
毛状副冠对高山植物喉毛花的适应意义
2018-09-10 01:17张婵张新米兆荣黄雨晗马剑敏
广西植物 2018年2期
张婵 张新 米兆荣 黄雨晗 马剑敏


摘 要: 高山地区生境极端,却拥有许多形态特化的植物,非生物因素在塑造花部性状及其进化过程中发挥着重要作用。该研究选取龙胆科典型高山植物喉毛花(Comastoma pulmonarium)为对象,探究其毛状副冠在多雨、强辐射的极端高山环境中的适应意义及其对植物雌雄繁殖适合度的影响。结果表明:通过比较自然状态和人工去除副冠的花,毛状副冠有效减少了雨水对花粉的冲刷(t=2.61,P<0.05),提高了受精比率(t=2.05,P<0.05),但是对种子的质量,即种子重量和种子萌发率的影响不显著。另外,花粉浸泡在蒸馏水中后,其萌发率显著低于蔗糖溶液中(t=30.67,P<0.001),表明毛状副冠能够有效减小雨水浸泡对花粉活力的影响;同时,与自然状态相比,去除毛状副冠后的花,经太阳暴晒后,其花粉萌发率同样极显著地降低(t=9.89,P<0.001),表明毛状副冠有效地避免了太阳辐射对花粉质量产生的不利影响。该研究结果表明喉毛花的毛状副冠结构是应对高山恶劣环境的一种适应策略,对植株的雌性和雄性繁殖成功都具有重要意义,从而进一步证实了非生物因素在植物花部特征演化中的重要作用。
关键词: 高山环境, 喉毛花, 毛状副冠, 繁殖适合度
中图分类号: Q948
文献标识码: A
文章编号: 1000-3142(2018)02-0153-06
Adaptive significance of hairy corona to an alpine plant Comastoma pulmonarium
ZHANG Chan1, ZHANG Xin1, MI Zhaorong2, HUANG Yuhan1, MA Jianmin1*
( 1. College of Life Sciences, Henan Normal University, Xinxiang 453007, Henan, China; 2. Farmland Irrigation Research Institute, Chinese Academy of Agricultural Sciences, Xinxiang 453003, Henan, China )
Abstract: Despite of the harsh environmental conditions, alpine regions have very specialized and diverse plants. Abiotic factors play important roles in the formation and evolution of floral traits of these alpine plants. In this paper, the hairy corona of alpine plant Comastoma pulmonarium was studied to explore the evolutionary and adaptive significance of this distinctive floral structure in the extreme alpine environment and its effects on plant reproductive fitness. The results showed that in artificially corona removed flowers, pollen grains in anthers (t = 2.61, P<0.05) and fertilization rate (t=2.05, P<0.05) decreased greatly after rain-wash. However, there were no significant differences of the seed weight and seed germination rate between artificially corona removed flowers and control flowers. Additionally, pollen germination rate reduced significantly due to water immersion (t = 30.67, P<0.001) and direct exposure to sunlight (t = 9.89,P<0.001), indicating that hairy corona could maintain viability of pollen grains in front of rain immersion and intense solar radiation. This study indicates that the hairy corona of C. pulmonarium has adaptive significance for both male and female reproductive success in the harsh environment of alpine regions, and further confirms selection on the flower morphology of alpine plants by abiotic factors.
