
鄱阳湖苦草及马来眼子菜PSⅡ荧光参数对水深变化的光响应
2018-09-10 19:50高桂青吕顺华吕念泽
广西植物 2018年12期
关键词:水深
高桂青 吕顺华 吕念泽
摘要: 以典型沉水植物苦草和馬来眼子菜为材料, 利用水下饱和脉冲调制叶绿素荧光仪研究不同水深(0.5、1.0、1.5、2.0、2.5 m)对两种植物叶片最小荧光(Fo)、最大荧光(Fm)、 PSⅡ最大光化光效率(Fv/Fm)、有效量子产量[Y(Ⅱ)]、光化学淬灭系数(qP)、非光化学淬灭系数(qN)、非调节性能量耗散的量子产量[Y(NO)]等荧光参数的影响。结果表明:水深1.5~2.0 m处苦草生物量最大,而1.0~1.5 m处马来眼子菜的最大;两种植物的Fo均先降低后升高,而荧光参数[(Fm、Fv/Fm、Fv/Fo、Y(Ⅱ)、qP]均先升高后降低;2.0 m处苦草的Fv/Fm、Fv/Fo最大,1.5 m处马来眼子菜的最大;相同水深下,马来眼子菜的qN比苦草低,与qP变化趋势相反;苦草的Y(Ⅱ)最大值出现在水深1.5~2.0 m内,马来眼子菜的Y(Ⅱ)最大值出现在1.5 m处;两者的Y(NO)随水深变化均表现出显著差异,过高或过低水深均抑制植物生长;相对光合电子传递速率(ETR)在不同水深处理间均差异显著,苦草的最大ETR比马来眼子菜小,说明其有较强的耐弱光能力。综上所述,在水深1.5~2.0 m苦草光合能力最强,最适宜生长;水深1.0~1.5 m最适宜马来眼子菜生长。-
关键词: 苦草, 马来眼子菜, 水深, 叶绿素荧光, 光响应曲线-
中图分类号: Q948.1, X824
文献标识码: A
文章编号: 1000-3142(2018)12-1626-09--
Light-response of PS Ⅱ fluorescence parameters on -Vallisneria natans and Potamogeton malaianus -to various water depths in Poyang Lake
-
GAO Guiqing1,2, L Shunhua2, L Nianze2, LU Long1*, -LI Wei2, JI Yong2, YOU Jikang2, WAN Peng2
-
( 1. School of Resource Environment and Chemical Engineering, Nanchang University, Nanchang 330031, China; -2. School of Civil and Architecture Engineering, Nanchang Institute of Technology, Nanchang 330099, China )
-
Abstract: In order to study the effects of water depth on the chlorophyll fluorescence characteristics of typical submerged macrophyte, Vallisneria natans and Potamogeton malaianus were selected as test materials in Poyang Lake wetland. Water levels (0.5,1.0,1.5,2.0,2.5 m) were controlled by buckets, minimum fluorescence ( Fo), maximum fluorescence (Fm), the maximum actinic light efficiency of PS Ⅱ (Fv/Fm), effective quantum yield [Y(Ⅱ)], photochemical quenching coefficient (qP ), non-photochemical quenching coefficient (qN ), unregulated energy dissipation quantum yield [Y(NO)] were measured by a submerged, modulated fluorescence spectrometer (Diving-pam). The variation of each parameter with depths was explored. The results showed that the maximum biomass of Vallisneria natans appeared in 1.5-2.0 m water depth, and that of Potamogeton malaianus appeared in 1.0-1.5 m water depth. Fo of each macrophyte