
长江口东滩湿地不同盐沼植被样地内浮游植物群落结构的差异
2018-06-29 02:54叶锦玉张瑛瑛彭建新张建明
海洋渔业 2018年3期
张 衡, 叶锦玉, 张瑛瑛, 康 伟, 彭建新, 张建明
(1.中国水产科学研究院东海水产研究所,农业部东海与长江口渔业资源环境科学观测实验站, 上海 200090; 2. 上海海洋大学海洋科学学院,上海 201306; 3. 江苏省海洋渔业指挥部,南通 226006 )
浮游植物作为海洋生态系统的重要初级生产力,是河口及邻近水域渔业环境质量评估的重要指标之一[1-4]。长江口浮游植物的群落特征已有较多的报道,主要涉及种类数量[2-4]、生态区系[5-7]等方面,但针对长江口盐沼植被生境的水体浮游植物群落特征研究还未见报道。
除了河口近岸水体及光滩碎波带是河口鱼类的重要育幼场[8-12],已有证据表明,盐沼植被覆盖区也是鱼类(特别是幼鱼)的重要栖息地,一些植食性鱼类或虾类的幼体进入盐沼植被覆盖区躲避敌害并能获取相应的饵料(主要是浮游植物或底栖硅藻),如鲻科鱼类/白虾属等虾类生物[12-13]。盐沼改变了水体的水动力条件,使得水流流速相对缓慢,有利于浮游植物的群聚,不同的植被由于所处的高程不同且生长特征不一,使得植被样地内的浮游植物群落特征也存在差异,与邻近的光滩水域和潮下带水域浮游植物群落也存在较大差异,但对此研究报道极少[14]。
长江口东滩湿地主要生长有芦苇 (Phragmitesaustralis)和海三棱藨草 (Scirpusmariqueter)等土著植被,但自1995年引种互花米草 (Spartinaalterniflora)至崇明东滩以来,由于其强适应性、扩散能力和竞争力,严重威胁着崇明东滩生物多样性与生态安全[15-16]。互花米草早期阶段会入侵至海三棱藨草群落,造成后者生境呈斑块化,而互花米草入侵后期阶段则直接占领了芦苇和海三棱藨草的原有生态位[15]。目前,关于互花米草入侵对于土著植被生长的负面影响以及不同植被样地内底栖动物和鸟类的分布差异已有较多的报道[13, 15-16],但对于不同盐沼植被样地内的浮游植物群落差异影响还未见报道。盐沼植被水体的浮游植物与邻近潮下带水域的浮游植物是否存在差异尚不明确。本研究根据2015年5-12月的调查资料分析盐沼植被区水体浮游植物的种类和丰度的变化,了解该水域的浮游植物群落季节变化特征,以期初步掌握这一特殊生境的浮游植物群落变化和区系特征。
1 材料与方法
于2015年5-12月每月在长江口东滩湿地东北水域(即东旺沙水域,经纬度为31°34′51″~31°34′59″N、121°54′27″ ~121°54′36″E)的海三棱藨草(站位编号HS)、互花米草入侵早期(站位编号HZ)和入侵后期阶段(站位编号HH)、芦苇(站位编号LW)等4种植被样地采集水体浮游植物样本,由海向陆的方向沿高程大小不同把互花米草盐沼划分入侵早期(入侵期1~2年)和入侵后期(入侵期>5年)2类。其中,HS和HZ植被样地为同一高程,两者相距300 m;HH和LW植被样地为同一高程,两者相距300 m。LW植被样地高程相比HS植被样地高约0.9 m。
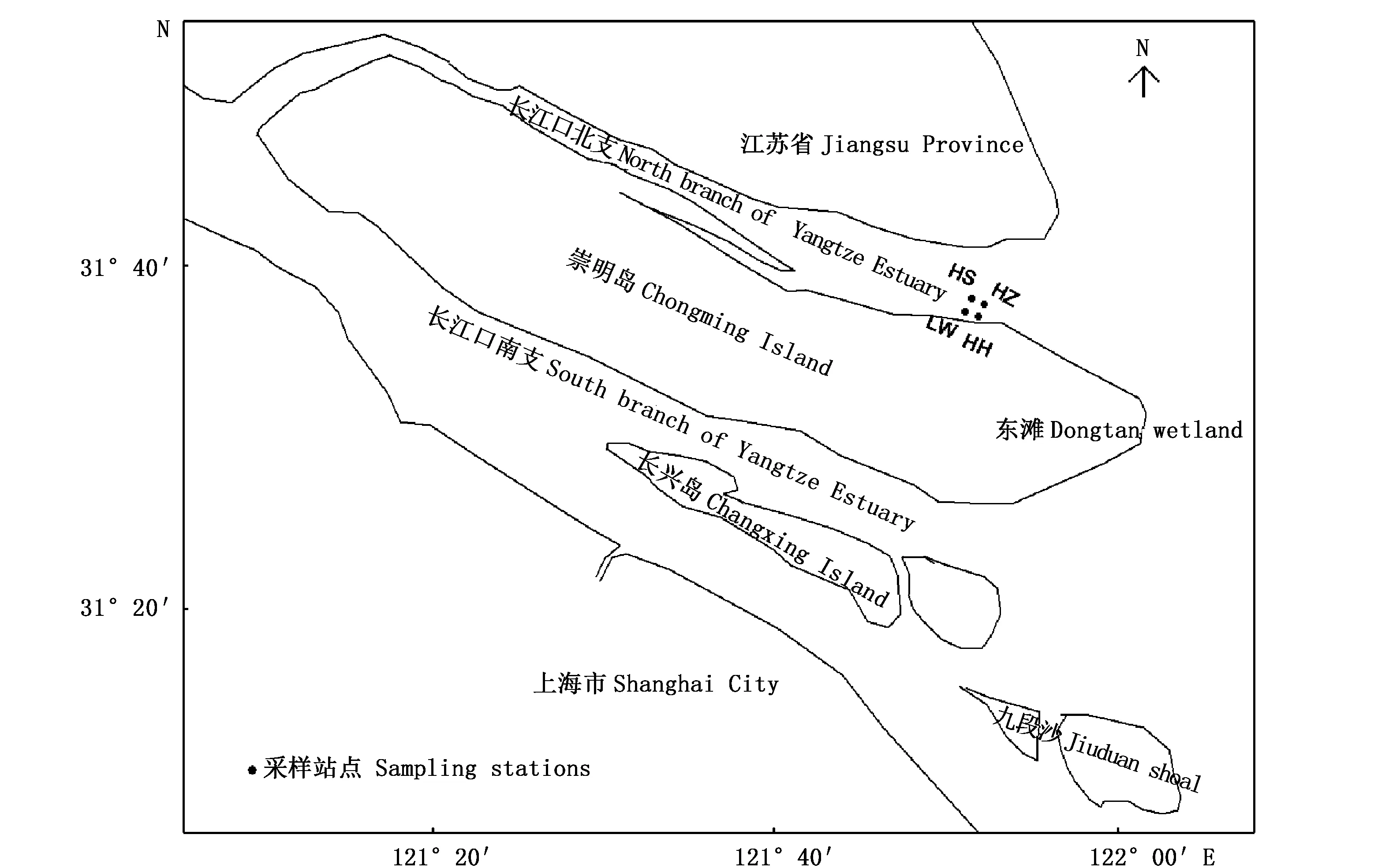
图1 采样站位示意图(●代表站点;HS,海三棱藨草;HZ,互花米草入侵早期阶段;HH,互花米草入侵后期阶段;LW,芦苇) Fig.1 Sampling stations (●) of 4 types of salt marshes (HS,Scirpus mariqueter; HZ, early stage of Spartina alterniflora invasion; HH, later stage of Spartina alterniflora invasion; LW, Phragmites australis )
