surg Oncol,2016,14(1):221.
[4] 裘建明,杨关根,沈忠,等.CD133+直肠癌细胞对直肠癌放射线敏感性的影响[J].中华肿瘤杂志,2014,36(2):103-107.
[5] 李庆庆,黄江鸿,何关健,等.Runx2及其在骨组织工程中的应用[J].国际骨科学杂志,2014,35(1):44-46.
[6] 熊加秀,叶棋浓,卫勃,等.敲低Runt相关转录因子2(Runx2)表达抑制结肠癌细胞生长和迁移[J].细胞与分子免疫学杂志,2016,32(7):950-953,957.
[7] 耿文文,张斌,曹旭晨.Runx2与肿瘤转移[J].国际肿瘤学杂志,2012,39(12):892-894.
[8] Browne G,Nesbitt H,Ming L,et al.Bicalutamide-induced hypoxia potentiates RUNX2-mediated Bcl-2 expression resulting in apoptosis resistance[J].Br J Cancer, 2012,107(10):1714-1721.
[9] 阮婷彦,楚建军,金建荣,等.人肝癌细胞BEL-7402非常规照射模式生物效应探讨[J].肿瘤防治研究,2010,37(10):1128-1131.
[10] Taipaleenmaki H,Browne G,Akech J,et al.Targeting of Runx2 by miRNA-135 and miRNA-203 impairs progression of breast cancer and metastatic bone disease[J].Cancer Res, 2015,75(7):1433-1444.
[11] Zhu Y,Zhao H,Feng L,et al.MicroRNA-217 inhibits cell proliferation and invasion by targeting Runx2 in human glioma[J].Am J Transl Res,2016,8(3):1482-1491.
[12] Zhang R,Yan S,Wang J,et al.MiR-30a regulated the proliferation,migration,and invasion of human osteosarcoma by targeting Runx2[J].Tumour Biol,2015,37(3):3479-3488.
[13] 王剑,郑洪新,刘妍,等.补肾益髓中药复方对去卵巢骨质疏松症大鼠骨质疏松症大鼠骨组织Runx2 mRNA及蛋白表达的影响[J].中国骨质疏松杂志,2014,8:890-895.
[14] 洪峰,郑冲,徐德淦,等.慢性氟砷联合暴露对大鼠骨骼Runx2及其下游相关因子的影响[J].中华预防医学杂志,2013,47(9):794-798.
[15] Wang ZQ,Keita M,Bachvarova M,et al.Inhibition of RUNX2 Transcriptional Activity Blocks the Proliferation,Migration and Invasion of Epithelial Ovarian Carcinoma Cells[J].PLoS One,2013,8(10):108-110.
[16] 张海元,刘娟.结直肠癌细胞系中Runx3及Runx2基因表达分析[J].长江大学学报:自科版,2008,5(2):1-3.
[17] Niu DF,Kondo T,Nakazawa T,et al.Transcription factor Runx2 is a regulator of epithelial-mesenchymal transition and invasion in thyroid carcinomas[J].Lab Invest, 2012,92(8): 1181-1190.
[18] 刘勇,张谷,钱俊,等.局部进展期直肠癌术前新辅助治疗后凋亡相关蛋白检测及临床意义[J].中华外科杂志,2012,50(10):914-917.
[19] Byon CH,Sun Y,Chen J,et al.Runx2-upregulated receptor activator of nuclear factor κB ligand in calcifying smooth muscle cells promotes migration and osteoclastic differentiation of macrophages[J].Arterioscler Thromb Vasc Biol, 2011,31(6):1387-1396.
[20] Ho WP,Chan WP,Hsieh MS,et al.Runx2-mediated bcl-2 gene expression contributes to nitric oxide protection against hydrogen peroxide-induced osteoblast apoptosis[J].J Cell Biochem, 2009,108(5):1084-1093.
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
山东冶金(2022年4期)2022-09-14
现代仪器与医疗(2021年4期)2021-11-05
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年5期)2021-07-22
世界科学技术-中医药现代化(2021年12期)2021-04-19
甘肃教育(2020年4期)2020-09-11
电子制作(2019年22期)2020-01-14
奥秘(创新大赛)(2019年9期)2019-10-09
小哥白尼(趣味科学)(2019年1期)2019-04-12