decreased first and then increased, while the fluorescence parameters [Fm, Fv/Fm, Fv/Fo, Y(Ⅱ), qP] all increased first and then decreased. Fv/Fm and Fv/Fo of Vallisneria natans reached the maximum under the condition of 2.0 m, but the maximum of Potamogeton malaianus appeared at 1.5 m. At the same depth, qN of Vallisneria natans was lower than that of Potamogeton malaianus. The change trend of qP was reversed. The maximum Y(Ⅱ) of Vallisneria natans appeared in the range of 1.5-2.0 m water depth, but for Potamogeton malaianus, it appeared at 1.0-1.5 m. Y(NO) showed significant differences with changes of water depths, too high or too low water depth all inhibited plant growth. The relative photosynthetic electron transport rate (ETR) was significantly different between different water depth treatments. The maximum ETR of Vallisneria natans was smaller than that of Potamogeton malaianus which indicates that it has strong resistance to weak light. In summary, under the condition of 1.5-2.0 m water depth, Vallisneria natans has the strongest photosynthetic capacity and can grow the best, and Potamogeton malaianus is the most suitable for growth in 1.0-1.5 m.-
Key words: Vallisneria natans, Potamogeton malaianus, water depth, chlorophyll fluorescence, light-response curve
-
沉水植物是湖泊生态系统的重要初级生产者,对维护湖泊生态系统功能起关键作用(胡振鹏和葛刚,2010)。随着水体污染的日益严重,沉水植物的恢复和重建已成为水域生态学研究的重点内容之一(Ewel,1987)。基质、水深、光照强度、透明度、营养盐等是影响沉水植物生长的重要因素。水深是影响沉水植物在浅水湖泊中能否成功定居并存活的一个重要因子(Valley & Ddrake,2007;王华等,2008)。不同植物在不同水位环境下,生物量和生存能力具有顯著差异(Fraser & Karnezis,2005;Luo & Xie,2009)。研究表明不同水深下苦草的株数、叶片长度、叶片厚度等随水深变化显著,透明度为1.0~1.5 m的条件下,适宜种植水深为0.5~1.0 m(顾燕飞等,2017)。植物生长的好坏可以通过生物量的变化直接表现出来(杨鑫等,2014b),其研究结果显示1.3 m较适宜苦草生长;在透明度较好的水域中1.0~1.4 m的水深范围恢复苦草种群(曹昀等,2014)。认为太湖马来眼子菜的最佳生长水深为0.6~1.2 m,水深较大或较小都会因为光照不足或光损伤等因素抑制其生长和繁殖(翟水晶等,2008)。以上研究表明沉水植物能通过形态和生理响应来适应不同的水深,而目前对于鄱阳湖沉水植物的研究较薄弱,从叶绿素荧光特性方面的差异研究还不够深入。
Kautsky发现叶绿素荧光诱导现象并将其与光合作用联系起来,叶绿素荧光技术能够反映光合系统“内在性”特点(Schreiber et al,1995),已越来越多地应用于植物生理生态研究(胡丰姣等, 2017)。调制荧光仪允许测量所有生理状态下的荧光(包括背景光很强时),使得叶绿素荧光由传统的“黑匣子”(避免环境光)测量走向了野外环境光下测量,水下调制荧光仪(Diving-pam)的出现使得原位测定沉水植物的光合作用成为可能(Yu et al,2010;Hussner et al,2011)。