每月大潮期在各站点采3次重复样,采样均在白天潮水涨憩时进行,同时进行水深(精确到0.1 m)、水温(精确到0.1 ℃)和盐度测量。在0.3 m水深处取1 L水样,并加5%浓度的福尔马林溶液保存,带回实验室后立即鉴定种类和计数。取25 mL浓缩后的浮游植物样品于Hydro-bios的Utermöhl计数框, 静置沉降12 h,然后用AO 型号倒置显微镜, 在200 或400倍下进行物种鉴定与计数。每个样品分别取3次平行样计数,取平均值作为该站位的浮游植物细胞丰度(cell· L-1)。细胞丰度的统计和误差处理参考孙军等[17]。
根据公式(1)计算物种的优势度(Y):
(1)
式(1)中,fi为第i种的出现频次(%),ni为第i种的丰度,N为所有物种的总丰度。
采用Margalef 物种丰富度指数(D)、Pielous种类均匀度指数(J)、Shannon-Weaver指数(H′)进行浮游植物群落多样性分析[18-20]:
D=(S-1)/log2N
(2)
(3)
(4)
式(2)~式(4)中,S为种类数,ni为第i种的丰度,N为总丰度,H′max=log2S。
采用SPSS 17.0统计软件分析不同植被样地浮游植物丰度变化的显著性差异(One-way ANOVA)。对浮游植物丰度数据进行四次方根转换后进行月份间Bray-Curtis相似性系数计算,采用等级聚类(即组平均法)的分类方法和非度量多维标序(non-Metric Multidimensional Scaling,简称nMDS)的排序方法分析其群落结构特征。由于这两种方法的自然互补,它们可共同成为分析群落结构数据的有效工具,并互相验证结果的正确性。检验nMDS分析结果的优劣用胁强系数(Stress)来衡量,通常认为Stress<0.2时,可用nMDS的二维点图表示,其图形有一定的解释意义;Stress<0.1时,可认为是一个好的排序,Stress<0.05时,认为具有很好的代表性[19-20]。应用相似性分析(Analysis of similarities,简称ANOSIM)检验各季节浮游植物群落组成的零假设,组内各站位的平均秩相似性与组间所有站位间的平均秩相似性之间的差异用R表示,其阈值为-1~1[20]。另外用SIMPER方法计算各种类对样本组内相似性和组间相异性的平均贡献率[19],以确定各样本组的指示种。上述分析皆使用PRIMER 7.0软件进行。
2 结果与分析
2.1 环境特征
调查期间,水温呈明显的季节变化特征,5-12月变化范围为5.0~30.5 ℃,5-8月水温逐渐升高,从20.1 ℃上升到30.5 ℃;9-12月水温逐渐下降,从22.8 ℃下降至5.0 ℃。盐度变化十分显著,5-12月变化范围为3.2~16.8,其中夏季6-8月盐度较低,变化范围为3.2~4.3,9-11月有所上升,12月达到最高值(16.8)。5-10月是海三棱藨草、芦苇和互花米草的主要生长季,而11月以后3种植被逐渐枯萎。调查期间,5-7月的白天最大潮高是3.4~3.7 m,8-10月逐渐增加(10月达到最高值为4.6 m),11月有所降低,12月降低至4.1 m。
2.2 种类
调查期间,共发现浮游植物64种(或属),隶属4个门;其中有12种鉴定到属。硅藻门种类最多,有48种(或属),占总种类数的73.8%;其次为蓝藻门种类,有10种(或属);绿藻门有6种,而甲藻仅发现1种(表1)。HS植被样地采集到浮游植物38种,HZ植被样地采集到35种,而HH和LW植被样地分别采集到38和36种。从出现频次来看,硅藻门的中肋骨条藻(Skeletonemacostatum)、洛氏菱形藻 (Nitzschialorenziana)、近缘斜纹藻 (Pleurosigmaaffine)和舟形藻属 (Naviculasp.)等出现频次较高,而蓝藻门的颤藻属 (Oscillatoriasp.)和极大节旋藻(Arthrospiramaxima)出现频次较高。各个站位的绿藻和甲藻门的种类出现频次均低于15%。从丰度来看,各个站位均以中肋骨条藻丰度最高,其次为洛氏菱形藻和近缘斜纹藻。综合来看,该水域浮游植物优势种为中肋骨条藻(Y=0.76)、洛氏菱形藻(Y=0.03)和近缘斜纹藻(Y=0.02)等3种;偶见种为齿角毛藻 (Chaetocerosdenticulatus)、蜂腰双壁藻 (Diploneisbombus)、大螺旋藻 (Spirulinamajor)和小球藻 (Chlorellavalgaris)等。
