
外源抗氧化剂α-生育酚在河蟹肌肉脂肪-蛋白质氧化体系中的作用
2018-07-24 08:44刘小莉彭欢欢夏秀东周剑忠赵江涛
食品科学 2018年14期
刘小莉,彭欢欢,,夏秀东,周剑忠,刘 源,赵江涛,*
(1.江苏省农业科学院农产品加工研究所,江苏 南京 210014;2.上海海洋大学食品学院,上海 201306)
淡水水产品肉质细嫩,营养丰富,不易保鲜和贮藏。冻藏是保存水产品的有效方法,但也不可避免地造成品质下降。这是由于水产品中含有大量高度不饱和脂肪酸和蛋白质,即使在低温条件下也会氧化变质[1]。越来越多的研究表明,脂肪在氧化的过程中会产生大量的自由基和活性次生氧化产物,进一步与蛋白质形成共价结合,从而诱导蛋白质聚合反应[2-3]。一方面,脂肪过氧化初级产物裂解产生脂质自由基,自由基通过氢反应充当印发剂使蛋白质分子变成自由基,蛋白质自由基又引发聚合式链反应最终导致蛋白质聚合[4]。另外,脂质氧化的次生产物,即醛类或酮类化合物的羰基能够与蛋白分子中的氨基侧链基团反应,使多肽链发生链内或链间交联现象[5]。蛋白质进一步通过氢键、疏水键和二硫键等的作用进行重排,形成高分子的蛋白质聚合物,从而改变水产品的组织、颜色、水合能力和风味等一系列营养特性。
Laurizsen等[6]研究表明鱼类产品在冷冻贮存过程中脂肪会发生氧化,氧化产物使鱼肉质构发生变化,如组织蛋白变硬、聚集。此外,脂肪氧化还会使部分氨基酸,如组氨酸、赖氨酸、蛋氨酸和半胱氨酸损失,并损害其他色素蛋白质,如细胞色素C和血红蛋白。Park等[7-8]比较了3 种模拟体系中猪肉肌原纤维蛋白的变化,发现在不同的氧化体系条件下肌原纤维蛋白发生不同程度的变化,主要表现为羰基的形成,蛋白热稳定性下降以及蛋白交联聚集等。Faustman等[9]发现脂肪氧化的初级产物和次级产物对于蛋白质氧化起促进作用,脂溶性抗氧化剂α-生育酚能够有效抑制脂肪氧化,亦可有效抑制水溶性蛋白的氧化。
河蟹是我国传统特色的经济蟹类,但上市时间比较集中,大部分以鲜活河蟹进行销售,很容易造成供大于求而滞销的局面,特别是小规格蟹价格低,严重影响了河蟹产业经济效益的提高[10]。因此有必要进行河蟹精深加工的研究,其中原料的贮藏问题是河蟹加工产业的瓶颈问题。然而目前对冻藏过程中河蟹肌肉脂肪氧化导致蛋白质变性方面的研究鲜见报道。α-生育酚是一种天然抗氧化物质,无毒性,目前广泛应用于多种食品的抗氧化研究[11]。本实验建立河蟹氧化脂肪-蛋白质的模拟体系,探讨氧化脂肪所引起的蛋白质特性的改变,以及α-生育酚对氧化脂肪诱导蛋白质发生变性的抑制作用,并以此为指导,控制脂肪氧化而减少蛋白质变性,为提高食品品质提供理论依据。
1 材料与方法
1.1 材料与试剂
鲜活公河蟹由常熟市金唐市水产有限公司提供,体质量(150±15)g,江苏省淡水水产研究所鉴定为中华绒螯蟹(Eriocheir sinensis H. Milne-Edwards)。