鄱阳湖作为中国最大的淡水湖,占长江流域9%的面积,具有调节水量,调节气侯,美化环境等多重功能(闵骞等,2009)。鄱阳湖生态系统健康对长江中下游的水环境和水生态具有重要意义(简敏菲等,2015)。2013年第二次鄱阳湖科学考察发现沉水植物分布面积明显下降,20世纪80年代马来眼子菜几乎遍布全湖,现在仅部分蝶形湖和浅水区域小面积分布,鄱阳湖水位低枯是湿地
植被退化的主要原因(胡振鹏等, 2015)。目前鄱阳湖分布最广的沉水植物为苦草,马来眼子菜为伴生种(简敏菲等, 2015)。本研究以苦草和马来眼子菜为研究对象,通过原位测定不同水深下植物叶片的荧光特性,结合生物量探讨两种植物生长的最适宜水深,以期为鄱阳湖水位调控及沉水植物种群的恢复提供科学依据。
1材料与方法
1.1 材料
2016年6月初,苦草和马来眼子菜取自鄱阳湖自然保护区的大湖池。先将长势一致的植物放在塑料桶内(120 cm×80 cm×110 cm)驯养2周,再从驯化植株中选取健康,长势一致的植株,移植到装有鄱阳湖底泥的小桶内。每桶6株苦草,平均每株湿重为(0.86±0.03)g,平均每株有6片叶片,高(11.25±0.34) cm;马来眼子菜采用扦插培养,每桶6株,每株平均湿重为(0.52±0.02) g,平均每株有3片叶片,高(30.12±0.46)cm。
1.2 试验设计
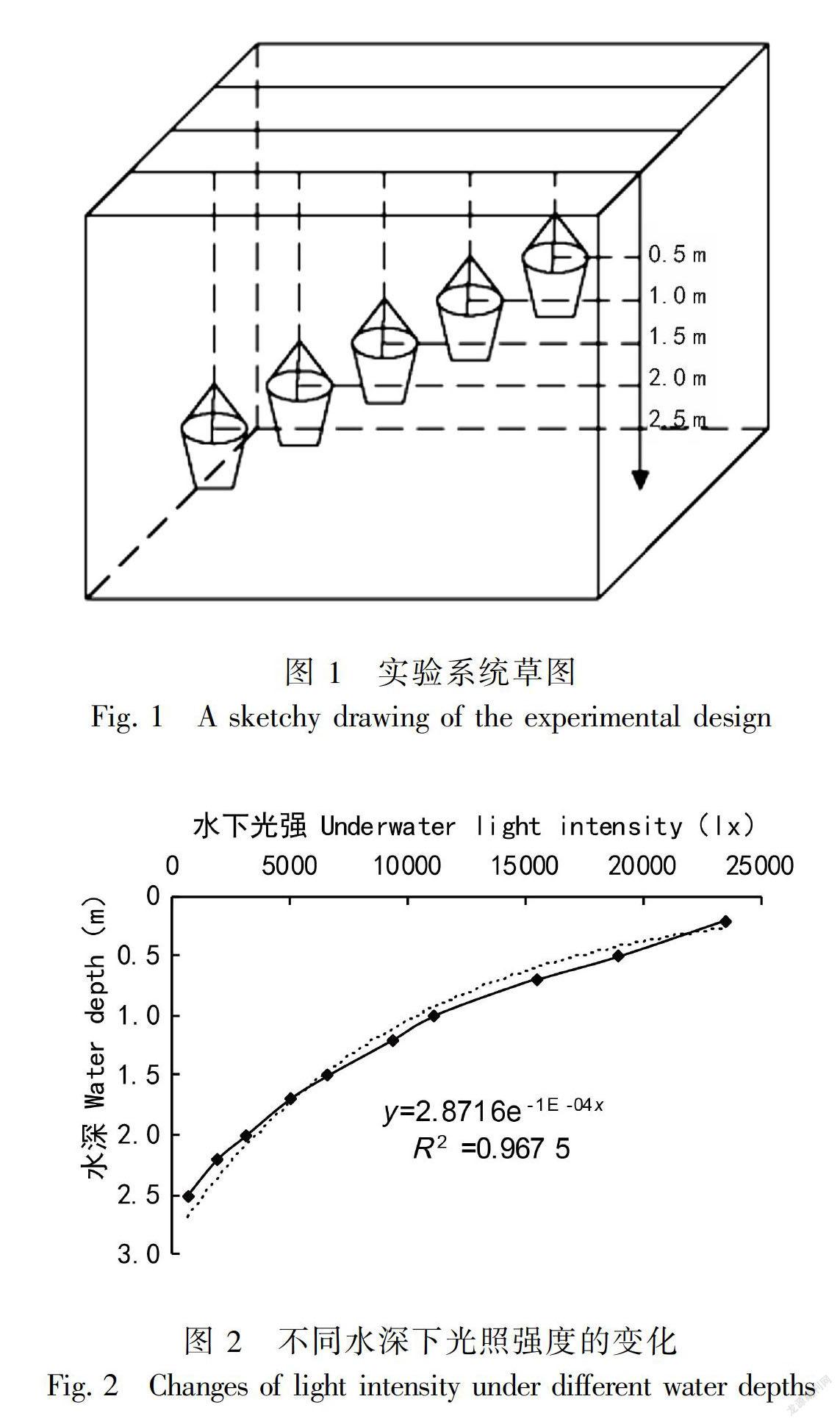
采用室外模拟控制试验,将装有驯化培养苦草和马来眼子菜的小桶悬挂于混凝土浇筑的水池(12.0 m×6.0 m×2.8 m)内,每个梯度做3个重复。水池内的水引自鄱阳湖,水质指标为pH值6.9~7.0、总氮0.52~0.59 mg·L-1、氨氮0.38~0.42 mg·L-1、总磷0.048~0.05 mg·L-1、化学需氧量14.2~14.9 mg·L-1、透明度1.6~1.8 m(每周测1次)。通过吊桶的方式控制水位,整个试验在中国科学院鄱阳湖湿地观测站(星子站)完成。水位变化梯度为0.5、1.0、1.5、2.0、2.5 m,每个梯度设3个重复(图1)。光照强度是影响沉水植物进行光合作用的主要限制因子(吴明丽和李叙勇,2012)。采用水下光照强度的变化规律(比尔定律):Ih=Ioe-kh,式中Io为水面下1 cm处的光照强度(lx);h为水面下深度(cm);k为光衰减系数(王华等,2008)。光在试验水体中的分布可拟合为y=2.8716e-1E-04x。随着水深增加,水下光强衰减显著(图2)。
1.3 生物量测定
用直尺测量活体植物的高;每种水深,取3个吊桶中的全部植株,用滤纸擦掉表面附着物后称其湿重。
1.4 叶绿素荧光参数的测定
培养30 d后,采用德国WALZ公司产的水下调制荧光仪(Diving-pam)测定叶片叶绿素荧光特性。测量前,将暗叶夹夹于叶片,经暗适应20 min 后,打开叶夹,开启检测光,首先测定诱导曲线,得到最小荧光(Fo),最大荧光(Fm),时间约5 min。 PSⅡ的最大光化光效率(Fv/Fm)、有效量子产量[Y(Ⅱ)]、光化学淬灭系数(qP)、非光化学淬灭系数(qN)、非调节性能量耗散量子产额[Y(NO)]等参数值由系统选定模式自动计算生成。再测定快速光曲线,有效辐射强度PAR梯度分别为0、100、200、300、500、700、900、1 100和1 250 μmol·m-2·s-1,时间约2 min。为避免太阳光直射到测量的叶片,测定在上午9:00前结束。
1.5 数据分析
用Excel 2016进行数据处理和图表制作,用SPSS19.0软件对不同水深下苦草和马来眼子菜的叶绿素荧光参数进行单因素方差分析,并用Duncan法进行多重比较分析,显著水平为P<0.05。
2结果与分析
2.1 水深对株高及生物量的影响
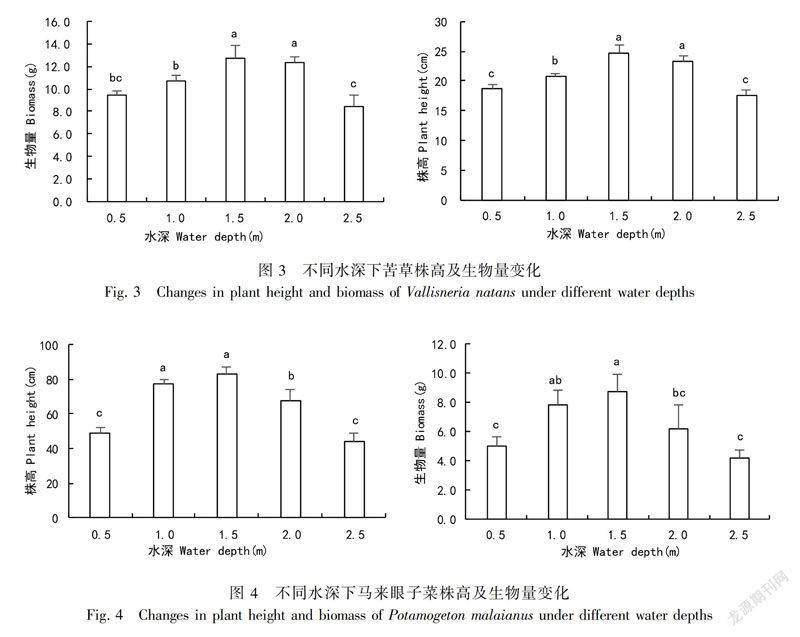
苦草在0.5~1.5 m水深范围内,植株高度随水深的增加显著增加,在1.5 m达到最大值24.73 cm,水深1.5 m和2.0 m变化差异不显著(P>0.05);在2.5 m处,株高仅为17.57 cm,达到最小(图3)。生物量的变化趋势与株高相似,所不同的是水深0.5 m和1.0 m处生物量差异不显著,原因是浅水位苦草新叶生成多,是苦草对水深变化的形态可塑性响应(Yang et al,2004)。
马来眼子菜株高和生物量在0.5~1.5 m水深范围内,随着水深的增加显著增加,在1.5 m达到最大值83.37 cm;1.0 m和1.5 m变化差异不显著。水深1.5 m以后,株高和生物量显著下降,深水位对生物量的积累有抑制作用。
2.2 苦草和马来眼子菜Fo、Fm、Fv/Fm与Fv/Fo对水深的响应
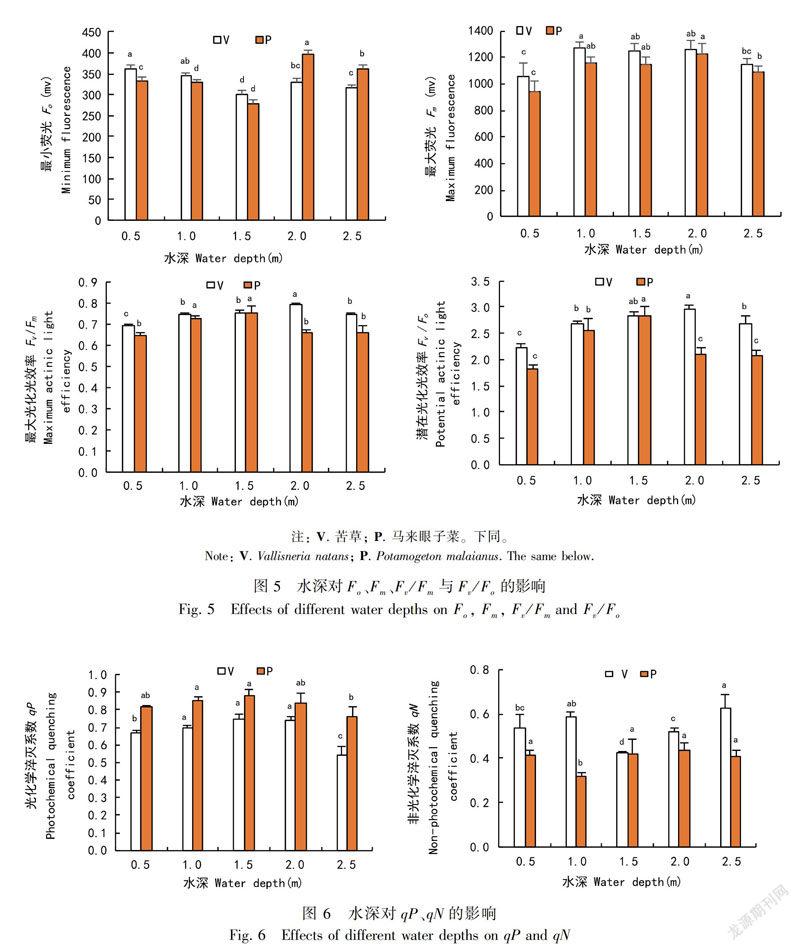