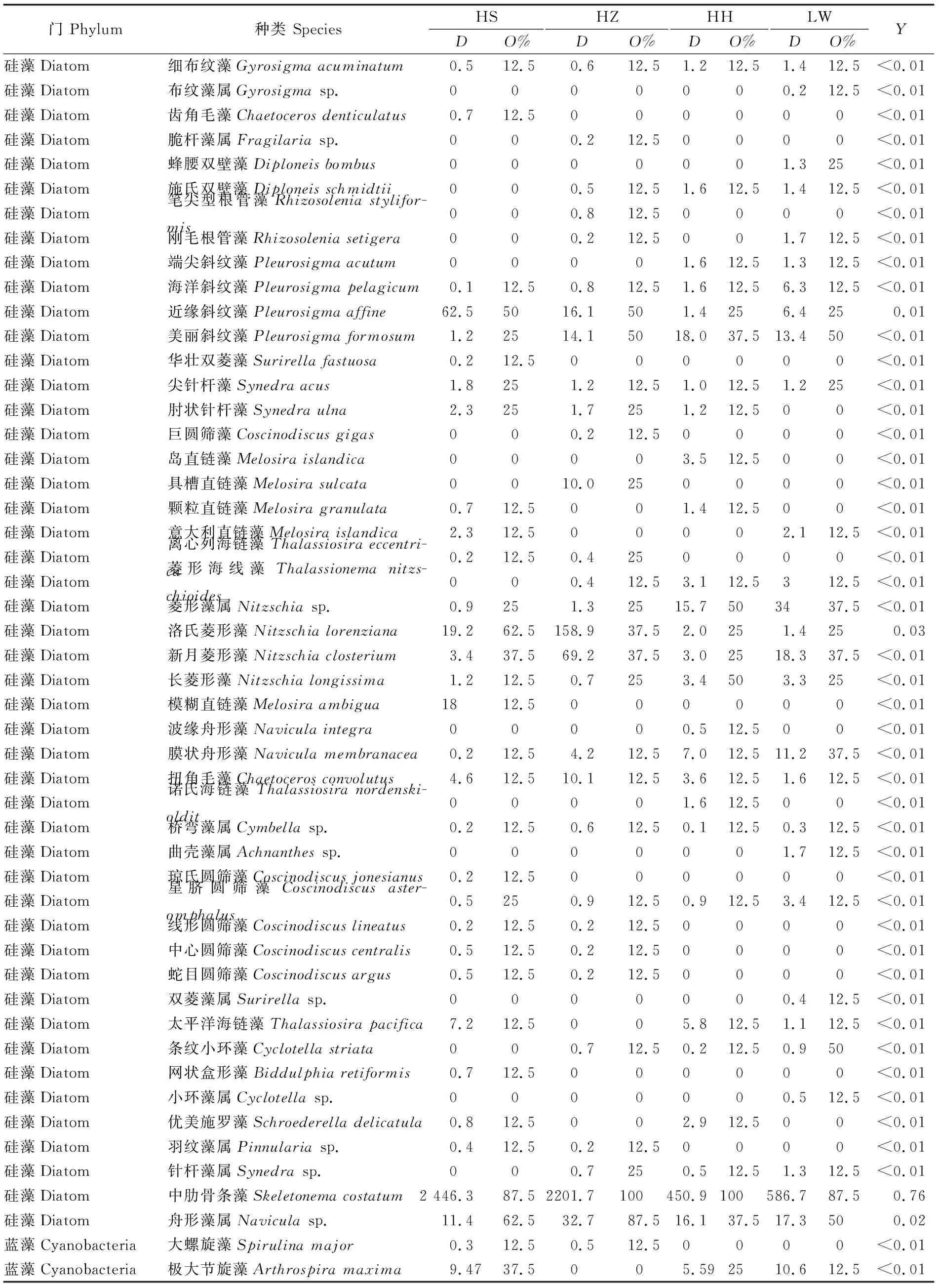
表1 监测站点浮游植物的月平均丰度(×104 cell· m-3)、出现频次(%)和优势度(Y)Tab.1 Mean density (×104 cell · m-3), occurrence frequency(%) and dominance of phytoplankton(Y) in the sampling stations
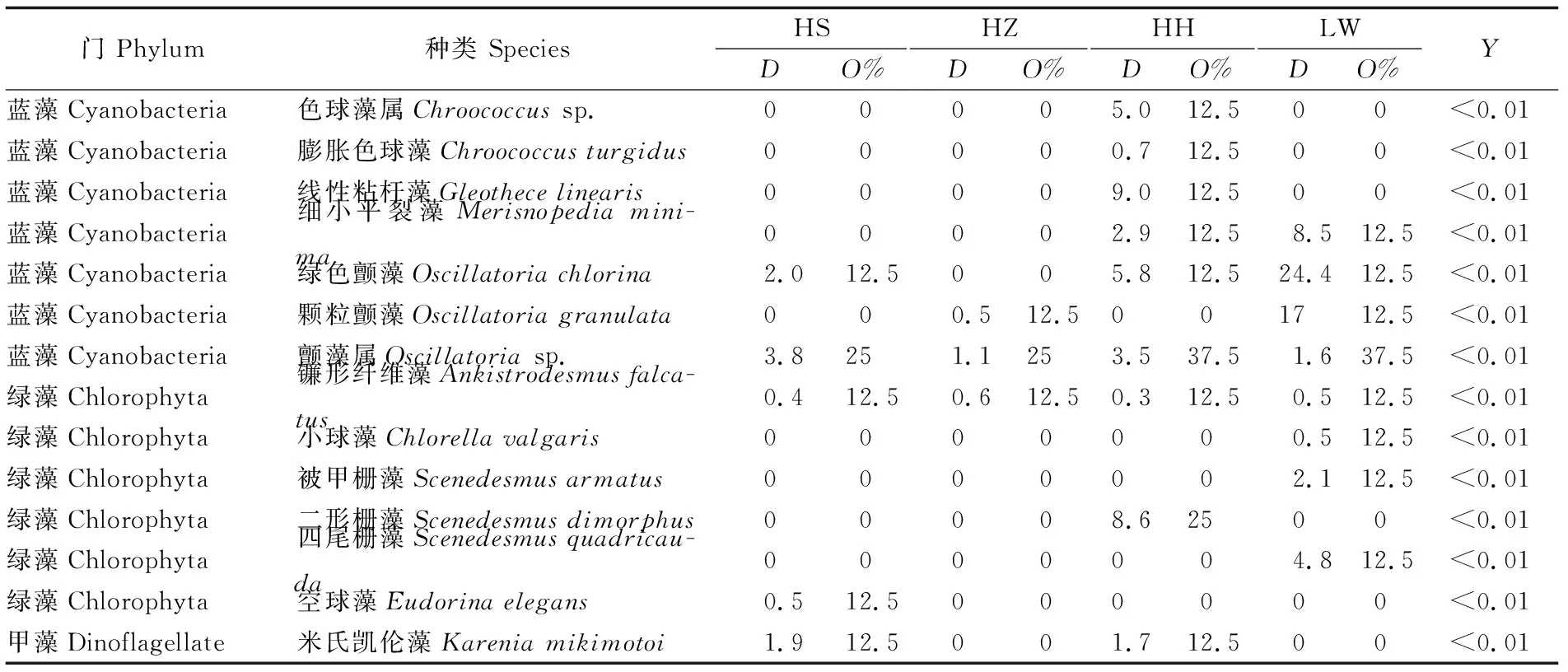
·续上表·
注:HS,海三棱藨草;HZ,互花米草入侵早期;HH互花米草入侵后期;LW,芦苇;D: 丰度;O%,出现频次;Y,浮游植物优势度
Note:HS,Scirpusmariqueter; HZ, early stage ofSpartinaalterniflorainvasion; HH, later stage ofSpartinaalterniflorainvasion; LW,Phragmitesaustralis;D, density;O%, occurrence frequency;Y, dominance of phytoplankton
4种植被样地均以硅藻门种类数最多,其次是蓝藻门,而甲藻门种类数最低;硅藻与其它藻类的种类数均呈显著性差异(P<0.01)(图2)。各个植被样地中硅藻的月平均丰度均明显高于其它藻类,而甲藻丰度最低。HS和HZ植被样地中的浮游植物群落的种类数和丰度并无显著性差异(P>0.05),HH和LW植被样地浮游植物的种类数和丰度同样没有显著性差异(P>0.05);前2种植被样地浮游植物的月平均丰度约是后2种植被样地的3.8倍(P<0.01),这主要是由于硅藻的丰度差异较大造成的(图2)。
2.3 月变化
5-8月盐沼植被区浮游植物的种类数相对较高,而9-12月(秋季至冬初)种类数明显下降(图3)。浮游植物月平均丰度在7月出现最大峰值,达8.96×104cell· L-1;8-11月丰度从0.54×104cell· L-1逐渐上升到1.11×104cell· L-1,12月则急剧下降至0.05×104cell· L-1,相比11月丰度下降了约95%。
从图4可知,4种植被样地的浮游植物种类数从5-12月均呈现逐渐下降趋势,且各个植被样地浮游植物种类数无显著性差异(P>0.05)。从丰度来看,HS和HZ植被样地的浮游植物月平均丰度明显高于HH和LW植被样地,特别是6月和7月;而其它月份未发现显著性差异(P>0.05)。4种植被样地的浮游植物均以冬初(12月)丰度最低,而其它月份丰度相对较高(图4)。
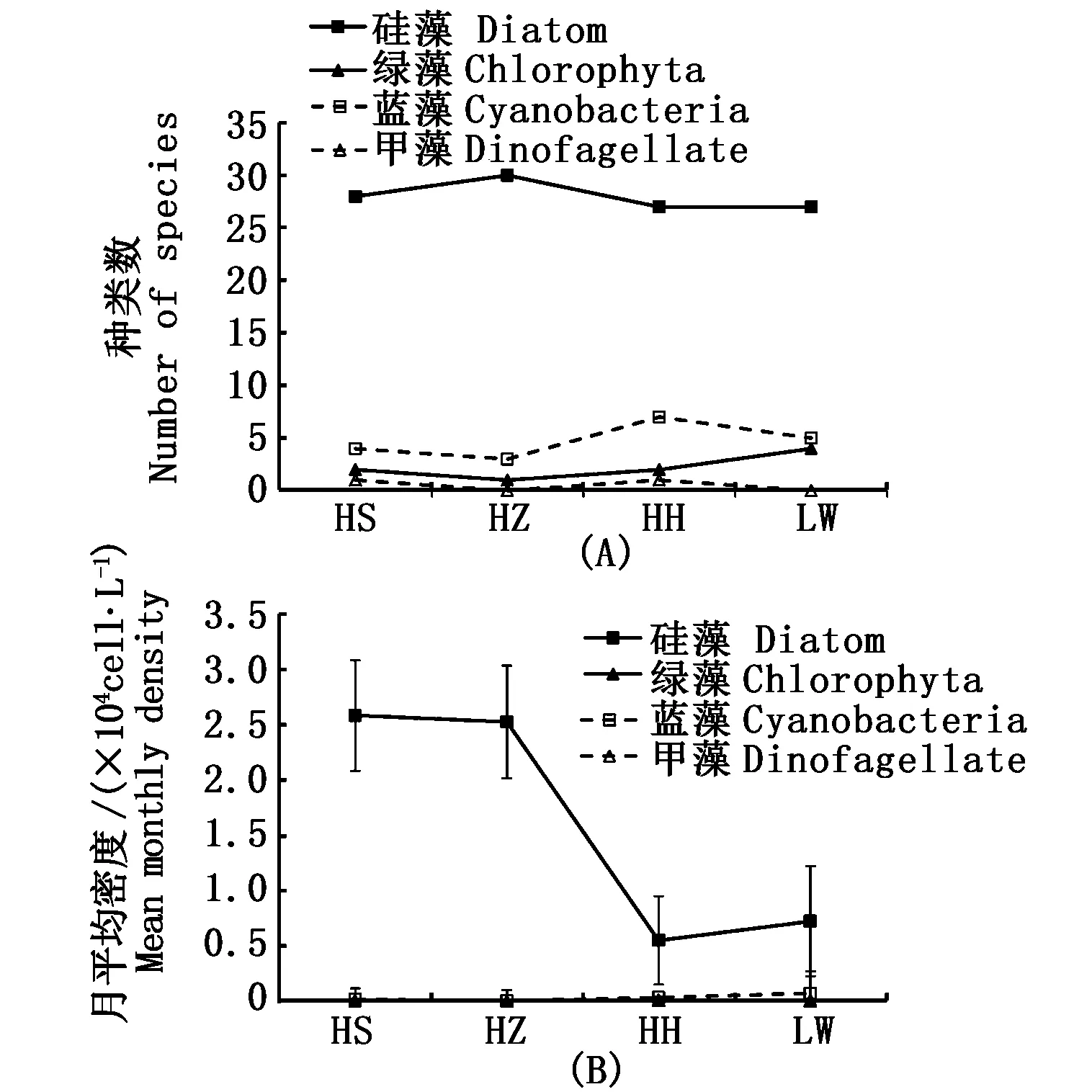
图2 四种植被样地的浮游植物种类数(A)和月平均丰度(B)的变化Fig.2 Changes in the species (A) and mean monthly density (B) of phytoplankton in 4 types of salt marshes
从物种丰富度指数(D)来看,5-8月4种植被样地浮游植物D值较高,而9-12月D值较低;两两植被样地间物种丰富度指数未发现显著性差异(P>0.05)(图5)。10月和11月均匀度指数(J)平均值明显低于其它月份;7月(HS和HZ)和9月(HZ和HH)植被样地浮游植物的J值均小于0.1。Shannon 指数(H′)变化趋势类似于均匀度指数(J),5-8月和12月H′值相对较高,但因7月(HS和HZ)植被样地浮游植物的优势种中肋骨条藻丰度过高导致其H′值均低于0.2(表1,图5)。
