
红缨海棠嫩枝扦插生根解剖学观察
2018-07-27 08:06史锋厚沈永宝
经济林研究 2018年3期
史锋厚,赵 瑞,罗 帅,沈永宝
(南京林业大学 a.林学院;b.南方现代林业协同创新中心,江苏 南京 210037)
扦插作为林木无性繁殖的重要方式,已被广泛应用于林木种苗生产。影响插穗生根的因素较多,既包括树种的遗传因素和插穗的生理状态,又包括扦插环境。一般插穗的不定根原始体按其形成时间分为潜伏根原始体和诱生根原始体两种[1]。林木扦插生根难易和生根时间因树种(品种)而异,源于枝条内是否存在潜伏根原始体和根原基诱导的难易,一般而言,存在潜伏根原始体的树木扦插较易生根[2],诱导根原基发生部位较多的树种较易生根[1]。植物激素或生长调节剂可以促进插穗生根,尤其对于一些难生根树种效果明显,它不仅可以提高生根率、缩短生根时间,还可以提高生根质量,如经600 mg/L NAA溶液处理的绣球插穗生根率可达94%[3],外施6 000 mg/L K-IBA可显著提高北美香柏插穗的生根率及生根质量[4],经500 mg/L IBA溶液浸泡处理的钩栗插穗生根率可达99.7%[5]。植物激素或生长调节剂促进插穗生根的原因由解剖学分析与其对根原基的诱导有关。
红缨海棠于2017年3月10日被江苏省林木品种审定委员会审定为良种,可采用扦插方式繁殖[6]。本文将以红缨海棠嫩枝插穗作为研究材料,使用NAA对插穗进行促根处理后扦插于全光照自动喷雾扦插池中,定期取样插穗基部制作石蜡切片,观察插穗根原基诱导发生部位及不定根发育过程。
1 材料与方法
1.1 嫩枝扦插及管理
2017年7月底,红缨海棠采穗圃采集半木质化枝条制作100根插穗。插穗规格为长度15 cm左右,保留2~3个芽,插穗上部保留2片半叶,上下切口平截。80根插穗基部使用1 000 mg·L-1NAA溶液浸蘸5 s后,按照10 cm×5 cm间距插入以蛭石为基质的全光照间歇喷雾扦插池中,扦插深度为插条长度的1/2;其余20根插穗以清水浸泡作为对照,插于扦插池中。
扦插池每隔20 min喷雾1次,每次喷15 s,阴天酌量减少喷雾频次和时间。扦插后每隔7 d,使用50%多菌灵可湿性粉剂800倍液对插穗和基质喷雾消毒1次。
1.2 取样、固定、制作切片
扦插后,每隔2 d取样1次,共取15次;取样的同时,对插穗外部形态进行观察。每次取3根插穗,将插穗基部1 cm左右切成小段放于盛有70% FAA固定液的玻璃小瓶中固定。样品固定24 h后,转移至甘油和95%乙醇(体积比为1∶1)混合液中,软化处理60 d后取出,制作石蜡切片。
参照侯福林[7]的方法,样品经脱水、透明、渗蜡后即可包埋。使用石蜡切片机切片,切片厚10 μm,用番红固绿染色,二甲苯透明,中性胶封藏,制成永久切片,用OLYMPUS显微镜观察拍照。
2 结果与分析
2.1 插穗生根过程中的外部形态观察
扦插4 d后,大多数插穗下切口处未产生愈伤组织,极少数插穗切面髓部出现少量愈伤组织突起,呈晶莹透明状。扦插8 d后,插穗下切口部位产生肿胀并伴有大量的白色突起。扦插12 d后,部分插穗中下部皮部突起或皮孔出现白点,皮孔开裂,对照处理插穗仍未产生愈伤组织;此外,部分插穗下切口处木质部出现不同程度的腐烂状况。扦插16 d后,少量对照插穗产生愈伤组织,大部分NAA处理插穗产生愈伤组织且体积较大;同时,还发现部分插穗未产生不定根也无愈伤组织,多数此类型插穗下切口处出现腐烂。扦插18 d后,插穗分生出大量不定根,不定根根长超过5 cm,具有高位不定根的插穗基部腐烂较为严重。扦插20 d后,约半数插穗生根。扦插22 d后,大部分插穗生根且不定根生长较长。见图1。综合插穗外部形态观察,插穗愈伤组织部位和皮部均可产生不定根,其中愈伤组织部位生根较多,判定NAA处理后的红缨海棠嫩枝扦插生根类型为以愈伤组织生根为主型[6]。