根据实验所设置的5个不同水位,测定苦草和马来眼子菜的Fo、Fm、Fv/Fm及Fv/Fo,分别统计其变化特征及相关显著性,结果如图5所示。
苦草的Fo随水深的增加先降低后增加,水深1.5 m处最低,而在水深2.0~2.5 m之间苦草的Fo差异不显著(P>0.05)。马来眼子菜也出现相似的趋势(图5),不同之处是在2.0~2.5 m水深之间,Fo差异显著。在0.5~1.5 m范围内,苦草Fo大于马来眼子菜;当水深大于2.0 m时,正好相反,表明不同水深下苦草和马来眼子菜PSⅡ反应中心原初开放状态不一。
苦草的Fm在1.0~2.0 m之间差异不显著。随着水深的增加,马来眼子菜的Fm先增加后降低(1.5 m水深除外),最大值出现在2.0 m处。苦草的Fm值均大于马来眼子菜,说明苦草的电子通过PSⅡ时的最大传递潜力较大。随着水深的增加,两种植物的Fv/Fm、Fv/Fo均呈现先增加后降低的趋势,其中苦草最大值出现在2.0 m水深处,而马来眼子菜最大值出现在1.5 m水深处。
综上所述,苦草的Fm、Fv/Fm和Fv/Fo在试验水深范围内均显著高于马来眼子菜,说明苦草PSⅡ原初光能转化效率高于马来眼子菜。
2.3 苦草和马来眼子菜qP、qN对水深的响应
根据实验所设置的5个不同水位,测定苦草和马来眼子菜的qP和qN,分别统计其变化特征及相关显著性,结果如图6所示。
光化學淬灭系数qP是指PSⅡ天线色素吸收的光能用于光化学电子传递的比例,是将捕获的光子能量用于光化学反应的能力指标。苦草的qP在1.0~2.0 m范围内差异不显著(P>0.05),2.5 m处出现大幅下降,马来眼子菜qP最小值也出现在2.5 m,说明植物叶片光化学淬灭受到抑制,这与高氮水抑制挺水植物,qP下降相似(宫兆宁等,2016)。整个水深范围内,苦草的qP均小于马来眼子菜,说明苦草吸收的光能参与电子传递的份额比马来眼子菜低。qN反映植物耗散过剩光能为热的能力(吴昊等,2016),已被证明为检测植物早期胁迫最敏感的参数,与qP呈现相反趋势。相同水深下,马来眼子菜的qN比苦草低,说明其光保护能力比苦草弱。
2.4 苦草和马来眼子菜Y(Ⅱ)、Y(NO)、Y(NPQ)对水深的响应
分别测定上述5个不同水深下苦草和马来眼子菜Y(Ⅱ)、Y(NO)和Y(NPQ),两种植物的Y(Ⅱ)随着水深的增加均呈现先增加后降低的趋势,其中苦草最大值出现在1.5~2.0 m范围内,马来眼子菜的最大值出现在1.0~1.5 m水深处。而苦草的Y(Ⅱ)小于马来眼子菜,说明苦草的实际光合效率较低。
苦草的Y(NO)均表现出显著性差异(P<0.05),最大值出现在<1.0 m和>2.0 m处,说明过高或过低水深植物均已受到损伤。马来眼子菜的Y(NO)未表现出显著性差异。两种植物的Y(NPQ)
均表现出显著性差异(P<0.05),其中苦草的Y(NPQ)大于马来眼子菜,说明相同水深下苦草接受的光强过剩。
2.5 快速光曲线对水深的响应
通过设定有效辐射强度PAR测定5个不同水深下植物叶片的相对光合电子传递速率ETR,拟合出快速光曲线,结果如图8所示。随着PAR增大,相对光合电子传递速率ETR总体上呈现先增大后降低趋势。PAR在小于300 μmol·m-2·s-1范围,表现出快速增长的趋势,而大于此值时,降低较为缓慢。苦草和马来眼子菜的ETR最大值均出现在300 μmol·m-2·s-1,达到最大饱和。苦草ETR峰值是32.6 μmol·m-2·s-1,出现在2.0 m处;而马来眼子菜峰值是35.9 μmol·m-2·s-1,出现在1.5 m水深处,随着有效辐射强度的不断增强,两者都出现光抑制现象。有研究对不同水深下黑藻与苦草光合作用进行比较,苦草的ETR峰值比黑藻低,说明黑藻的光合作用比苦草强(经博翰和袁龙义,2014)。在对玉溪大河五种沉水植物叶绿素荧光特征的比较研究中,ETR最小者说明其耐弱光能力强,适合在底层生长(卢国理等,2017)。在本研究中,苦草的ETR比马来眼子菜低,说明苦草比马来眼子菜耐弱光能力更强,适宜的水深更深。
3讨论
水深是影响水生植物生长和分布的一个重要性限制因子,水深处不仅光强弱(陈正勇等,2011),而且植物与水体间气体交换和营养物质交换改变,植物光合作用也会发生改变(Sculthorpe,1972)。本研究表明,水深显著影响鄱阳湖苦草和马来眼子菜叶绿素荧光特性。