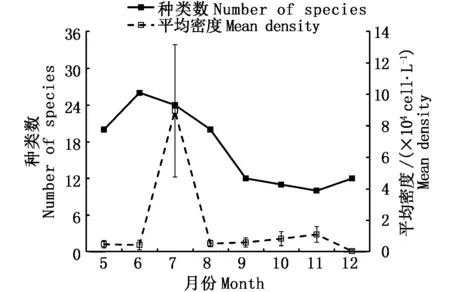
图3 长江口盐沼植被区浮游植物群落种类数和平均丰度的月变化Fig.3 Monthly changes of mean number of species and density for phytoplankton in salt marshes in the Yangtze Estuary
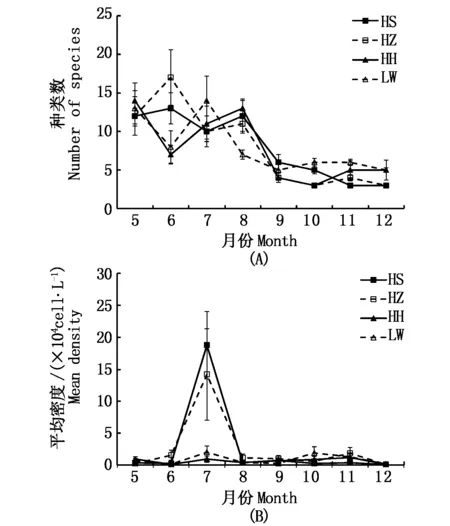
图4 4种植被样地中浮游植物的种类数(A)和平均丰度(B)的月变化Fig.4 Monthly changes of number of species (A) and mean density (B) of phytoplankton in 4 types of salt marshes
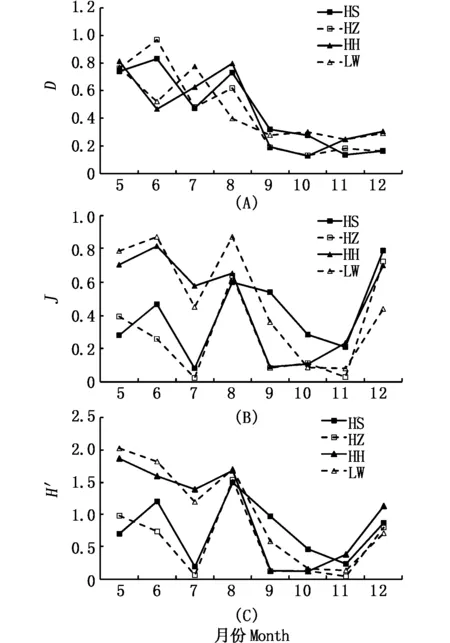
图5 4种植被样地中浮游植物群落多样性月变化 [物种丰富度指数D (A), Pielous种类均匀度指数J(B),Shannon-Weaver指数H′ (C)]Fig.5 Monthly changes in biodiversity of phytoplankton in 4 types of salt marshes (Margalef’s species richness index (D) (A), Pielou’s index (J') (B)and Shannon-Weaver index (H′) (C))
2.4 聚类和排序
从等级聚类(即组平均法)的分类方法可知,浮游植物群落在43%相似性水平上可分为两组,A组为9-12月(包括HS5、HZ5站位),B组为5-8月(包括HS12站位)(图6)。从nMDS排序图也可知,7月和8月的4个植被样地站位以及HZ6、HH5、LW5等站位与其它月份的站位排序差异较大(图7)。经ANOSIM检验可知,在时间分布上,A 、B组浮游植物群落差异显著(R=0.32,P<0.01);而在空间分布上各植被样地无显著差异(R=-0.01,P>0.05)
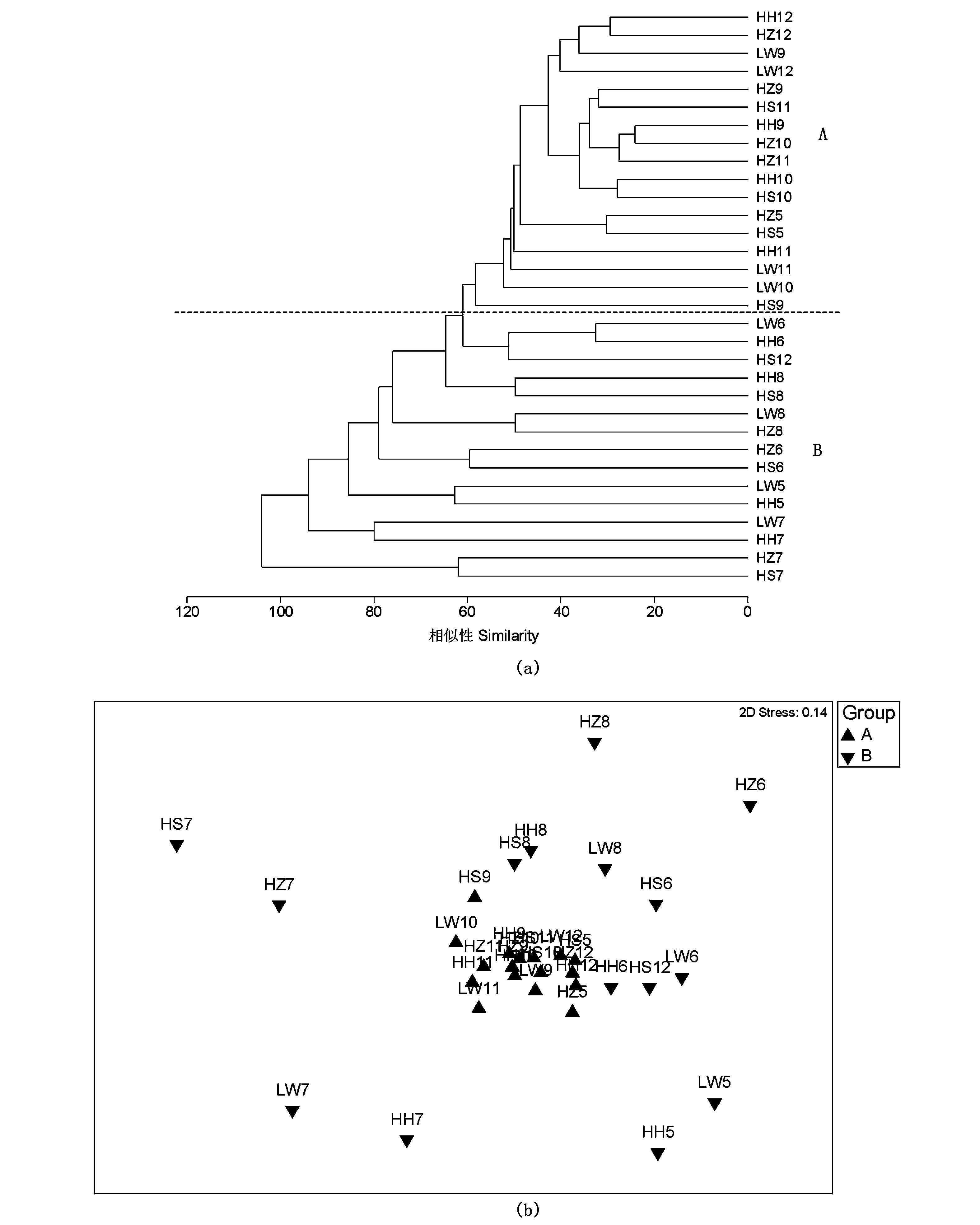
图6 4种植被样地中浮游植物群落的Bray-Curtis similarity聚类(a)和nMDS排序图(b)(HS 5~12:代表5-12月海三棱藨草植被样地内样本;HZ5~12:代表5-12月互花米草入侵早期阶段植被样地内样本;HH5~12:代表5-12月互花米草入侵后期阶段植被样地内样本;LW5~12:代表5-12月芦苇植被样地内样本;Group A:9-12月,Group B:5-8月)Fig.6 Analysis on Bray-Curtis similarity CLUSTER (a)and nMDS (b) on abundance of phytoplankton community in 4 types of salt marshes (HS 5~12: samples in Scirpus mariqueter from May to December; HZ 5~12: samples at the early stage of Spartina alterniflora invasion from May to December; HH 5~12: samples at the later stage of Spartina alterniflora invasion from May to December; LW 5~12: samples in Phragmites australis from May to December;Group A:Sepetmber-December,Group B:May-August)
