
杂交F1代myostatin基因编辑肉牛的肉质特性分析
2018-08-14 05:19高广琦魏著英白春玲韩红燕李光鹏
中国牛业科学 2018年3期
王 鑫,高广琦,魏著英,白春玲,佟 彬,韩红燕,张 立,李光鹏
(省部共建草原家畜生殖调控与繁育国家重点实验室,内蒙古大学,呼和浩特,010070)
1 前言
肉质的评价指标有多个方面,包括pH、肉色、嫩度、营养价值、风味等,这些肉质性状主要受遗传和环境因素影响[1]。我国的黄牛在历史上主要为役用,具有前驱发达、后驱成流线型、臀部肌肉欠发达等特点,生产性能不及国际上著名的肉牛品种[2]。近年来,随着人们对牛肉需求量的增加,如何改良本地黄牛品种,以提高其产肉性能,是我国肉牛育种工作的核心任务之一。多年来,主要是引进国外优良肉牛品种用于本地黄牛的杂交改良,虽然取得了显著成效[3-5],但也造成我国黄牛遗传种质的流失。对双肌型比利时蓝牛(Belgian Blue)和皮埃蒙特牛(Piemontese)的解析,发现肌肉生长抑制基因(myostatin,MSTN)是引起双肌表型的关键基因[6-8]。MSTN基因突变会使牛产生肌肉过度发达的性状,从而大幅提高肉牛的屠宰率和产肉率。利用双肌比利时蓝公牛与荷斯坦牛杂交后,其后代的活体重、胴体重和屠宰率等显著高于普通牛的杂交后代[9-10],但对于肌肉中脂肪含量和脂肪酸组成,在不同报道中有差异[9-11]。
为提高鲁西黄牛的生长与产肉性能,利用基因编辑技术对其MSTN进行定点突变,获得了双肌性状明显的基因编辑牛[12]。本研究以MSTN基因人工突变鲁西黄牛与中国西门塔尔牛的杂交F1代为材料,比较分析屠宰和肉质性状,为进一步改良牛肉的品质、培育高产肉牛新品种提供依据。
2 材料与方法
2.1 试验材料
2.1.1 试验动物 本研究所用公牛为经锌指核酸酶技术编辑的MSTN突变鲁西黄牛[12],以其为父本,以中国西门塔尔牛为母本,经人工繁育得到F1代牛,对照组为同龄的中国西门塔尔牛。由于纯系鲁西黄牛的购置困难及费用等问题,本试验并没有做基因编辑鲁西牛与普通纯系鲁西牛之间杂交,所用的鲁西牛屠宰数据来自《中国畜禽遗传资源志-牛志》[13]。所用牛群的生产与饲养管理等均在内蒙古内大圣牧高科牧业有限公司基地完成。
2.1.2 仪器设备 本试验所用仪器包括PH-STAR胴体肉质值直测仪(MATTH?US,德国)、C-LM3B 肌肉嫩度仪(Tenovo,中国)、Tenovo Meat-1 压肉机(Tenovo,中国)、B400均质仪(Buchi,瑞士)、SERIES 3000近红外肉质分析仪(NIR Technology,澳大利亚)、N-EVAP-111氮吹仪(Organomation,美国)、Centrifuge 5417R冷冻离心机、Centrifuge 5810R高速冷冻离心机(Eppendorf,德国)、Precellys 24生物样品均质器(Bertin Technologies,法国)和GCMS-QP2010 SE气相色谱-质谱联用仪(SHIMADZU,日本)等。
2.1.3 试剂耗材 本试验所用试剂中,氢氧化钾、甲醇、三氯甲烷为国产分析纯,正己烷为国产色谱纯。脂肪酸标准品为F.A.M.E.Mix, C4-C24100MG NEAT(SUPELCO,美国)。
2.2 试验方法
2.2.1 育肥与屠宰 本试验共选用8头公牛,试验组与对照组各4头。在同一环境、常规饲养管理方式下饲养,待其生长至24月龄时,进行屠宰。屠宰前禁食24 h,称量活体重;屠宰后,收集各器官组织样本,称量胴体重,计算屠宰率(胴体重/活体重×100%)。
胴体于0~4℃、95%湿度条件下,排酸24 h后进行分割。分别取每头牛的上脑、眼肌、米龙与腱子部位肌肉各1 kg左右,现场检测排酸后pH值,然后于4℃真空保存样品,运输至实验室,先分装小块肉样于真空条件下保存于-80℃冰箱,其余样品迅速处理(去除表面结缔组织和肌间脂肪)并完成系水力、剪切力、营养成分等检测。
2.2.2 pH检测 利用PH-STAR检测排酸24 h后的肉样pH值,每个部位检测3次,取平均值。