图1 插穗生根过程中的外部形态观察Fig. 1 The external morphology observation of rooting
2.2 扦插前插穗解剖构造

图2 扦插前插穗解剖构造Fig. 2 The anatomical structure of stem before cutting
扦插前插穗横切面层次分明,由外向内依次为表皮、皮层、韧皮部、维管形成层、木质部、髓,见图2。由图2可知,表皮(E)细胞为规则的长方形细胞且排列紧密,染色较深。表皮里面是皮层(C),细胞为不规则的卵石形,排列较紧密。次生韧皮部(P)主要由韧皮薄壁细胞、韧皮纤维及筛管组成,与木质部相比,其只占很小的部分。维管形成层(VC)由较小且排列紧密的若干层分生细胞组成。次生木质部(X)所占比例较大,主要由导管和木纤维组成,细胞木质化程度较高。插穗的中央部分是髓(PI),原生木质部在髓与次生木质部之间,其染色相比次生木质部较深。髓部细胞较大,排列疏松,染色较浅。在髓与木质部结合处存在一条染色较深的波浪状环带,其细胞与髓部细胞相比,体积小且排列紧凑,为环髓带(PZ)。狭窄的髓射线(PR)从环髓带扩展到韧皮部,细胞染色较深。通过石蜡切片观察,扦插前插穗中不存在根原始体,证实红缨海棠枝条内不存在潜伏根原基。
2.3 不定根诱导发生的部位及发育过程
红缨海棠嫩枝扦插后,可由插穗愈伤组织和皮部长出不定根,即插穗愈伤组织及皮部均可诱导产生根原基,各处根原基诱导及不定根发育过程如下:
2.3.1 愈伤组织内分化的根原基
愈伤组织生根是树木嫩枝扦插生根的主要类型之一,通过石蜡切片观察,红缨海棠嫩枝扦插主要的生根部位就是愈伤组织。
愈伤组织是扦插后插穗基部的形成层恢复分裂能力后产生的一种组织,由维管形成层发育而来,由薄壁细胞组成[8]。薄壁细胞个大、近圆形、排列疏松,细胞核不明显。愈伤组织在细胞数量不断增加的同时其内部的一些细胞开始分化,伴随着细胞壁的增厚一些细胞分化成输导组织。愈伤组织内的输导组织一部分是插穗基部维管形成层分裂分化的延续(图3A),另一部分独立形成于愈伤组织内部(图3C)。
红缨海棠愈伤组织中可分化形成木质部和形成层。在木质部外侧,有些细胞特化形成具有根原始体特征的细胞,与周围的愈伤薄壁细胞有明显的界限(图3B),在其发展过程中,由近圆形的细胞团变成先端较尖的诱发根原基。这些根原基细胞继续伸长,突破愈伤组织表面伸出体外。伴随着根原基及不定根的发育,不定根内部维管系统同时发育,最终与插穗内的维管系统相连。扦插后16 d,由愈伤组织形成的不定根伸出插穗。
2.3.2 髓射线与皮层交界处薄壁细胞分化形成根原基

图3 愈伤组织内分化的根原基解剖结构Fig. 3 The anatomical structure of root primordium in the callus
红缨海棠插穗内部髓射线与皮层交界处存在一团分裂能力较强的细胞,该团细胞与周围的皮层薄壁细胞明显不同,细胞体积小、排列紧密、细胞质浓,染色较深(图4A)。这团细胞继续分裂分化可诱导成根原基,扦插12 d后,根原基发育成排列紧密的圆球状细胞(图4B)。扦插16 d后,圆球状的根原基继续发育,一端伸长,形似蝌蚪状(图4C)。髓射线与皮层交界处形成的根原基较多地发生于皮孔附近,由此处根原基发育形成的不定根可以较容易地由皮孔穿出;若形成根原基的部位不处于皮孔附近,不定根将突破皮层伸出体外,突破皮层时,根原基已发育成具有根冠、分生区和伸长区等较为完整的根尖组织(图4D)。这种根原基的发生、发育与愈伤组织的出现及发育几乎同时进行,所形成的不定根约在扦插后16~18 d伸出插穗。