SIMPER过程分析的结果表明,A、B组间种类组成的平均相异性为52.65%,21种藻类对平均相异性的累积贡献率>70%,主要献率来自中肋骨条藻、舟形藻属、美丽斜纹藻(Pleurosigmaformosum)、近缘斜纹藻、洛氏菱形藻和新月菱形藻(Nitzschiaclosterium)等(表2)。其中,中肋骨条藻对A和B组群落的贡献率均最大,达31%以上。
3 讨论
3.1 种类和丰度
长江口附近水域浮游植物物种丰富,丰度极高,生态类型多样化[1-7, 21]。一些研究表明,长江口共有浮游植物约300多种[22],仅崇明东滩附近水域就多达130种[3]。但以往研究多涉及潮下带浮游植物群落,极少涉及到潮间带水域。本研究虽只调查了东滩湿地东北水域植被区浮游植物群落,就已发现64种(属),隶属4门,月平均丰度高达1.63×104cell · L-1,说明盐沼植被区浮游植物物种同样丰富,可为盐沼生态系统提供基础饵料和初级生产力。
长江口口门区浮游植物优势种以硅藻门的中肋骨条藻和圆筛藻属为主[1, 3],长江口最大浑浊带水域以根管藻属、圆筛藻属、中肋骨条藻为优势种[2],而长江口外海则以硅藻门的菱形海线藻(Thalassionemanitzschioides)、中肋骨条藻和甲藻门的纺锤角藻(Ceratiumfusus)等为优势种[23]。本研究发现,东滩东北部盐沼植被样地浮游植物群落同样以中肋骨条藻为绝对优势种,其次为洛氏菱形藻和近缘斜纹藻,而蓝藻以极大节旋藻和颤藻属为优势种(类),绿藻以二形栅藻(Scenedesmusdimorphus)等栅藻属为优势种(类),这与前人稍有区别,因研究区域与前人研究差异较大导致。张松艳等[14]在长江口南支潮间带碎波带水域进行了周年调查,发现硅藻同样是优势门类,但优势种以小环藻属为主要优势种,其次是中肋骨条藻,与本研究有所差异,这是因为长江口南支水域以淡水为主,而本研究调查水域以咸淡水为主。
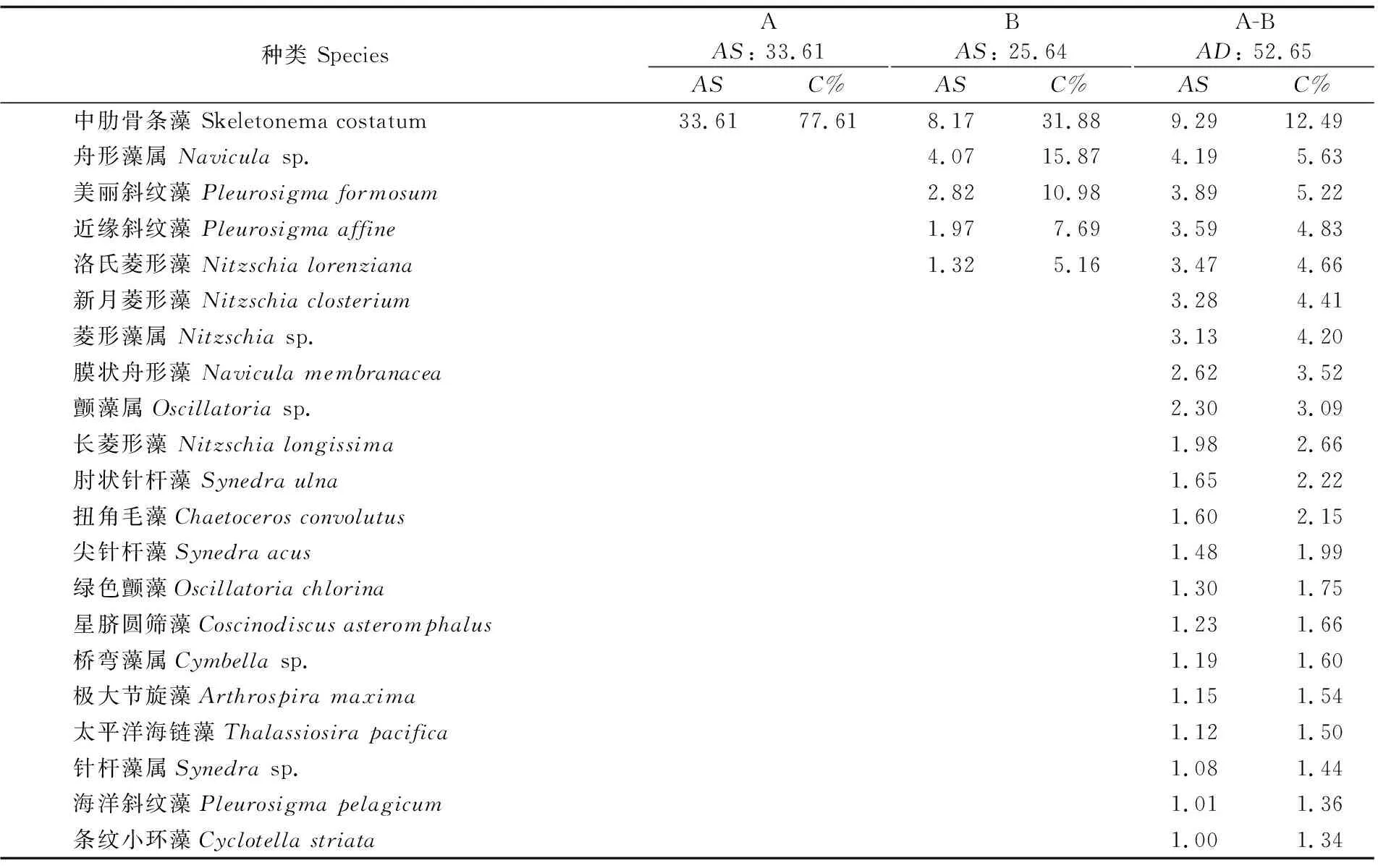
表2 A、B组组内相似性和组间相异性贡献率较大的物种(总贡献率>70%)Tab.2 Species with accumulated contribution>70% of similarity within a group and dissimilarity between group A and B
注:AS为组内平均相似性,AD为组间相异性,C%为各物种对组内相似性或组间相异性的贡献率
Note:AS: average similarity within group,AD: average dissimilarity between groups,C%: contribution of similarity within group and dissimilarity between groups by each species
盐沼植被样地的浮游植物主要随着潮水的涨落进入或退出盐沼生境,与长江口口门区和口外区的丰度相比稍低[22-23],也低于长江口南支碎波带水域的平均丰度(37.70×104cell· L-1)[14],这可能是因为浮游植物在进入盐沼植被区时会有一部分附着在植被和潮间带底质上。有研究认为淡水河流或湖泊的挺水植物[如芦苇和宽叶香蒲(Typhalatifolium)]会对藻类产生化感作用,抑制其大量生长,但河口盐沼湿地植被是否也会对藻类产生明显的化感作用,还有待于研究。
但本研究也发现盐沼植被样地浮游植物丰度明显高于2004-2005年东滩中华鲟(Acipensersinensis)保护区水域[3]以及2001-2003年的杭州湾和舟山水域[1]。其中,优势种中肋骨条藻的优势度高出长江口南支水域和长江口口门区及邻近海域10倍以上[1-2, 14],而中肋骨条藻是盐沼湿地鱼类优势种(如鲻科、鲤科)和大型底栖动物的天然良好饵料[9, 16],因此可知盐沼生态系统中浮游植物群落构成与邻近水体有着较大的不同,具有独特的生态功能。