Key words: alpine environment, Comastoma pulmonarium, hairy corona, reproductive fitness
自然界中花部特征的多樣性及其进化反映了植物对生长环境的长期适应与响应,是权衡了多种选择压力的结果(Brown,2002)。以往的研究更多地关注传粉者对花部特征的选择压力(Darwin,1859;Fenster et al,2004),然而温度、湿度、降水和光照等非生物因素在塑造花部性状及其进化过程中同样发挥重要的作用(王赟,2009)。尤其在环境恶劣的高海拔地区,具有气候多变、昼夜温差极大、暴雨频发、紫外辐射强等不利于植物生长的环境因子,因此高山植物的花部特征通常表现出对环境高度特化的响应(彭德力等,2012)。
目前,研究最多的非生物因素之一是雨水。雨水会冲刷掉花药和柱头上的花粉、降低花粉萌发力以及稀释花蜜浓度,从而影响植株的繁殖适合度(Huang et al,2002)。防雨的花部机制有多种表现形式,如在多雨的生境下很多植物会形成下垂花冠,有效地避免了雨水的伤害(Tadey & Aizen,2001;Huang et al,2002;Aizen,2003);低温诱导的暂时性花闭合现象,也是对高山夏季暴风雨频繁发生的适应机制(Bynum & Smith,2001;He et al,2005);再如珙桐(Davidia involucrata)宿存的花苞片能夠有效地减少降雨时花粉粒的损失(Sun et al,2008)。
除了雨水,太阳辐射也会严重降低花粉活性(Demchik & Day,1996;冯虎元等,2002),尤其在高山环境中,随着海拔高度上升,太阳辐射越来越强烈,植物也相应地演化出不同的花部结构以适应强烈的太阳辐射。如Zhang et al(2014)通过对横断山脉的 42 种高山植物两种不同花结构特征研究表明,花药暴露型的植物与花药被花部结构保护型的植物相比,其花粉对紫外辐射的敏感性较低,这证明紫外辐射对高山植物花部的形态演变具有重要作用。花的特殊结构或形态可以保护敏感的花粉远离太阳辐射带来的伤害,如铃铛子(Anisodus luridus)和赛莨菪(A. carniolicoides)下垂的花冠能有效保护花药上的花粉既不受雨水的冲刷,也不受强烈太阳辐射的照射,对维持花粉数量和花粉活力都有重要意义,能显著提高植株的雄性适合度(王赟,2009)。
传粉者与非传粉者因素在塑造花部特征及其进化过程中都发挥重要作用,但当非传粉者因素与传粉者产生的选择压力同向时,非传粉者因素产生的影响常被传粉者掩盖,进而被研究者忽视(Strauss & Whittall,2006)。本研究选取的对象高山植物喉毛花(Comastoma pulmonarium)在海北地区完全通过自交方式繁殖(Zhang et al,2011),因此可以排除传粉者选择压力的干扰,从而为准确评价非传粉者因素在花部特征演化中的作用提供一个理想的研究材料。喉毛花是龙胆科一年生植物,其花冠裂片基部有一圈特殊的白色毛状副冠,完全封盖住冠筒口,是龙胆科喉毛花属极为特化的花部特征,其生态功能未见报道。喉毛花开花时间主要集中在6—8月,正值青藏高原地区夏季暴雨频发、太阳辐射最为强烈的时期,排除传粉者的选择压力,我们推测该毛状副冠结构可能对其适应高山环境有重要意义。因此,本研究选取喉毛花为研究对象,重点回答以下两个问题:(1)毛状副冠对高山极端环境(如频繁降雨和强烈太阳辐射等)有何适应意义?(2)毛状副冠对喉毛花雌雄繁殖适合度分别有何影响?
1 材料与方法
1.1 研究地点