图4 髓射线与皮层交界处根原基的解剖构造Fig. 4 The differentiation of root primordium
2.3.3 维管形成层分化形成根原基
红缨海棠插穗维管形成层具有较强的分裂能力,扦插后约10 d,维管形成层便恢复强烈的分裂能力,次生木质部与韧皮部间有5~6层分生细胞,形成层带明显加宽,细胞分裂分化更加活跃,向内向外分裂出大量的薄壁细胞。在髓射线与维管形成层交界处的薄壁细胞具有很强的分裂能力,形成根原基,这些细胞不断分裂并伸向皮层,最终发育成不定根伸出插穗(图5A、5B)。在突破表皮之前,整个不定根呈圆锥状,其根冠较为发达,既起到保护分生区的作用,又有利于突破表皮(图5C)。由维管形成层处形成的根原基至不定根突破表皮的时间不超过20 d。
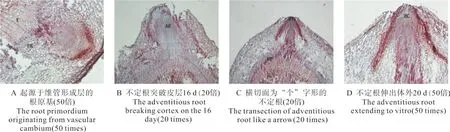
图5 维管形成层根原基的解剖结构Fig. 5 The anatomical structure of root primordium induced by vascular cambium
2.3.4 髓射线与环髓带交界处形成根原基

图6 髓射线与环髓带交界处诱发根原基Fig. 6 The root primordium induced between vascular rays and perimeter
枝条髓射线起源于环髓带,环髓带细胞较小、排列紧密、染色较深,细胞非常活跃。髓射线端部连接着环髓带,沿着环髓带呈发射状排列。红缨海棠插穗扦插后,与髓射线交界处的环髓带逐渐加宽(图6A),其细胞较小,排列整齐紧凑,细胞分裂形成规则的带状并向木质部延伸。此处细胞分裂延伸至与髓射线的交界处,髓射线加宽(图6B),形成根原基,此过程仅需8 d。插穗内根原基逐步发育成不定根,穿过维管形成层、韧皮部和皮层,最后突破表皮,此过程约需12 d。
3 讨论与结论
3.1 插穗根原始体类型
插穗形成的不定根是由根原始体发育而来的,分为潜伏根原始体或诱发根原始体[1],有些树种扦插不定根可产生于两种根原始体,如桑树[9],有些树木则只存在一种根原始体,如棱角山矾[10]、欧李[11]和华北五角枫[12]插穗均只有诱生根原始体。针对苹果属植物扦插研究报道较多,但涉及扦插不定根发生解剖学的研究较少,对于插穗中是否存在潜伏根原始体未开展论证。本实验证实,红缨海棠扦插前枝条内不存在潜伏根原始体,其扦插生根属于诱导生根型,这与许晓岗对垂丝海棠、海棠果、楸子的研究结论相符合[8,13-14]。
3.2 插穗诱发根原基发生部位
插穗扦插后分化形成的根原始体被称为诱发根原始体,它可以由维管形成层、韧皮薄壁组织细胞、韧皮射线、髓射线及其复合组织等部位产生,也可在愈伤组织中产生[2]。扦插繁殖过程中,不同树种诱发根原始体产生的部位不同,或来源于单一部位或来源于多个部位,如欧李插穗不定根原基起源于髓射线[11]、棱角山矾插穗根原基是由髓射线与形成层交接处的细胞分化而成[10]、华北五角枫不定根根原基起源于愈伤组织[12]、欧美杂种山杨[15]和柚木[16]不定根根原基起源于维管形成层,沙棘扦插根原基起源于维管形成层细胞处和韧皮部薄壁细胞区域[17],油茶诱生根源于形成层、韧皮部及愈伤组织等部位[18]。本实验证实,红缨海棠嫩枝插穗诱生根原基可产生于愈伤组织、髓射线与皮层交界处薄壁细胞、维管形成层、髓射线与环髓带交界处等4个部位,以往文献报道同属的垂丝海棠插穗诱生根原基形成于愈伤组织[8],而海棠果插穗愈伤组织内却未发现根原基,其诱生根原基由初生射线与维管形成层交汇处细胞的分裂分化形成[13]。