2.2.3 嫩度检测 对牛肉嫩度的检测包括剪切力和系水力两个方面。用采样器平行于肌纤维切下横截面积为1 cm2的肌肉条,于C-LM3B 肌肉嫩度仪检测样品的剪切力(kg·f)。另取一块约100 g的样品,切成方块,称量初始重量W1,然后用Tenovo Meat-1压肉机垂直于肌纤维开始挤压,参数为默认值,仪器停止后用吸水纸吸干表面水分,称量挤压后重量W2。本研究以失水率(Pressing Loss,PL)反映牛肉样品的系水力,计算公式为:PL=(W1-W2)/W1×100%。
2.2.4 营养成分检测 取约100 g肌肉样品,用B400均质仪搅拌均匀后,于SERIES 3000近红外肉质分析仪中检测样品中水分、脂肪、蛋白的组分,参数为默认值。
2.2.5 脂肪酸检测 对上脑、眼肌、米龙、腱子部位的肌肉样品,分别取约0.1 g放置于匀浆管中,加入1.5 ml氯仿-甲醇(2:1)溶液,低温匀浆2 min。在4℃条件下于4 000 r·min-1离心5 min,静置5 min后,然后吸上清于15 ml离心管中,该过程重复两次以充分提取样品中的脂类。在通风条件下,用氮吹仪吹干溶剂,先加入1 ml正己烷,震荡30 s以加速溶解,再加入1 ml正己烷并震荡30 s。室温静置过夜后加入1 mL饱和氢氧化钾-甲醇溶液进行甲酯化,剧烈震荡30 s, 在4℃条件下于4 000 r·min-1离心5 min,吸1ml上清于脂肪酸上样瓶中,利用气相色谱质谱联用仪检测脂肪酸含量。
色谱条件:色谱柱为HP-88高分离度气相色谱柱(100 m×0.25 mm×0.25 μm, Agilent,美国);柱箱60℃,进样口 220℃,分流比 20:1;进样量 1 μl;载气为氦气,总流量24 ml/min,柱流量 1 ml/min,线速度20 cm/s;升温程序:初温60℃,保持1 min,40℃/min升温至140℃,保持10 min,4℃/min升温至 240℃,保持15 min。质谱条件:离子源温度 200℃,接口温度 240℃,溶剂延迟时间为9.7 min,扫描范围50~500 m/z。脂肪酸的定性定量分析采用外标法。
2.2.6 统计学方法 本研究数据统计方法为平均值±标准差(±SD),显著性差异分析采用T-test。
3 结果与分析
3.1 屠宰性状
屠宰后,与对照组牛相比,MSTN杂交F1代牛的各器官的大小与色泽没有变化,内脏器官周围及器官之间的脂肪分布也没有明显差异。测量结果显示,试验牛平均活体重为653.50±65.16 kg,胴体重392.85±43.90 kg,屠宰率为60.07±1.69%;对照牛平均体重为632.93±71.12 kg,胴体重346.50±49.36 kg,屠宰率为54.63±2.08%;两组之间屠宰率具有显著性差异(P<0.05)(表1)。说明MSTN基因编辑牛的杂交F1代的产肉性能显著提高。与纯系鲁西黄牛资料数据比较[13],本研究的杂交F1代肉牛的胴体重和屠宰率均明显提高。
3.2 肉质特性
3.2.1 不同部位肌肉的pH 本研究中,试验组和对照组的上脑、眼肌、米龙、腱子4个部位牛肉的pH值之间没有显著差异(P>0.05),两组样品的pH值都在5.6左右,说明MSTN基因突变没有影响牛肉的pH值(表2)。
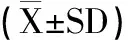
表1 试验牛与对照牛的屠宰性状
* 本组数据来自《中国畜禽遗传资源志-牛志》,为18~24月龄的鲁西公牛[13]

表2 不同部位牛肉的
3.2.3 不同部位肌肉的嫩度 为了进一步分析MSTN人工突变牛牛肉的嫩度,分别检测了不同部位肌肉的剪切力和系水力。结果表明,上脑和眼肌的剪切力低于米龙和腱子,压挤后的失水率也相对较低,说明这两个部位肌肉的嫩度较好。试验组牛肉相比于对照组,剪切力和系水力(失水率)在上脑、眼肌、米龙部位略高,腱子部位略低,然而都没有显著性差异(P>0.05),说明不同部位的两种牛肉嫩度相似(表3)。
表3 不同部位牛肉的嫩度