α-生育酚、抗坏血酸(食品级) 郑州思源食品添加剂有限公司;5,5’-二硫代双-硝基苯甲酸 美国Sigma公司;BCA法蛋白定量测试盒、丙二醛测定试剂盒 南京建成生物工程研究所;过硫酸铵(ammonium persulphate,APS)、N,N,N’,N’-四甲基乙二胺(tetramethylethylenediamine,TEMED)、预混合分离胶、预混合浓缩胶、5×十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)蛋白上样缓冲液、快速蛋白染色液 新赛美生物科技有限公司;石油醚、氢氧化钾、甲醇、三氟化硼、乙醚、氯化钠、正己烷、磷酸氢二钠、磷酸二氢钠、三羟甲基氨基甲烷(2-amino-2-(hydroxymethyl)-1,3-propanediol,Tris)、氯化钾、溴酚蓝均为国产分析纯。
1.2 仪器与设备
VK-6001绞肉机 德国欧诺华公司;RE-6000旋转蒸发仪 上海亚荣生化仪器厂;T25匀质分散机 德国IKA公司;TSQ 800 Evo气相色谱-质谱联用仪 赛默飞世尔科技公司;DHG-9146A电热恒温鼓风干燥箱 上海精宏实验设备有限公司;3K15高速离心机 北京五洲东方科技发展有限公司;UV-1600PC紫外分光光度计上海美谱达仪器有限公司;SynergyH1酶标仪 美国柏腾仪器有限公司;GS-800垂直电泳槽 美国Bio-Rad公司;150型河蟹采肉机 山东省诸城市兴和机械有限公司。
1.3 方法
1.3.1 蟹肉采集及预处理
采用河蟹采肉机,通过挤压方式采集蟹肉,-20 ℃保存备用。
1.3.2 脂质的提取及氧化指标测定
采用Folch等[1]的方法提取蟹油。制备的蟹肉用20 倍体积的石油醚(沸程60~90 ℃)混合,室温振摇40~60 min,收集有机溶剂相。下层水相样品再加入有机溶剂提取2 次。合并有机相,40 ℃旋转蒸发挥干有机溶剂,得到粗蟹油,待用。按照GB/T 5009.37—2003《食用植物油卫生标准的分析方法》中第二法所述测定过氧化值。依据硫代巴比妥酸法原理采用试剂盒检测丙二醛含量,结果以1 mg样品蛋白质中丙二醛含量(nmol/mg)表示。
1.3.3 气相色谱-质谱联用分析脂肪酸组成
参照Liu Wei等[13]的方法对油脂进行甲酯化处理。取所得蟹油0.2 mL,加入2 mL体积分数为5%的氢氧化钾-甲醇溶液,75 ℃水浴15 min,冷却后加入2 mL体积分数为14%的三氟化硼乙醚-甲醇溶液,75 ℃水浴2 min,加入饱和氯化钠溶液2 mL和正己烷1 mL,混匀后静置,取上清液,采用气相色谱-质谱联用法检测蟹油脂肪酸组成。
色谱条件:SLB-5MS毛细管色谱柱(30 m×0.25 mm,0.25 µm);升温程序:120 ℃保持6 min,以3 ℃/min升温到250 ℃,保持25 min;载气(He)流速1.2 mL/min,压力2.4 kPa,进样量1.0 µL;分流比30∶1。
质谱条件:电子电离源;电子能量70 eV;灯丝发射电流200 μA;离子源温度200 ℃;接口温度250 ℃;检测器电压350 V;质量扫描范围m/z 33~800。