3.2 浮游植物群落组成的季节变化特征
众所周知,中纬度河口区浮游植物群落组成具有明显的季节变化和月变化[25-27],一般来说长江口浮游植物夏季种类数和丰度最高,其次是春季或秋季,冬季较低[1, 14]。本研究发现,5-8月(春末至夏末)浮游植物种类数较高,而9-11月(秋季)种类数下降明显,12月(冬初)上升至12种;从平均丰度来看,夏季最高,其中7月丰度最高,高达8.96×104cell· L-1,其次是秋季,而冬初(12月)丰度相比11月下降了约95%,主要是中肋骨条藻数量急剧下降导致。各个植被样点的浮游植物丰度变化受水温影响较大,而受潮高的影响较小。
各个月份4个植被样地浮游植物的种类数没有显著性差异,但HS和HZ植被样地平均丰度明显高于HH和LW植被样地,反映了高程会导致不同盐沼植被样地浮游植物的分布差异,因HH和LW植被样地高程相对较高导致潮水较难进入,水体与滩涂的摩擦、悬浮物絮集作用等原因,导致细胞数量减少。但在一些月份(如10-12月)高程较高的HH和LW植被样地浮游植物丰度却高于HS和HZ植被样地,因此可知芦苇和互花米草入侵后期阶段植被样地浮游植物同样具有较高的丰富度。由于HH和LW植被样地更靠近大堤处,夏季丰水期(7-8月)因崇明岛水闸的排水等原因会将岛内河道的淡水排到监测区域,造成该时期HH和LW植被样地蓝、绿藻丰度是高程较低的HS和HZ植被样地的5.5倍(图2)。
互花米草作为一种外来入侵种,1995年以来逐渐入侵崇明东滩和长江口北支,由于其强适应性、扩散能力和竞争力,严重威胁着崇明东滩生物多样性保护与生态安全,侵占了大部分的土著植被芦苇和海三棱藨草原有生态位[15]。本研究发现,互花米草入侵早期(HZ)和后期阶段(HH)植被样地浮游植物月平均值略低于同一高程的海三棱藨草和芦苇植被样地,但差异并不显著,可能因互花米草覆盖度较高、植株更密导致浮游植物丰度相对较低[15],但由于水体是流动的,互花米草入侵并没有显著降低水体浮游植物的种类数和丰度。
3.3 多样性和群落结构
对河口区浮游植物群落的月动态仅有极少的研究[14],而对于不同盐沼植被样地内浮游植物群落的研究则未见报道。长江口南支碎波带水域浮游植物5-12月Shannon-Weaver指数(H′)变化范围为1.61~2.26(平均值为1.84)[14],而本研究发现盐沼植被区浮游植物H′指数变化范围为0.20~1.60(平均值为0.85),物种丰富度指数(D)和Pielous种类均匀度指数(J)同样低于南支碎波带水域,这可能由于采样点位于长江口北支东北部水域(盐度平均值比南支高约10.5)和优势种中肋骨条藻优势度过高等原因导致浮游植物群落多样性指数偏低。从浮游植物群落聚类和排序结果来看,从空间上4种植被样地难以区分开,而从时间上大致可以分为2组[即5-8月(春初至夏末)与9-12月(秋季至冬初)];这是因为春夏季随着温度的快速升高,浮游植物大量生长使其丰度较高,而进入秋季至冬初因温度逐渐下降,且浮游植物生长能力下降使其丰度降低,造成9-12月4个植被样地间浮游植物群落差异很小。引起浮游植物群落A 、B组差异贡献率最大的物种是中肋骨条藻,其次是舟形藻属、美丽斜纹藻、近缘斜纹藻、洛氏菱形藻和新月菱形藻等。
参考文献:
[1] 王云龙, 袁 骐, 沈新强. 长江口及邻近水域春季浮游植物的生态特征[J]. 中国水产科学, 2005, 12(3): 300-306.
WANG Y L, YUAN Q, SHEN X Q. Ecological character of phytoplankton in spring in the Yangtze River estuary and adjacent waters [J]. Journal of Fishery Sciences of China, 2005,12(3): 300-306.