3.2.4 不同部位肌肉的营养成分组成 本研究中,主要对牛肉中水分、蛋白质、脂肪的含量进行了测定。经近红外光谱检测得出,牛肉中的水分含量约为70%,蛋白含量约为20%,不同部位的肌内脂肪含量为4.7%~10.1%。其中,试验牛和对照牛的上脑和眼肌的肌内脂肪含量较高,说明这两个部位的牛肉更有利于脂肪的沉积。比较分析发现,试验组4个部位的牛肉中,肌内脂肪含量均低于对照组,其中上脑和眼肌具有显著性差异(P<0.05)(图1)。结果说明MSTN基因突变会减少F1代牛肌内脂肪的沉积。

图1 不同部位牛肉的营养成分组成
3.2.4 不同部位肌肉的脂肪酸组成 为了评价肌内脂肪的减少是否影响到肉质脂肪酸的含量,本研究进一步对肉质的脂肪酸组成进行了分析。结果如表4所示,MSTN基因突变牛与对照牛相比,脂肪酸组成没有发生显著改变(P>0.05)。不同部位的牛肉中,饱和脂肪酸(Saturated fatty acids,SFA)是最主要的组成部分,为38.05%~51.61%,其中棕榈酸(C16:0)和硬脂酸(C18:0)含量最高。SFA在腱子的占比少于其它3个部位,对于这一结果,试验组和对照组有相同的趋势。不饱和脂肪酸中,单不饱和脂肪酸(Monounsaturated fatty acid,MUFA)含量较多,主要为棕榈烯酸(C16:1)和油酸(C18:1n9c),多不饱和脂肪酸(Polyunsaturated fatty acids,PUFA)主要为亚油酸(C18:2n)、二十烷三烯酸(C20:3n6、C20:3n3)和花生四烯酸(C20:4n6)。试验牛和对照牛的眼肌部位PUFA比例低于各自的其它3个部位。PUFA/SFA比值是评定肉中脂肪对人体健康的一个指标,这一比值同样在眼肌部位最低,在腱子中最高,但在试验组和对照组相应部位的比较中不存在显著性差异(P>0.05)。
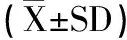
表4 不同部位牛肉中脂肪酸的含量
SFA:饱和脂肪酸;MUFA:单不饱和脂肪酸;PUFA:多不饱和脂肪酸;PUFA/SFA:多不饱和脂肪酸与饱和脂肪酸的比值
4 讨论
4.1 MSTN基因编辑牛的杂交F1代的产肉性能显著提高
在以提高产肉性能为目的的肉牛育种研究中,往往采用肌肉增长快的双肌牛为父本进行杂交选育,常用的品种包括比利时蓝牛、皮埃蒙特牛和超级夏洛莱牛。Domingo等发现,10月龄的比利时蓝牛×荷斯坦牛的屠宰率可达59.44±1.22%,显著高于利木赞×荷斯坦牛(55.17±1.26%)和Rubia Gallega×荷斯坦牛的杂交种(54.86±1.43%)[9]。同样,Keady等分别利用比利时蓝牛和安格斯牛与荷斯坦牛杂交,并于299d时屠宰。结果显示,比利时蓝×荷斯坦杂交牛平均胴体重为369 kg,安格斯×荷斯坦杂交牛平均胴体重为354 kg,二者之间有显著性差异(P=0.02)[10]。国内学者的统计数据表明,比利时蓝改良 F1 代肉牛屠宰率达到 69.2%,比本地黄牛屠宰率56.2%提高了13个百分点,提高幅度达到 23.1%[14]。另外,用皮埃蒙特公牛对南阳牛[15]、晋南牛[16]、柴达木本地黄牛[17]、辽育白牛[18]的杂交改良也显著提高F1代杂交牛的屠宰率。
据报道,18~24月龄的纯系鲁西公牛,其活体重、胴体重和屠宰率分别为512.5±95.5 kg、284.0±52.3 kg和55.4±0.1%;18~22月龄的育肥中国西门塔尔阉牛的活体重和屠宰率分别为573.6±69.9 kg和60.4±4.9%[13]。在利用西门塔尔与鲁西牛的杂交试验中,约20月龄的育肥西鲁杂交牛屠宰率可达60.5%~60.96%[19-20],说明西鲁杂交牛的生产性能与中国西门塔尔牛基本一致。在本研究中,选用了中国西门塔尔牛为母本与MSTN基因人工突变鲁西牛杂交,MSTN杂交F1代牛的屠宰率既显著高于纯系鲁西牛,也高于中国西门塔尔牛。说明利用MSTN基因编辑鲁西黄牛的杂交能够提高F1代肉牛的生产性能,这一结果与利用自然突变牛杂交改良的效果是一致的。因此,同MSTN基因自然突变牛一样,人工突变牛的F1代的生产性能与产肉性能等都显著提升。另外,本试验所用的基因编辑牛为纯种鲁西牛,只是将其MSTN基因的一小段碱基删除了,仍保留鲁西牛几乎完整的基因组,也即保留了鲁西牛的遗传种质。