Fo的大小与PSⅡ原始激发色素密度以及反应所处类囊体结构状态及叶绿素含量有关,而与光化学反应无关(陈正勇等,2011)。本研究表明,苦草和马来眼子菜的Fo先降低后略有增加,原因可能是较高水深(>2.0 m)或较低水深(<1.0 m)均会激发色素密度升高和内囊体膜的改变,以适应水深变化。1.5~2.0 m可能促进植物叶片叶绿体类囊体膜上的蛋白复合体活性增强使电子传递和光合磷酸化增强。苦草和马来眼子菜Fv/Fm,Fv/Fo值均出现先增加后降低趋势,本研究结果和杨鑫等(2014a)的一致,其中苦草高于马来眼子菜。说明浅水和深水均会降低苦草和马来眼子菜的原初光能转化效率和PSⅡ反应中心潜在活性,原因可能是深水区(>2.0 m)光照强度减弱,植物叶片接受光强过小,不能满足PSⅡ反应中心捕获激发能而造成Fv/Fm低,其次,可能是深水区CO2含量少,光合作用过程中暗反应阶段CO2供给不足,造成转化效率和反应中心活性低,此还有待进一步研究。浅水区(<1.0 m)可能是强光对苦草和马来眼子菜光合机制造成损害,抑制了PSⅡ反应中心活性,降低光化光效率。另外,在相同水深处苦草光化光效率强于马来眼子菜,原因可能是苦草的光补偿点和饱和点均低于马来眼子菜(苏文华等,2004;陈开宁等,2002),在较小光照下,苦草已进行了光合作用,而马来眼子菜却没有。
本研究苦草和马来眼子菜qP随着水深的增加呈现先增加后降低,浅水区(<1.0 m)qN显著高于1.0~2.0 m水深处,原因可能是浅水区过剩的光能对苦草和马来眼子菜产生了光抑制,而用于光化光电子传递的份额qP不变的情况下,超出了PSⅡ反应中心自我保护机制的自我调节最大限,最终导致实际光合速率的降低;而对于深水区(>2.0 m),苦草和马来眼子菜qP低、qN较高,原因可能是光照弱,水压大,叶片气孔受压迫;CO2含量不足,导致CO2固定的电子少以及光能活性低,天线
图 8快速光曲线对水深的响应-
Fig. 8Responses of rapid light curve under different water depths
---
色素吸收的光能更多的以热能的形式耗散掉,实现光化自我保护。另外,相同水深条件下马来眼子菜Y(Ⅱ)和qP均高于苦草,而qN低于苦草,说明马来眼子菜的光合活性强于苦草,而光保护能力弱于苦草。
PSⅡ反应中心吸收光能后,产生的光量子主要用于三部分转化与耗散,即Y(Ⅱ)、Y(NPQ)和Y(NO),所有的光量子产额始终接近1,即Y(Ⅱ)+Y(NPQ)+Y(NO)=1(钱永强等,2011)。本研究表明,苦草和马来眼子菜Y(Ⅱ)均随着水深的增加呈现先增加后降低,不同水深之间Y(NPQ)和Y(NO)差异显著。原因可能是为适应不同水深下叶片接受的光照强度,致使苦草和马来眼子菜在完成自身对光能的需求外,光合机制还需将不能利用的量子能量以热能耗散的方式分解掉过多光能而使反应中心受伤害程度降至最低,实现植物自我保护。
相对光合电子传递速率ETR是反映实际光强下的表观电子传递和光化光反应导致碳固定的电子传递情况。本研究中,随着PAR的增大,两种植物的ETR均呈现快速上升而后缓慢下降趋势,PAR达到300 μmol·m-2·s-1时,ETR达到最大饱和,这表明PAR超过300 μmol·m-2·s-1时,苦草和马来眼子菜电子传递速率显著受到光抑制,天线色素吸收的过剩光能只能以增加热耗散来减轻过剩能量对植物光系统伤害,使伤害降至最低。无论是马来眼子菜还是苦草,水深在1.0~2.0 m之间时,ETR均是高于其他水深的,苦草2.0 m处最高,马来眼子菜1.5 m处最高,这与生物量测定结果相吻合。这说明不同植物之间对充分利用光能实现自我生存生长不一,是两种植物对环境的适应方式以及光合机构利用光能效率存在的显著性差异。
4结论
本研究结果表明,浅水和深水均会降低苦草和马来眼子菜的原初光能转化效率和PSⅡ反应中心潜在活性。浅水区(<1.0 m)可能是强光对苦草和马来眼子菜光合机制造成损害,抑制了PSⅡ反应中心活性,降低光化光效率。深水区(>2.0 m)光照强度减弱,植物叶片接受光强过小,不能满足PSⅡ反应中心捕获激发能,生物量最小。苦草的最大ETR比马来眼子菜小,说明其耐弱光能力强,适宜在较深水体生长。通过荧光特性分析,水深1.5~2.0 m最适宜苦草生长,水深1.0~1.5 m最适宜马来眼子菜生长。
-
参考文献:-
CAO Y, ZHANG SJ, LIU YY, et al, 2014. Effects of water gradient on seedling growth and biomass of Vallisneria natans [J]. Ecol Environ, 23(8): 1332-1337. [曹昀, 張素娟, 刘燕燕, 等, 2014. 水深梯度对苦莱生长和生物量的影响 [J]. 生态环境学报, 23(8): 1332-1337.]-
CHEN ZY, WANG GX, WU XD, et al, 2011. Ecological adaptability of Potamogeton crispus under different water depths [J]. J Lake Sci, 23(6): 942-948. [陈正勇, 王国祥, 吴晓东, 等, 2011.不同水深条件下菹草(Potamogeton crispus)的适应对策 [J].湖泊科学, 23(6):942-948.] -