1.3.4 模拟反应体系建立
将提取的油脂于90 ℃烘箱中分别放置12 h和24 h,得到氧化型蟹油。取未被氧化、氧化12 h、氧化24 h的蟹油各10 mL,分别加入到100 g蟹肉中,用匀浆机搅拌均匀,得到3 组蟹油-蛋白质模拟体系(编号I、II、III)。将上述3 组模拟体系各分为2 份,其中一组分别加入0.015%的α-生育酚和0.02%的抗坏血酸(编号IV、V、VI)。共6 组处理进行实验。将6 组样品置于常温条件下1周,定期取样进行蛋白质氧化指标的测定。
1.3.5 肌原纤维蛋白的提取
参照Fang Yang等[14]的方法进行不同处理的样品中肌原纤维蛋白的提取。称取5 g蟹肉样品加10 mL、4 ℃预冷的去离子水,12 000 r/min匀浆30 s,10 000 r/min、4 ℃离心20 min,弃去上清液,沉淀中加入去离子水,再重复提取1 次。沉淀中再加入20 mL、4 ℃预冷的0.05 mol/L磷酸缓冲液(pH 7.2)(其中补充0.6 mol/L的NaCl),12 000 r/min匀浆30 s,10 000 r/min、4 ℃离心20 min,收集上清液。沉淀用上述步骤再重复提取1 次,合并上清即为肌原纤维蛋白粗提取液,其浓度采用BCA试剂盒测定。
1.3.6 活性巯基的测定
参照Benjakul等[2]的方法进行测定。1.3.5节中提取的蛋白溶液0.5 mL,加入4.5 mL 0.2 mol/L Tris-HCl缓冲液(pH 6.8)。取该混合液1 mL,加入0.1 mL 0.1% 5,5’-二硫代双-硝基苯甲酸,40 ℃温育25 min,测定波长412 nm处的吸光度。空白样用0.6 mol/L KCl(pH 7.0)代替样品。活性巯基含量以1 mg蛋白质中活性巯基的物质的量计,按公式(1)计算:
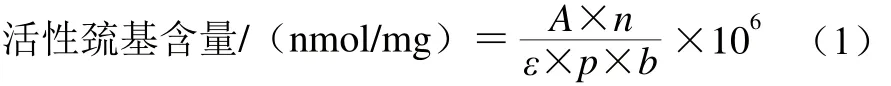
式中:A为412 nm波长处的吸光度;n为稀释倍数;ε为摩尔吸光系数13 600/(L/(mol·cm));p为蛋白质质量浓度/(mg/mL);b为吸光池光程(1 cm)。
1.3.7 表面疏水性的测定
参考Chelh等[3]的方法,并稍作修改。将提取的肌原纤维蛋白溶液质量浓度调整为1 mg/mL,取1 mL加入40 μL1 mg/mL的溴酚蓝溶液,空白对照为1 mL提取缓冲液加入40 μL1 mg/mL的溴酚蓝溶液,涡旋振荡混匀10 min,于4 ℃、4 000×g离心15 min,取上清液稀释10 倍后在波长595 nm处测定吸光度。表面疏水性按公式(2)计算:

式中:S为表面疏水性/μg;Acontrol为空白的吸光度;Asample为样品的吸光度;40为系数。
1.3.8 SDS-PAGE检测