本研究于 2016年7—8月,在中国科学院海北高寒草甸生态系统定位观测站(以下简称海北站)(101°12′—101°23′ E,37°29′—37°45′ N)附近开展。海北站位于青藏高原东北部祁连山谷地,海拔3 200~3 600 m,属于高原大陆性气候,年内没有绝对无霜期,年极端最高气温和最低气温分别为27.6 ℃、-37.1 ℃,年降雨量在426~860 mm,其中80%的降水集中在5—9月。海北站附近的植被类型为青藏高原典型的地带性植被,有以金露梅(Potentilla fruticosa)为建群种的高寒灌丛草甸和以矮嵩草(Kobresia humilis)或高山嵩草(K. pygmaea)为建群种的高寒嵩草草甸(李英年等,2004)。
1.2 研究材料
喉毛花是龙胆科喉毛花属的一年生草本植物,主要生长在海拔3 000~4 800 m范围内,花单生茎顶或枝顶,主花轴通常产生数个侧枝。喉毛花的花冠呈管状,裂片基部有一圈白色毛状副冠,开花时全部副冠向心弯曲,封盖住冠筒口,将雄蕊和柱头完全遮盖在花冠筒内。雄蕊5个,着生于花冠筒上。果实为蒴果,种子小,表面光滑。在海北站地区喉毛花主要通过自交产生种子,并且存在严重的近交衰退(Zhang et al,2011)。
1.3 研究方法
1.3.1 对雄性适合度的影响
1.3.1.1 雨水冲刷对花粉数量的影响 随机选取并标记40个植株的顶花花蕾,平均分为两组,一组在花刚刚开放时,将毛状副冠去掉;一组保持自然状态作为对照。喉毛花的单花花期为4~5 d,雄先熟,散粉期为2~3 d,在各株顶花开花散粉后注意观察,若在花期第2 天18:00前经历大于1 h的较强降雨过程则计入最后数据,否则需重新标记并处理新的顶花花蕾。经历降雨的40朵顶花花蕾在花期第2天18:00,用镊子摘取5枚雄蕊,分别放入装有1 mL FAA(福尔马林∶乙酸∶无水乙醇=5∶5∶90)的离心管中固定,带回实验室。室内加入少量的洗涤剂和70%的乙醇定容到2 mL,将其充分摇匀,使剩余花粉完全分散在溶液中。然后,用微量移液器取5 μL至载玻片上,在显微镜下统计花粉数目,计数重复10次,计算每朵花的平均剩余花粉数量。
1.3.1.2 水浸泡对花粉质量的影响 随机选取并标记50朵即将开放的顶花花蕾,当花开始散粉后,每朵随机摘取1枚雄蕊,平均分为2组。一组的25枚雄蕊分别放入装有10%蔗糖溶液的离心管中,另一组的25枚雄蕊分别放入装有蒸馏水的离心管中。两组花粉粒萌发8 h 之后,在显微镜下观察萌发情况,对花粉粒总数和萌发的花粉粒数目分别进行统计,最后计算两组处理相应的花粉萌发率。
1.3.1.3 太阳辐射对花粉质量的影响 随机选取并标记50朵即将开放的顶花花蕾,平均分为两组,一组在花刚刚开放时,将毛状副冠去掉;一组保持自然状态作为对照。当花开放并散粉后,待太阳直接照射 5 h 后,每朵花随机摘取一枚雄蕊,分别放入装有10%蔗糖溶液的离心管中。两组花粉粒萌发8 h 之后,在显微镜下观察萌发情况,对花粉粒总数和萌发的花粉粒数目分别进行统计,最后计算两组处理相应的花粉萌发率。
1.3.2 对雌性适合度的影响
1.3.2.1 受精比率 由于喉毛花的败育种子在果实成熟时会消失而难以统计结籽率,但是其未受精的胚珠和受精胚珠可以清晰区分,因此用受精比率代替结籽率加以统计。首先,随机选取并标记40个植株的顶花花蕾,平均分为两组,一组在花刚刚开放时,将毛状副冠去掉;一组保持自然状态作为对照。开花两周后,摘取各株顶花花朵,带回实验室内解剖每朵花的子房,在显微镜下分别统计子房内的发育胚珠数和未发育胚珠数,并据此计算受精比率(受精比率=發育胚珠数/总胚珠数)。