CHEN KN, QIANG S, LI WC, et al, 2002. Photosynthetic rate in Potamogeton pectinatus L. and factors of influence [J]. J Lake Sci, 4(14):357-361. [陈开宁, 强胜, 李文朝, 等, 2002. 蓖齿眼子菜的光合速率及影响因素 [J]. 湖泊科学, 4(14): 357-361.]-
EWEL JJ, 1987. Restoration is the ultimate test of ecological theory [J]. Restor Ecol: 31-33.-
FRASER LH, KARNEZIS JP, 2005. A comparative assessment of seedling survival and biomass accumulation for fourteen wetland plant species grown under minor water-depth diffe-rences [J]. Wetlands, 25(3):520-530.-
GONG ZN, FAN YB, LIU H, et al, 2016. Chlorophyll fluore-scence response characteristics of typical emergent plants under different total nitrogen gradient [J]. Chin Bull Bot, 51(5):631-638. [宫兆宁, 范云豹, 刘辉, 等, 2016. 不同水氮梯度下典型挺水植物叶绿素荧光的响应特性 [J].植物学报, 51(5):631-638.]-
GU YF, WANG J, WANG J, et al, 2017. Morphological response and growth strategy of the submerged macrophyte Vallisneria natans under different water depths [J]. J Lake Sci, 29(3):654-661. [顾燕飞, 王俊, 王洁, 等, 2017. 不同水深条件下沉水植物苦草(Vallisneria natans)的形态响应和生长策略 [J]. 湖泊科学, 29(3):654-661.]-
HU FJ, HUANG XH, ZHU F, et al, 2017. Application of chlorophyll fluorescence analysis in environmental stress [J]. Guangxi For Sci, 24(3):381-386. [胡丰姣, 黄鑫浩, 朱凡, 等, 2017. 叶绿素荧光动力学技术在胁迫环境下的研究进展 [J]. 广西林业科学, 46(1):102-106.]-
HU ZP, GE G, 2010. Structure poyang lake wetland plants ecosystem and influence of lake water level for the structure [J]. Res Eviron Yang Bas, 19(6): 597-605. [胡振鹏, 葛刚, 2010. 鄱阳湖湿地植物生态系统结构及湖水位对其影响研究 [J]. 长江流域资源与环境, 19(6): 597-605.]-
HU ZP, GE G, LIU CL, 2015. Cause analysis and early warning of the poyang lake wetland vegetation degradation [J]. Res Environ Yang Bas, 24(3):381-386. [胡振鹏, 葛刚, 刘成林, 2015. 鄱阳湖湿地植被退化原因分析及其预警 [J]. 长江流域资源与环境, 24(3):381-386.]-
HUSSNER A, HOFSTRA D, JAHNS P, 2011. Diurnal courses of net photosynthesis and photosystem Ⅱ quantum efficiency of submerged Lagarosiphon major under natural light conditions [J]. Flora, 206(10):904-909. -
JIAN MF, JIAN MF, LI LY, et al, 2015. Distribution pattern of submerged plants in typical wetlands of poyang lake and its influencing factors of water environment [J]. Res Environ Yang Bas, 24(5):765-771. [簡敏菲, 简美锋, 李玲玉, 等, 2015. 鄱阳湖典型湿地沉水植物的分布格局及其水环境影响因子 [J]. 长江流域资源与环境, 24(5): 765-771.]-