[2] 顾新根, 袁 骐, 杨蕉文, 等. 长江口羽状锋海区浮游植物的生态研究[J]. 中国水产科学, 1995, 2(1): 1-15.
GU X G, YUAN Q, YANG J W,etal. An ecological study on phytoplankton in frontal region of Changjiang estuary area [J]. Journal of Fishery Sciences of China, 1995, 2(1): 1-15.
[3] 王云龙, 袁 骐, 庄 平,等. 长江口中华鲟自然保护区附近水域浮游植物分布[J]. 长江流域资源与环境, 2008, 17(1): 72-77.
WANG Y L, YUAN Q, ZHUANG P,etal. Distribution of Phytoplankton Around Yangtze Esturine Nature Reserve For Chinese Sturgeon[J]. Resources and Environment of Yangtze Catchments, 2008, 17(1): 72-77.
[4] 吴 琼, 吴 波, 赵爱萍, 等. 长江口九段沙附近水体浮游植物的种类组成与数量分布[J]. 上海师范大学学报(自然科学版), 2007, 36(1): 54-59.
WU Q, WU B, ZHAO A P,etal. Species composition and quantitative distribution of phytoplankton near the Jiu duansha waters of Yangtze River estuary [J]. Journal of Shanghai Normal University (Natural Sciences), 2007, 36(1): 54-59.
[5] 徐兆礼, 白雪梅, 袁 骐. 长江口浮游植物的生态研究[J]. 中国水产科学, 1999, 5(5): 52-54.
XU Z L, BAI X M, YUAN Q. An ecological study on phytoplankton in the Yangtze River estuary [J]. Journal of Fishery Sciences of China, 1999, 5(5): 52-54.
[6] 张锦平, 夏 静. 长江口浮游植物群落的聚类分析[J]. 上海水产大学学报, 2006, 16(5): 456-460.
ZHANG J P, XIA J. Cluster analysis of phytoplankton communities in the Yangtze River estuary during summer [J]. Journal of Shanghai Fisheries University, 2006, 16(5): 456-460.
[7] 顾新根, 袁 骐, 沈焕庭,等. 长江口最大浑浊带浮游植物的生态研究[J]. 1995, 2(1):16-27.
GU X G, YUAN Q, SHEN H T,etal. The ecological study on phytoplankton in maximum turbid zone of Changjiang estuary [J]. Journal of Fishery Sciences of China, 1995, 2(1):16-27.
[8] ZHANG H. Diel, semi-lunar and seasonal patterns in the fish community of an intertidal zone of the Yangtze Estuary [J]. Journal of Applied Ichthyology, 2013, 29(6): 1252-1258.