1.3.2.2 种子重量 随机选取并标记40个植株的顶花花蕾,平均分为两组,一组在花刚刚开放时,将毛状副冠去掉;一组保持自然状态作为对照。待各株的顶花果实发育成熟后,采集分装在纸袋内带回实验室,将各组种子在阴凉通风处自然干燥一个月,然后用万分之一电子天平称量每组的种子重量。称量时每100 粒为一个单位,每组重复称量15次。
1.3.2.3 种子萌发率 种子称量结束后,先将两组种子在-20 ℃ 冰箱内进行春化处理。1个月后,在24 ℃的人工气候培养箱内进行萌发试验。每组种子随机挑选20个放到直径为6 cm的小培养皿上,培养皿表面包裹湿润的滤纸,每组做3个重复,随后将这3个小培养皿放到直径为15 cm的大培养皿中,在大培养皿底部保持加入少量的水。每天对每个小培养皿上的萌发种子数(以胚根长度>0.5 mm 时记作萌发)进行统计,一天统计2次并把发霉的种子及时移出培养皿,且要记录数量。若萌发的种子数连续5 d没有增加,则停止试验并计算种子的萌发率。
1.3.3 数据统计 利用SPSS 16.0 for Windows软件进行平均值和标准误的计算,用独立样本t检验(Independent t Test)比较两组数据之间的差异性。
2 结果与分析
2.1 对雄性适合度的影响
2.1.1 雨水冲刷对花粉数量的影响 去副冠的花与自然状态相比,其剩余花粉数显著降低(表1)。
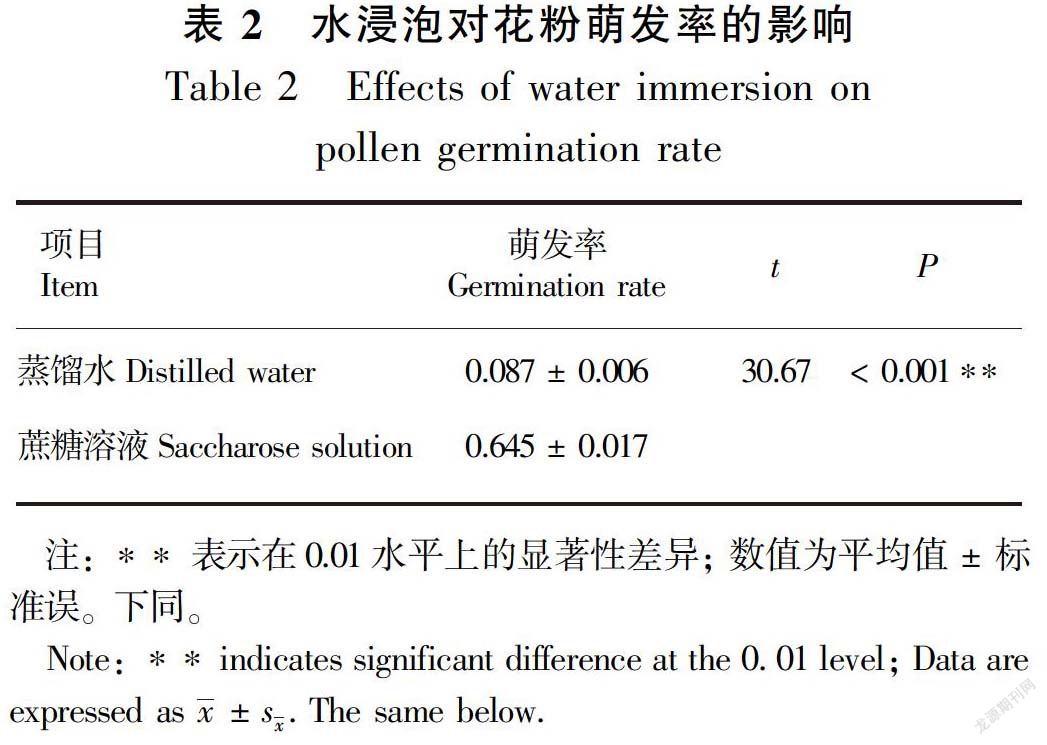
2.1.2 水浸泡对花粉质量的影响 在蒸馏水中浸泡了8 h的花粉萌发率低于在10%蔗糖溶液中浸泡相同时间的花粉萌发率,达到了极显著差异水平(表2)。
2.1.3 太阳辐射对花粉质量的影响 去副冠的花与自然状态相比,其花粉在10%蔗糖溶液中浸泡8 h的萌发率极显著地降低(表3)。
2.2 对雌性适合度的影响
去副冠的花与自然状态相比,其受精比率显著降低,但种子重量和种子萌发率差异不显著(表4)。
3 讨论与结论
高山带降水事件频发并伴随着降温,防雨策略对高山植物的繁殖成功具有重要意义。雨水会冲刷掉花药和柱头上的花粉(王赟,2009),对花粉粒造成不可逆的损伤(Corbet & Plumridge,1985;Jacquemart,1996),以及阻止花粉在柱头上的萌发(Bynum & Smith,2001),从而最终降低植物的繁殖适合度。例如,Bynum & Smith(2001)研究结果表明,雨水通过冲刷柱头花粉而明显降低了高山龙胆(Gentiana algida)的种子产量和质量。去除喉毛花的毛状副冠后,其剩余花粉数显著降低,另外,花粉浸泡在蒸馏水中后,其萌发率显著低于蔗糖溶液中。这表明,失去了毛状副冠的保护,雨水不仅会冲刷掉花药和柱头上的花粉,而且雨水的浸泡会极显著降低柱头上花粉的萌发率,两者的共同作用进一步导致胚珠的受精比率显著降低,但是对种子的质量,即种子重量和种子萌发率的影响不显著。以上结果表明,在多雨的高山环境中,喉毛花的毛状副冠有一定的防雨功能,能够显著提高植物的雄性和雌性繁殖适合度。
此外,与自然状态相比,去除毛状副冠后的花,经太阳暴晒5 h后,其花粉在蔗糖溶液中的萌发率同样极显著地降低。前人研究表明,大部分物种的花粉粒对温度都极其敏感,因此,高温暴晒可能导致花粉粒失水过度,从而使花粉活力严重受损(Speranza et al,1997;Heslop-Harrison,2000)。此外,花粉活力的降低还可能与高原强烈的紫外辐射有关,紫外辐射对花粉粒的萌发以及花粉管的生长都有明显的抑制作用(Torabinejad et al,1998;冯虎元等,2002),而且夏季正值青藏高原地区紫外辐射最强的时期(祝青林等,2005)。虽然本研究并不能确定太阳辐射后花粉活力显著下降的具体原因,但喉毛花的毛状副冠确实有效地避免了太阳辐射对花粉质量产生的不利影响。