4.2 MSTN基因编辑对杂交F1代肉牛肌肉的pH值和嫩度的影响
屠宰前牛肌肉的pH值通常为中性,屠宰后在排酸过程中,由于糖酵解积累乳酸,使pH值下降,能够减少微生物侵染,有利于肉产品的保存[21]。pH是衡量牛肉品质的一个关键参数,与牛肉系水力和肉色等性状相关。屠宰后的牛肉,肌肉内糖原酵解速率过慢会导致pH过高,使肌肉颜色变暗,肉质表面干硬,形成DFD肉(Dark,Firm,Dry)[22]。相反,如果屠宰后pH降低速率过快,会影响肌肉中钙蛋白酶(Calpain)的活性,降低肉品的系水力,从而导致血水渗出,形成PSE肉(Pale,Soft,Exudative)。本研究中,杂交F1代MSTN基因编辑牛4个部位牛肉的pH值与对照牛相比没有显著差异(P>0.05),并且pH值在正常范围内(5.6左右),说明MSTN基因突变没有影响牛肉的pH值。结果与比利时蓝牛杂交对肉质pH值的影响相一致[9]。
作为肉质评价的重要指标之一,嫩度与肌纤维和肌纤维相关蛋白的水解作用有关[23]。本研究结果显示,杂交F1代MSTN基因编辑肉牛不同部位肌肉的剪切力和系水力与对照牛相比都没有显著性差异(P>0.05),说明其肉质嫩度没有受到影响。有研究表明,双肌牛牛肉的嫩度偏低,可能是由于肌肉中胶原蛋白含量较少所致[24-25],然而Wiener等并没有发现肉质的嫩度与MSTN缺失突变存在显著的相关性[26]。唐中伟等研究结果同样表明,皮埃蒙特杂交F1代牛的肉质嫩度与对照组相近[16]。
4.3 MSTN基因编辑对杂交F1代肉牛肌肉的肌内脂肪和脂肪酸组成的影响
MSTN基因功能的缺失,通常会导致牛肉肌内脂肪含量的减少[27],这一现象出现在纯合突变的比利时蓝牛[28]及杂合突变的F1代牛中[9, 29]。本研究中,杂交F1代MSTN基因编辑肉牛上脑和眼肌部位的肌内脂肪含量显著低于对照牛,米龙和腱子部位尽管脂肪含量偏低,但没有显著差异,说明杂交F1代肉牛MSTN基因的突变主要影响肌内脂肪在背最长肌的沉积。低脂肪、高蛋白的牛肉符合当今人们追求的健康饮食,然而脂肪低于3%会影响牛肉的适口性[30]。本文所研究的MSTN基因突变肉牛,其各部位牛肉的肌内脂肪含量尽管低于对照牛,但仍在合理范围之内(大于3%)。
在以往的研究中,MSTN基因自然突变的双肌牛(MSTN-/-)牛肉含有更高含量的PUFA[26, 28, 31],可能是由于MSTN基因突变影响到细胞膜中磷脂的组成,进而改变了肌肉中PUFA的含量[27]。然而本研究结果发现,利用MSTN基因人工突变的肉牛杂交产生的F1代个体中,PUFA含量以及PUFA/SFA比值与对照牛相比没有显著差异,这也许是因为本研究所用的F1代肉牛的基因型为MSTN杂合突变(MSTN+/-),在基因调控上并没有完全的失去功能。
5 结论
鲁西黄牛主要产于山东西南部,是我国役、肉兼用型优良品种,但其生长缓慢、后驱欠发达,经济效益低于国际上著名的肉牛品种[32]。中国西门塔尔牛是20世纪50~80年代引进的德系、苏系、奥系西门塔尔牛与本地黄牛进行级进杂交选育而成的乳肉兼用品种,其生产性能和肉质通常优于纯种鲁西黄牛[3, 33, 34]。
本研究利用MSTN基因人工突变的鲁西黄牛与中国西门塔尔牛进行繁育,F1代杂交牛表现出优于父母本的生产性能,而且肉质品质没有受到影响,表明利用MSTN突变技术可以实现改良国内黄牛的目的。
猜你喜欢
今日农业(2022年2期)2022-11-16
美食(2022年4期)2022-04-16
国外畜牧学·猪与禽(2020年8期)2020-11-30
中国牛业科学(2020年4期)2020-08-15
美食(2020年5期)2020-06-01
农家致富顾问·下半月(2018年1期)2018-10-21
肉类研究(2017年10期)2017-11-20
湖北农业科学(2016年18期)2016-12-08
肉类研究(2016年6期)2016-07-22
肉类研究(2016年4期)2016-05-17