取10 mL的8%预混合分离胶加入100 μL 10% APS和5 μL TEMED,混匀后灌入制胶板内,加入适量的醇或水压平分离胶,待20~30 min后凝聚成分离胶。取5 mL的预混合浓缩胶加入50 μL 10% APS和5 μL TEMED,混匀后灌入制胶板内,缓慢插入梳子,待20~30 min后凝聚即为浓缩胶。取12 μL的蛋白上样液进行电泳,全程150 V,待溴酚蓝到达底部,即可完成电泳。结束后,用去离子水洗涤3 次,每次约5 min,再加入约20 mL快速蛋白染色液过夜,用去离子水脱色1 h后成像拍照。
1.3.9 傅里叶变换红外光谱的测定
将提取的肌原纤维蛋白溶液放入真空冷冻干燥机中干燥得到粉末。将样品粉末均匀地铺满在采样器上,进行红外光谱扫描,扫描次数256,分辨率为4 cm-1。
1.4 数据处理
本实验数据为3 次重复的平均值。采用SPSS 13.0软件进行数据统计和方差显著性分析。
2 结果与分析
2.1 蟹油氧化指标及脂肪酸组成
过氧化值和丙二醛是常用的脂质氧化评价指标,本研究中新鲜蟹油的过氧化值和丙二醛含量分别为(3.61±0.26) meq/kg、(11.64±1.26) nmol/mg,氧化12、24 h后过氧化值分别达到(5.50±0.47) meq/kg和(8.26±0.21) meq/kg,丙二醛含量分别达到(16.51±0.87) nmol/mg和(29.86±0.73) nmol/mg,可见在高温条件下,蟹油快速发生氧化,导致酸败。由表1可以看出,新鲜的蟹油饱和脂肪酸相对含量为15.21%,氧化12、24 h后分别达到29.83%、44.85%。蟹油脂肪酸中油酸相对含量最高,高温氧化后,油酸相对含量显著下降,分别仅为57.01%、43.87%。由于油脂在氧化过程中,自由基会连锁攻击不饱和脂肪酸,从而使得不饱和脂肪酸发生衰败,蟹油中不饱和脂肪酸总量会随氧化时间的延长呈下降趋势,相应饱和脂肪酸的含量升高。其他学者也报道过类似的研究结果[17]。研究表明,脂肪氧化与蛋白质氧化之间是相互关联的,且两者中的任一种物质氧化产生的化合物都会促进另一种物质的氧化[18-19]。脂肪氧化形成的羟自由基可以夺取蛋白质分子的氢离子,使得蛋白质发生与脂肪氧化类似的自由基链式反应[20-21]。
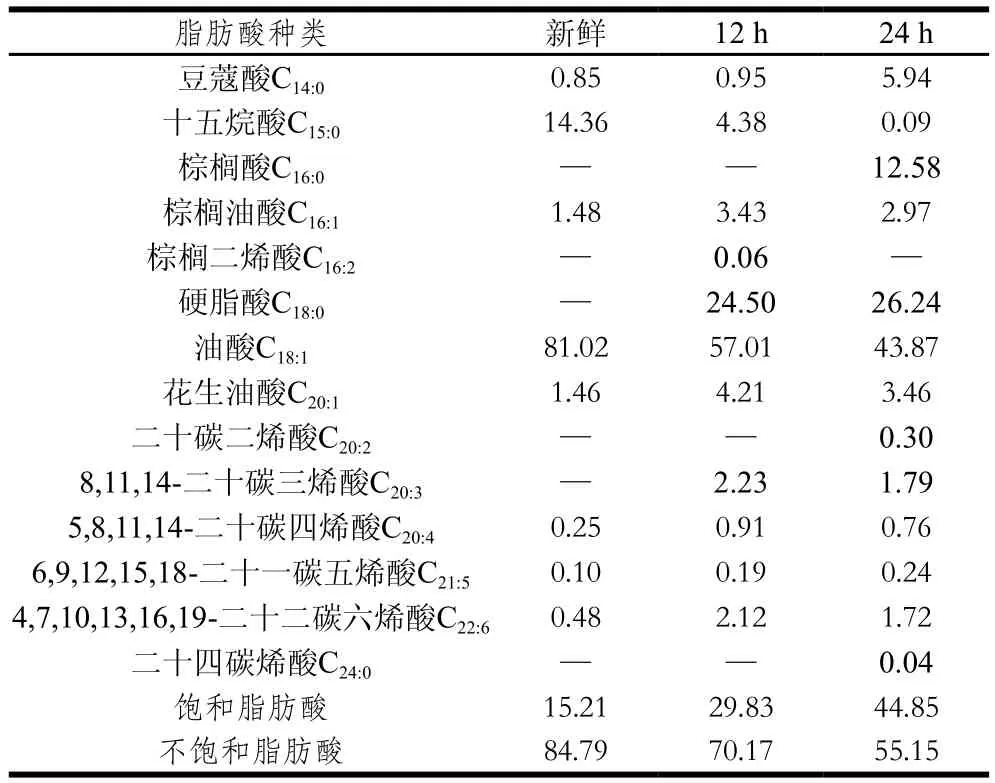
表1 新鲜与氧化蟹油中脂肪酸相对含量Table1 Contents of fatty acids in fresh and oxidized crab oils%
2.2 活性巯基含量的变化
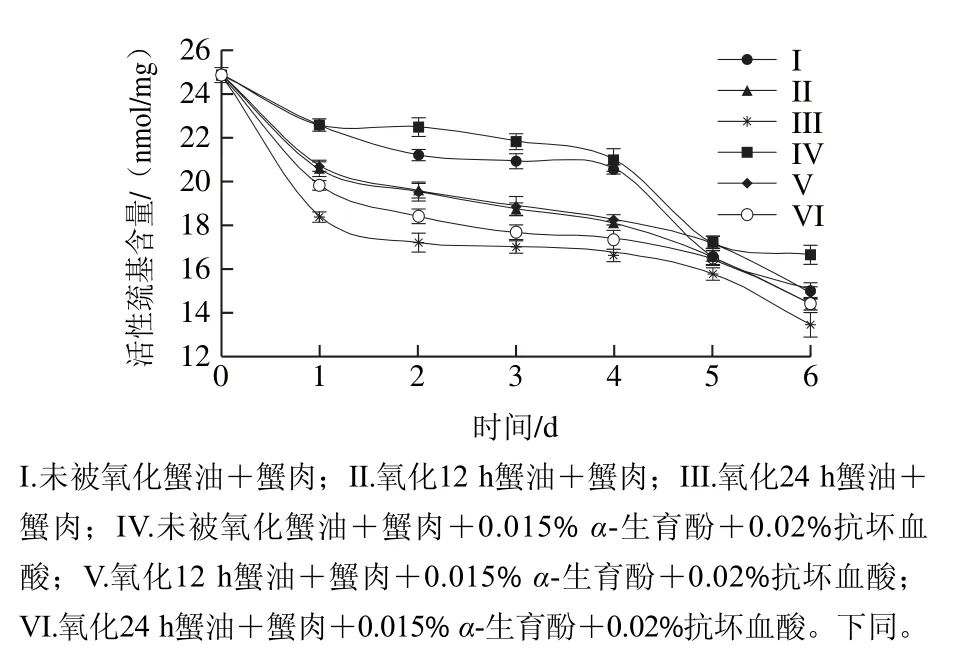
图1 肌原纤维蛋白中巯基含量的变化Fig.1 Changes in active sulfhydryl content of myofibrillar protein