如此亲缘关系较近的树种,插穗诱发根原基产生部位却存在很大差异,分析该问题的原因可能源于三个方面:其一,树种(品种)间诱发根原基产生部位存在很大差异,可能由树种的遗传特性所决定,如蒙椴诱生根原基源自髓射线与皮层交界处、愈伤组织、维管形成层,属于较易生根树种,而糠椴诱生根原基源自愈伤组织、维管形成层,属较难生根树种[19]。其二,扦插处理方法不同导致诱发根原基产生部位存在差异,加温处理的四倍体刺槐硬枝插穗根原基产生于髓射线与皮层交界处薄壁细胞和愈伤组织2个部位,不加温处理的硬枝插穗根原基则仅产生于愈伤组织[20];采用立体育苗的桑树插穗诱发根原基由维管形成层的薄壁细胞发育形成,而常规绿枝插穗诱发根原基却源自愈伤组织[9];笔者也曾证实IBA和NAA溶液均可促进红缨海棠插穗愈伤组织生根,NAA处理插穗还可促进插穗皮部生根[6]。其三,扦插穗条木质化程度不同,诱发根原基产生部位存在差异,四倍体刺槐硬枝插穗根原基产生于髓射线与皮层交界处薄壁细胞和愈伤组织2个部位,嫩枝插穗根原基产生于愈伤组织[20];中国沙棘木质化枝条根原基起源于木质部中的木射线细胞,而半木质化枝条根原基则发生于韧皮薄壁细胞与形成层之间[21]。
3.3 扦插生根的难易与生根类型之间的关系
插穗中若存在潜伏根原始体,扦插容易生根,扦插后需诱发根原始体的插穗则不容易生根[2]。笔者前期已经证实红缨海棠扦插属于较难生根树种,不使用生长调节剂处理的插穗生根率不足10%,且无皮部生根;使用IBA和NAA处理插穗后可以促进插穗生根,且生根多属于愈伤组织生根型,实验数据显示NAA处理有利于诱导插穗皮部生根[6]。同时,扦插生根的难度还与诱发根原始体部位的数量有关,若插穗中诱发根原基的部位较少,则不宜诱导生根,如欧美杂种山杨由于扦插不定根根原基起源单一导致扦插难生根[15]。本实验证实,诱发根原始体发生部位较多有利于生根,经NAA处理的红缨海棠嫩枝扦插不定根根原基来自于愈伤组织等4个部位,生根率接近70%[6],这与油茶扦插具有相似之处,其根原基可产生于形成层、韧皮部及愈伤组织等部位,生产实践证实油茶属于较易生根树种[18]。凡属难生根的树种,插条的不定根绝大多数是愈伤部位生根[22]。插穗是否形成愈伤组织与插穗生根难易因树种而不同,柚木嫩枝扦插所形成的愈伤组织与生根呈负相关[16],油茶不定根的发生与愈伤组织没有直接关系[18],华北五角枫[12]不定根根原基则仅来源于愈伤组织。本实验及前期实验已经证实,使用NAA和IBA处理插穗后,有利于诱导形成愈伤组织,且促进了愈伤组织生根,对于红缨海棠而言愈伤组织形成有利于扦插生根。
本研究以1 000 mg·L-1NAA溶液对嫩枝插穗进行促根处理,于扦插过程中取样制作石蜡切片,证实红缨海棠嫩枝扦插生根属于诱导生根型,诱生根原基可产生于愈伤组织等4个部位。本实验仅选用NAA单一浓度促根生根剂,未对NAA其他浓度生根剂和IBA等其他种类生根剂开展对比研究,其他生根剂对于插穗根原基的诱导作用及诱生根原基产生部位有待进一步研究。同时,实验过程中每次取样的穗条仅为3根,制作石蜡切片的数量也较为有限,后续实验中将加大取样数量和切片制作数量,以验证诱导根原基是否还有其他产生部位。
猜你喜欢
作物研究(2022年1期)2022-11-27
智慧农业导刊(2022年22期)2022-11-17
作物学报(2022年10期)2022-07-21
现代农业研究(2021年10期)2021-11-02
西藏人文地理(2020年4期)2020-11-19
农业工程技术·综合版(2020年1期)2020-04-23
植物研究(2020年6期)2020-03-05
计算机应用(2018年10期)2018-11-22
天然产物研究与开发(2018年4期)2018-05-07
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