JING BH, YUAN LY, 2014. The comparative study on photosynthesis of Hydrilla verticillata in the different depth [J]. J Jiangxi Norm Univ (Nat Sci Ed), 38(6):645-649. [经博翰, 袁龙义, 2014. 黑藻与苦草在不同水深下光合作用的比较研究 [J]. 江西师范大学学报(自然科学版), 38(6):645-649.]-
SCHREIBER U, BILGER W, NEUBAUER C, 1995. Chlorophyll fluorescence as nonintrusive indicator for rapid assessment of in vivo photosynthesis [J]. Ecol Stud: 49-70.-
LUO W, XIE Y, 2009. Growth and morphological responses to water level and nutrient supply in three emergent macrophyte species [J]. Hydrobiologia, 624(1):151-160.-
LU GL, ZHOU YQ, LI SY, et al, 2017. Comparative of chlorophyll fluorescence characteristics of five submerged macrophytes in Yuxi River [J]. Environ Sci Surv, 36(2):46-50. [卢国理, 周元清, 李淑英, 等, 2017. 玉溪大河 5 种沉水植物的叶绿素荧光特征比较研究 [J]. 环境科学导刊, 36(2):46-50.]-
MIN Q, TAN GL, JIN YW, 2009. Main issues and regulation measures of Poyang Lake ecosystem [J]. Chin Wat Res, (11):44-47. [闵骞, 谭国良, 金叶文, 2009. 鄱阳湖生态系统主要问题与调控对策 [J]. 中国水利, (11): 44-47.]-
QIAN YQ, ZHOU XX, HAN L, et al, 2011. Rapid light-response curves of PS Ⅱ chlorophyll fluorescence parameters in leaves of Salix leucopithecia subjected to cadmium-ion stress [J]. J Lake Sci, 4(14):357-361. [钱永强, 周晓星, 韩蕾, 等, 2011. Cd2+胁迫对银牙柳PSⅡ叶绿素荧光光响应曲线的影响 [J]. 湖泊科学, 4(14):357-361.]-
QU SJ, HU WP, DENG JC, et al, 2008. Effects of different water depths and sediments on Potamogeton malaianus in lake taihu [J]. Acta Ecol Sin, 28(7):3035-3041. [翟水晶, 胡维平, 邓建才, 等, 2008. 不同水深和底质对太湖马来眼子菜 (Potamogeton malaianus) 生长的影响 [J]. 生态学报, 28(7):3035-3041.]-
SCHREIBER U, BILGER W, NEUBAUER C, 1995. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis [M]//Ecophysiology of photosynthesis. Berlin, Heidelberg: Springer: 49-70. -
SCULTHORPE CD, 1972. The biology of aquatic vascular plants [J]. J Ecol, 56 (3):915.-
SU WH, ZHANG GF, ZHANG YS, et al, 2004. The photosynthetic characteristics of five submerged aquatic plants [J]. Acta Hydrobiol Sin, 28(4):391-396. [蘇文华, 张光飞, 张云孙, 等, 2004. 5种沉水植物的光合特征 [J]. 水生生物学报, 28(4):391-396.] -