活性巯基含量的变化能够反映出蛋白质变性集合的程度,活性巯基含量下降的主要原因是巯基氧化形成二硫键所致[22]。由图1可以看出,6 组样品在贮藏期间活性巯基含量都不断减少。未加抗氧化剂的3 组对比发现,添加氧化蟹油,下降速度越快,这可能是经人工氧化的蟹油产生大量的自由基,进而诱导肌原纤维蛋白分子发生变性和蛋白质结构改变,而使得隐藏的分子内部的巯基活性基团暴露出来,巯基易于氧化形成二硫键,导致活性巯基含量的不断下降。添加抗氧化剂的活性巯基减少量低于未加抗氧化剂组,由于α-生育酚属于酚类物质使得羟基释放活泼氢,与自由基的结合会抑制其对脂质的攻击。由此可以看出,α-生育酚对巯基具有一定的保护作用,从而抑制了蛋白质分子的交联、聚合现象的发生。
2.3 表面疏水性的变化
脂质氧化还可能引起蛋白质分子内部结构和性质发生改变。其中表面疏水性是与外界极性水环境相连的蛋白质表面疏水性基团数量的一个重要标志[23]。蛋白质在降解发生变性的过程中,隐藏在蛋白质内部的分子暴露出来,改变其疏水性,导致疏水性基团的增加,因此蛋白质表面疏水性可以在一定程度上衡量蛋白质的变性程度。当蛋白质分子空间结构发生变化时,分子内部的疏水基团和亲水基团相对位置也会发生改变,导致表面疏水性指数发生变化[24-25]。

图2 肌原纤维蛋白中表面疏水性的变化Fig.2 Changes in surface hydrophobicity of myofibrillar protein
由图2可以看出,在贮藏过程中,随着蟹油氧化程度的增加,肌原纤维蛋白的表面疏水性显著升高(P<0.05),可能是蛋白质分子折叠,使肽链断裂或结构伸展,分子的内部疏水基团暴露导致表面疏水性的增加。说明油脂的氧化在一定程度上可以促使肌原纤维蛋白疏水基团的暴露,这与章银良等[26]的研究结果一致。添加α-生育酚的样品,表面疏水性都较相应的未添加α-生育酚的对照样品模拟体系少。α-生育酚抗氧化作用的机制主要是因为α-生育酚可以首先替代其他的物质被氧化,从而延缓不饱和脂肪酸的氧化作用,且以抗坏血酸为增效剂,可以提高其抗氧化效能[27-28]。在贮藏初期,表面疏水性随着时间延长不断提高,从而诱导蛋白质表面疏水性不断增大,当反应到后期脂肪氧化趋于饱和。α-生育酚可以抑制微生物的生长,延缓蛋白质的降解作用及蛋白变性,进而延缓表面疏水性的增加;而且α-生育酚可能抑制了脂肪氧化体系对蛋白质折叠的作用,而延缓表面疏水性的增加。
2.4 SDS-PAGE结果

图3 河蟹肌肉肌原纤维蛋白的SDS-PAGE图谱Fig.3 SDS-PAGE profiles of myofibrillar protein
由图3可以看出,条带从上到下分别是肌球蛋白重链、肌动蛋白、原肌球蛋白和肌球蛋白轻链,其中肌球蛋白重链和肌动蛋白是主要的蛋白质条带[29]。6 组样品的肌原纤维蛋白条带随着贮藏时间延长,发生不同程度的变淡。贮藏1 d后蟹肉肌原纤维蛋白条带清晰,可以清楚的辨别蛋白中的肌球蛋白重链、肌动蛋白、原肌球蛋白和肌球蛋白轻链。蛋白质在贮藏3 d后,肌球蛋白重链条带明显减弱,且氧化程度越高,条带减弱程度越明显,添加抗氧化剂的3 组IV、V、VI较相对应的未添加抗氧化剂3 组I、II、III条带降解相对缓慢,但Gel Image System软件对条带光密度分析结果显示,除了贮藏1 d后的V、II组肌球蛋白重链、肌动蛋白存在显著差异,其他组间条带差异不显著。贮藏结束后,几乎所有蛋白质条带变浅消失。结合2.2节和2.3节结果,贮藏过程中不同处理组间活性巯基含量和表面疏水性指标差异显著,可见SDS-PAGE用于检测蛋白质氧化程度不及活性巯基和表面疏水性指标灵敏。