在植物生长的各个阶段中,繁殖期对环境最为敏感(孙儒泳等,2002),高山植物生长季短,大部分植物的花期集中在6—8月,而此时频繁且不可预测的降雨以及强烈的紫外辐射是青藏高原地区的典型气候特征,严酷的环境严重影响了植物的繁殖适合度。本研究表明,喉毛花的毛状副冠结构是应对高山恶劣环境的一种适应策略,有利于维持花粉数量和花粉质量,从而进一步提高了胚珠受精比率,对植株的雌性和雄性繁殖成功都具有重要意义。目前,本研究仅在一个生长季内的一个喉毛花居群内开展,然而高山生态环境因子在不同时间和不同空间内可能存在很大变异,因此,在今后的研究工作中,应该涉及同一年内的多个居群或者同一居群的不同年份,从而对高山植物特殊花部结构的进化意义进行更为全面而准确的评价。
参考文献:
AIZEN MA, 2003. Down-facing flowers, hummingbirds and rain [J]. Taxon, 52(4):675-680.
BROWN K, 2002. Floral evolution: A compromise on floral traits [J]. Science, 298(5591):45-46.
BYNUM MR, SMITH WK, 2001. Floral movements in response to thunderstorms improve reproductive effort in the alpine species Gentiana algida (Gentianaceae) [J]. Am J Bot, 88(6):1088-1095.
CORBET SA, PLUMRIDGE JR, 1985. Hydrodynamics and the germination of oil-seed rape pollen [J]. J Agric Sci, 104(2):445-451.
DARWIN C, 1859. The origin of species by means of natural selection: or, the preservation of favored races in the struggle for life [M]. London, UK: John Murray.
DEMCHIK SM, DAY TA, 1996. Effect of enhanced UV-B radiation on pollen quantity, quality, and seed yield on Brassica rapa (Brassicaceae) [J]. Am J Bot, 83(5):573-579.
FENG HY, AN LZ, TAN LL, et al, 2002. Cumulative effect of solar ultraviolet-B radiation on pollen germination and tube growth of 19 species in vitro [J]. Chin J Appl Ecol, 13(7):814-818. [馮虎元, 安黎哲, 谭玲玲, 等, 2002. UV-B 辐射对植物花粉萌发率和花粉管生长的累积效应 [J]. 应用生态学报, 13(7):814-818.]
FENSTER CB, ARMBRUSTER WS, WILSON P, et al, 2004. Pollination syndromes and floral specialization [J]. Ann Rev Ecol Evol Syst, 35:375-403.