VALLEY RD, DRAKE MT, 2007. What does resilience of a clear-water state in lakes mean for the spatial heterogeneity of submersed macrophyte biovolume? [J]. Aquat Bot, 87(4):307-319. -
WANG H, PANG Y, LIU SB, et al, 2008. Research progression influencing of environmental factors on the growth of submersed macrophytes [J]. Acta Ecol Sin, 28(8):3958-3968. [王华, 逄勇, 刘申宝, 等, 2008. 沉水植物生长影响因子研究进展 [J]. 生态学报, 28(8):3958-3968.]-
WU ML, LI XY, 2012. Research progress on influencing of light attenuation and the associated environmental factors on the growth of submersed aquatic vegetation [J]. Acta Ecol Sin, 32(22):7202-7212. [吴明丽, 李叙勇, 2012. 光衰减及其相关环境因子对沉水植物生长影响研究进展 [J]. 生态学报, 32(22):7202-7212 ]-
WU H, GAO Y, DU ME, et al, 2016. Characteristics of chlorophyll fluorescence parameters in Forsythia suspensa (Thunb.) vahl under Nacl stress [J]. N Gard, (7):55-60. [吴昊, 高永, 杜美娥, 等, 2016. 盐胁迫对连翘叶绿素荧光参数的影响 [J]. 北方园艺, (7):55-60.]-
YU H, YE C, SONG X, et al, 2010. Comparative analysis of growth and physio-biochemical responses of Hydrilla verticillata to different sediments in freshwater microcosms [J]. Ecol Eng, 36(10):1285-1289. -
YANG X, SUN SY, BAI X, et al, 2014a. Influences of water depth gradient on photosynthetic fluorescence characteristics of Vallisneria natans [J]. J Lake Sci, 26(6):879-886. [杨鑫, 孙淑雲, 柏祥, 等, 2014a. 水深梯度对苦草(Vallisneria natans)光合荧光特性的影响 [J]. 湖泊科学, 26(6):879-886.]-
YANG X, ZHANG QC, SUN SY, et al, 2014b. Effects of water depth on the growth of Vallisneria natans and photosynthetic system Ⅱ photochemical characteristics of the leaves [J]. Chin J Appl Ecol, 25(6):1623-1631. [楊鑫, 张启超, 孙淑雲, 等, 2014b. 水深对苦草生长及叶片 PSⅡ 光化学特性的影响 [J]. 应用生态学报, 25(6):1623-1631.]-
YANG Y, YU D, XIE YH, et al, 2004. Phenotypic plasticity of two submerged plants in response to flooding [J]. J Freshw Ecol, 19: 69-76.
猜你喜欢
软件导刊(2018年5期)2018-06-21
小学生作文选刊·中高年级版(2017年10期)2017-11-07
科技视界(2017年4期)2017-05-22
健康之路(医药研究)(2016年5期)2016-12-10
发明与创新·中学生(2016年12期)2016-09-10
河北渔业(2015年11期)2015-11-18
数学大王·中高年级(2014年1期)2015-02-12
江苏农业科学(2014年9期)2014-11-15
中小企业管理与科技·下旬刊(2009年9期)2009-03-08