2.5 傅里叶变换红外光谱测定
将肌原纤维蛋白溶液冷冻干燥,利用傅里叶变换红外光谱仪进行光谱扫描,选择波数1 600~1 700 cm-1范围经Peakfit 4.12软件分析得到自动去卷积后的蛋白质二级结构。在蛋白质的二级结构中,α-螺旋结构是蛋白质分子的有序结构,具有高度的结构稳定性;而β-转角和无规卷曲为蛋白质分子的无序结构,因此可将α-螺旋结构的相对含量用于判断蛋白质结构的稳定性[30-31]。如表2所示,蟹肉与氧化12 h的蟹油混合,贮藏期结束后蛋白质β-转角相对含量显著增加,其他二级结构没有显著差异;氧化24 h后的蟹油则对蟹肉中蛋白质二级结构影响显著,与对照相比,除了无序结构外其他结构均有显著变化,α-螺旋和β-折叠相对含量显著减少,β-转角相对含量显著增加。本研究中添加0.015% α-生育酚和0.02%抗坏血酸的抗氧化剂处理组,很好地维持了蟹肉中蛋白质的二级结构稳定,与对照相比只有氧化24 h蟹油组的β-转角相对含量显著增加,其他二级结构无显著差异,说明添加抗氧化剂能很好地维持蟹肉蛋白质的二级结构稳定。

表2 贮藏结束后不同样品中蛋白质二级结构相对含量Table2 Protein secondary structures in diff erent samples after storage%
3 结 论
河蟹肌肉蟹油经过高温氧化后,脂肪酸组成发生变化,不饱和脂肪酸相对含量明显下降。在氧化蟹油的作用下,河蟹肌肉蛋白质中活性巯基含量下降,表面疏水性上升,且油脂氧化程度越高,所引起的蛋白质变性越显著。添加抗氧化剂的处理组对巯基具有保护作用,且减少表面疏水值的增加;SDS-PAGE结果显示,随着贮藏时间的延长,氧化程度越高,条带减弱程度越明显,添加抗氧化剂的3 组IV、V、VI较相对应的未添加抗氧化剂3 组I、II、III条带降解相对缓慢,但效果并不显著,说明SDS-PAGE法用于判定蛋白质氧化不及活性巯基和表面疏水性指标灵敏。傅里叶变换红外光谱结果显示,油脂氧化程度越高,相应的处理组蛋白质二级结构中α-螺旋、β-折叠相对含量越少,β-转角则越多,添加抗氧化剂能维持蟹肉蛋白质相对稳定的二级结构。
猜你喜欢
发明与创新(2022年27期)2022-09-16
广州化学(2022年4期)2022-09-01
科学养鱼(2022年12期)2022-02-04
工业水处理(2020年10期)2020-10-22
食品科学(2020年11期)2020-07-13
小资CHIC!ELEGANCE(2016年7期)2016-05-14
海洋科学进展(2015年1期)2015-02-27
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年11期)2014-03-11