HE YP, DUAN YW, LIU JQ, et al, 2005. Floral closure in response to temperature and pollination in Gentiana straminea Maxim. (Gentianaceae), an alpine perennial in the Qinghai-Tibetan Plateau [J]. Plant Syst Evol, 256(1-4):17-33.
HESLOP-HARRISON Y, 2000. Control gates and micro-ecology: The pollen-stigma interaction in perspective [J]. Ann Bot, 85(Suppl.1):5-13.
HUANG SQ, TAKAHASHI Y, DAFNI A, 2002. Why does the flower stalk of Pulsatilla cernua (Ranunculaceae) bend during anthesis? [J]. Am J Bot, 89(10):1599-1603.
JACQUEMART AL, 1996. Selfing in Narthecium ossifragum (Melanthiaceae) [J]. Plant Syst Evol, 203(1-2):99-110.
LI YN, ZHAO XQ, CAO GM, et al, 2004. Analyses on climates and vegetation productivity background at Haibei Alpine Meadow Ecosystem Research Station [J]. Plateau Meteorol, 23(4):558-567. [李英年, 赵新全, 曹广民, 等, 2004. 海北高寒草甸生态系统定位站气候、植被生产力背景的分析 [J]. 高原气象, 23(4):558-567.]
PENG DL, ZHANG ZQ, NIU Y, et al, 2012. Advances in the studies of reproductive strategies of alpine plants [J]. Biodivers Sci, 20(3):286-299. [彭德力, 张志强, 牛洋, 等, 2012. 高山植物繁殖策略的研究进展 [J]. 生物多样性, 20(3):286-299.]
SPERANZA A, CALZONI GL, PACINI E, 1997. Occurrence of mono- or disaccharides and polysaccharide reserves in mature pollen grains [J]. Sex Plant Reprod, 10(2):110-115.
STRAUSS S, WHITTALL JB, 2006. Non-pollinator agents of selection on floral traits [M]//HARDER LD, BARRETT SCH. Ecology and Evolution of Flowers. Oxford:Oxford University Press:120-138.
SUN JF, GONG YB, RENNER SS, et al, 2008. Multifunctional bracts in the dove tree Davidia involucrata (Nyssaceae: Cornales): Rain protection and pollinator attraction [J]. Am Nat, 171(1):119-124.
SUN RY, LI QF, NIU CJ, et al, 2002. Basic ecology [M]. Beijing: Higher Education Press. [孙儒泳, 李庆芬, 牛翠娟, 等, 2002. 基础生态学 [M]. 北京:高等教育出版社.]
TADEY M, AIZEN M, 2001. Why do flowers of a hummingbird-pollinated mistletoe face down? [J]. Funct Ecol, 15(6):782-790.
TORABINEJAD J, CALDWELL MM, FLINT SD, et al, 1998. Susceptibility of pollen to UV-B radiation: An assay of 34 taxa [J]. Am J Bot, 85(3):360-369.
WANG Y, 2009. Pollination mechanism and ecological adaptation in Anisodus luridus and A. carniolicoides [D]. Kunming:Kunming Institute of Botany, Chinese Academy of Sciences:12-14. [王赟, 2009. 铃铛子(Anisodus luridus)和赛莨菪(A. carniolicoides)的传粉机制与生态适应 [D]. 昆明:中国科学院昆明植物研究所:12-14.]
ZHANG C, IRWIN RE, WANG Y, et al, 2011. Selective seed abortion induced by nectar robbing in the selfing Comastoma pulmonarium [J]. New Phytol, 192(1):249-255.
ZHANG C, YANG YP, DUAN YW, 2014. Pollen sensitivity to ultraviolet-B (UV-B) suggests floral structure evolution in alpine plants [J]. Sci Relp, 4:4520.
ZHU QL, YU GR, CAI F, et al, 2005. Spatiaization research on ultraviolet radiation in China [J]. Resourc Sci, 27(1):108-113. [祝青林, 于貴瑞, 蔡福, 等, 2005. 中国紫外辐射的空间分布特征 [J]. 资源科学, 27(1):108-113.]