[9] 张 衡,朱国平. 长江河口潮间带鱼类群落的时空变化[J]. 应用生态学报, 2009, 20(10): 2519-2526.
ZHANG H, ZHU G P. Spatio-temporal changes of fish community in the Yangtze Estuary intertidal zone [J]. Chinese Journal of Applied Ecology, 2009, 20(10): 2519-2526.
[10] 钟俊生, 俞蔚文, 刘必林, 等. 长江口沿岸碎波带仔稚鱼种类组成和季节性变化[J]. 上海水产大学学报,2005, 14(4): 375-382.
ZHONG J S, YU W W, LIU B L,etal. Seasonal occurrences of fish larvae and juveniles in the surf zone of the Yangtze River estuary[J]Journal of Shanghai Fisheries University, 2005, 14(4): 375-382.
[11] 张 衡, 全为民, 陈渊戈, 等. 长江口口门区潮下带水域鱼类群落组成的季节变化[J]. 长江流域资源与环境, 2014, 23(11): 1535-1540.
ZHANG H, QUAN W M, CHEN Y Q,etal. Seasonal changes of fish community composition in the subtidal water in the mouth area of Yangtze estuary [J]. Resources and Environment in the Yangtze Basin, 2014, 23(11): 1535-1540.
[12] 张 衡, 何文珊, 童春富, 等. 崇西湿地冬季潮滩鱼类群落结构分析[J].长江流域资源与环境, 2007, 16(3): 308-313.
ZHANG H, HE W S, TONG C F,etal. An analysis on fish community structure in winter in Chong-xi wetland [J]. Resources and Environment of Yangtze Catchments, 2007, 16(3): 308-313.
[13] 童春富. 长江河口潮间带盐沼植被分布区及邻近光滩鱼类组成特征[J]. 生态学报, 2012, 32(20): 6501-6510.
TONG C F. Characteristics of the fish assemblages in the intertidal salt marsh zone and adjacent mudflat in the Yangtze estuary [J]. Acta Ecologica Sinica, 2012, 32(20): 6501-6510.
[14] 张松艳,王 琼,薛俊增,等. 长江口碎波带浮游植物群落结构特征[J]. 生物学杂志, 2014, 31(3): 15-19.
ZHANG S Y, WANG Q, XUE J Z,etal. Ecological features of phytoplankton community in the surf zones of the Yangtze River estuary[J]. Journal of Biology, 2014, 31(3): 15-19.
[15] 王睿照. 互花米草入侵对崇明东滩盐沼底栖动物群落的影响[D]. 上海:华东师范大学, 2010.
WANG R Z. Impacts ofSpartinaalterniflorainvasion on the benthic communities of saltmarshes in Chongming Dongtan, Shanghai [D]. Shanghai:East China Normal University, 2010.
[16] 张 衡,叶锦玉,梁晓莉,等.长江口东滩湿地芦苇生境大型底栖无脊椎动物群落结构的月动态[J].应用生态学报,2017, 28(4): 1360-1369.
ZHANG H, YE J Y, LIANG X L,etal. Monthly changes in the benthic macro-invertebrates community structure in the habitats ofPhragmitesaustralismarsh in the Dongtan wetland of the Yangtze River estuary, China [J]. Chinese Journal of Applied Ecology, 2017, 28(4): 1360-1369.
[17] 孙 军, 刘东艳, 钱树本. 一种海洋浮游植物定量研究分析方法Utermöhl方法的介绍及其改进[J]. 黄渤海海洋, 2002, 20(2): 105-112.
SUN J, LIU D Y, QIAN S B. A quantitative research and analysis method for marine phytop lankton: An introduction to Utermöhl method and its modification[J]. Journal of Oceanography of Huanghai & Bohai Seas, 2002, 20(2): 105-112.
[18] 钱迎倩, 马克平. 生物多样性研究的原理与方法[M]. 北京: 中国科学技术出版社, 1994.
QIAN Y Q, MA K P. Principles and methods of biodiversity research [M]. Beijing: China Science and Technology Publishing Press,1994.
[19] CLARKE K R, WARWICK R M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness [J]. Marine Ecology Progress Series, 2001a(216): 265-278.
[20] CLARKE K R, WARWICK R M. Change in marine communities: An approach to statistical analysis and interpretation (2nd edition)[M]. Plymouth: PRIMPER-E, 2001b.
[21] 赵 冉, 白 洁, 孙 军,等. 2006 年夏季长江口及其邻近水域浮游植物群集[J]. 海洋湖沼通报, 2009(2): 88-96.
ZHAO R, BAI J, SUN J,etal. Phytoplankton assemblages in the Yangtze River estuary and its adjacent water in summer, 2006[J]. Transactions of Oceanology and Limnology, 2009(2): 88-96.
[22] 王金辉. 长江口3 个不同生态系的浮游植物群落[J]. 青岛海洋大学学报, 2002, 32(3):422-428.
WANG J H. Phytoplankton Communities in Three Distinct Ecotypes of the Changjiang Estuary [J]. Journal of Ocean University of Qingdao, 2002, 32(3): 422-428.
[23] 林 军,朱建荣, 张 经,等. 长江口外海区浮游植物生物量分布及其与环境因子的关系[J]. 水产学报,2011, 35(1): 74-87.
LIN J, ZHU J R, ZHANG J,etal. Distribution of phytoplankton biomass and its relationships with the environmental factors off the Changjiang (Yangtze River) Estuary [J]. Journal of Fisheries of China, 2011, 35(1): 74-87.
[24] 鲜啟鸣, 陈海东, 邹惠仙, 等. 淡水水生植物化感作用研究进展[J]. 生态学杂志, 2005 , 24(6): 664-669.
XIAN Q M, CHEN H D, ZOU H X,etal. Research advances in allelopathy of aquatic macrophytes [J]. Chinese Journal of Ecology, 2005, 24(6): 664-669.
[25] 栾青杉, 孙 军. 2005 年秋季长江口及其邻近水域浮游植物群集[J]. 长江流域资源与环境, 2010, 19(2): 202-208.
LUAN Q S, SUN J. Phytoplankton assemblage of the Yangtze River estuary and its adjacent waters in autumn, 2005[J]. Resources and Environment of Yangtze Catchments, 2010, 19(2): 202-208.
[26] MATOS J B, SILVA N I S, PEREIRA L C C,etal. Phytoplankton of the surf zone in Amazon beach [J]. Acta Botanica Brasilica, 2012(26): 979-990.
[27] KAHN A E, CAHOON L B. Phytoplankton productivity and photophysiology in the surf zone of sandy beaches in North Carolina [J]. Estuaries and Coasts, 2012, 35(6): 1393-1400.
猜你喜欢
法人(2022年3期)2022-03-30
海洋通报(2021年1期)2021-07-23
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
海洋通报(2020年2期)2020-09-04
家教世界·V家长(2020年6期)2020-07-14
——乌尤尼盐沼
家教世界(2020年17期)2020-07-06
流行色(2019年10期)2019-12-06
环球时报(2019-11-14)2019-11-14
百科探秘·航空航天(2017年10期)2017-11-08
