
玉米株高主效QTL定位研究综述
2018-10-09 08:53刘忠祥
甘肃农业科技 2018年9期
刘忠祥
(甘肃省农业科学院作物研究所,甘肃 兰州 730070)
玉米是重要的粮食、饲料、工业原料和生物质能源作物,在解决粮食安全和能源安全这两个全球性问题当中扮演着极其重要的角色。株高是玉米的重要农艺性状,与籽粒产量、生物学产量、抗倒伏性密切相关。随着分子生物学的发展,对控制玉米株高的QTL基因研究成为玉米育种工作的热点。在玉米诸多株型构成成分中,茎秆高度尤为重要,它与产量密切相关,解析玉米株高的遗传基础,将为如何合理利用玉米株高开展株型育种,乃至提高玉米产量都具有重要意义。适当降低株高、增加种植密度,提高抗倒伏能力可以增加产量;增加株高也可以增加玉米生物学产量,提高玉米在生物能源和青贮饲料中的应用价值。近年来,控制玉米株高性状的QTL基因很受育种者关注,在基因定位、克隆和功能研究方面都取得了较大进展,对更深入研究玉米株高QTL基因定位、克隆和功能研究具有重要指导意义。笔者总结了玉米株高主效QTL定位研究进展及与株高相关基因的功能与响应途径,供玉米遗传育种工作者参考。
1 作物株高与产量高度相关
农业生产史上,农作物整体株高的降低对世界农业产生了深远的影响。20世纪70年代,在小麦和水稻生产中正是由于推广种植株高较矮的新品种爆发了绿色革命,使世界小麦和水稻产量大幅度增长。学者和种植者对矮小品种的偏爱,是由于它们能抵御大风、降水、高密度种植导致的倒伏,同时矮小作物品种也能提高肥料的利用率[1-3]。
玉米株高与籽粒产量、生物学产量、抗倒伏性密切相关,是一个重要的产量相关性状。增加玉米种植密度是当前提高玉米产量的主要方式。但随着种植密度的增加,玉米植株倒伏的风险也随之增大,降低株高是避免倒伏的主要育种策略之一。随着人们对玉米的需求量越来越大,玉米单位面积产量也逐步增长。在美国,平均玉米籽粒产量从1930年的1 500 kg/hm2,增加到现在的8 500 kg/hm2,增加了4倍多,大约每年玉米产量增加109 kg/hm2[4]。在中国,玉米籽粒产量从新中国建立时的1 060 kg/hm2提高到2008年的5 290 kg/hm2,单位面积产量提高了近4倍[5]。同时,玉米种植密度也在不断增加。中国玉米单产的增长主要是由于种植密度的增加。同样在美国洛瓦州,相比于1965年,2008年该州玉米种植密度增长了96%,平均玉米产量增长了110%,可见该州玉米单位面积产量增加的原因主要是由于种植密度的增加,而不是玉米单株产量的增加[5-6]。目前增加种植密度是我国及世界增加玉米产量所采取的关键措施之一,但种植密度的增加,往往会出现倒伏现象,带来更大程度的减产,严重的可能减产30%~50%[7]。这是由于随着种植密度的增加,田间植株竞争性生长,使得玉米植株茎秆过分伸长而变的细弱,同时导致植株的雌穗高度增加,植株整体重心升高,在遭遇大风和暴雨时,常常出现玉米大范围倒伏[8]。也有研究显示,玉米倒伏出现的可能与种植密度的增加高度相关[7],而推广种植植株高度较矮的新品种不仅有利于增加种植密度,也能有效减少倒伏的出现。所以,选育适当高度的玉米新品种有利于玉米产量的增加,而剖析控制玉米植株高度的遗传因素也有重要意义。
2 玉米株高QTL研究进展
作物株高表现数量变异。在玉米中,株高是遗传力最高(预计超过90%)且容易测量的性状之一。由于株高的这个特性,从Mendel的杂交试验以来[9],不同研究机构对玉米株高进行了大量的遗传研究,同时玉米株高也作为剖析QTL遗传基础的模式性状。
早在1987年,Edwards等[10]利用两个F2群体对玉米株高QTL进行了鉴定,共检测到21个与株高连锁的标记位点。随后,Beavis等[11]利用4个群体共鉴定出16个株高QTL,其中12个与已知的株高突变体所在位点接近,说明影响数量性状的位点与导致突变产生的质量位点可能是同一基因,而Berke等[12]报道了在玉米染色体上有11个区域与株高密切相关。随着分子标记技术的发展,到目前为止已经鉴定到大量的株高QTL,同时对主效QTL进行了图位克隆。张志明等[13]指出,目前已定位的株高QTL多达280个,从第1到第10染色体上已检测的个数分别为35,14,34,16,22,18,22,23,20,17,另有 60多个未提供染色体信息[14-15]。Lima等[16]利用热带种质玉米F2群体,在第1、2、4、5染色体上总共定位到6个株高QTL。Tang等[17]选取综3和87-1为亲本发展了包含294个系的RIL群体,并利用该群体鉴定到6个株高相关QTL和6个平均节间长的QTL,其中株高QTL分布在第1、2、3、5号染色体上,6个平均节间长的QTL中有4个与定位到的株高QTL位于相同染色体区域。杨晓军等[18]利用一个包含397个家系的分离群体,共检测到21个株高相关的QTL,其中第1、2、3和5四条染色体上分布的株高QTL较多,并推测第5染色体的bin 5.05-5.07区域可能存在控制株高和穗位高度的主效QTL;严建兵等[19]利用玉米自交系综3和87-1构建的266个F2群体对玉米的五个不同生长时期进行了株高QTL定位,共定位到8个QTL,其中3个QTL在不同时期都能稳定检测到,但同一QTL在不同时期所解释的表型变异差别较大,说明株高QTL随着植株的生长表达会发生变化。
相对于初级作图群体,利用导入系群体检测株高QTL,由于消除了遗传背景的干扰,检测效力将会显著提高,同时可以检测基因间的互作。Salvi等[20]用 Gaspé Flint作为供体亲本,B73 为受体亲本,构建了含有75个系的导入系群体。利用该导入系进行株高及其他农艺性状的定位,其中检测到的4个株高QTL分布于第1、3、8、10染色体上。Bai等[21]利用HB522为供体,综3为受体,发展了一套包含98个系的片段代换系群体,并利用该群体鉴定到9个QTL,分别位于1、2、3、5、6染色体,其中位于第3染色体上的株高QTL,在之前许多研究中也被多次被检测到。利用同一导入系群体,Teng等[22]在该区域克隆到一个控制株高的主效QTL。Salvi等[23-24]利用NIL群体在第8染色体上定位到的一个控制玉米花期同时对株高有影响的QTL,并在2007年最终对该基因实现了克隆,发现Vgt1充当顺式作用因子调控下游基因表达。
除了连锁作图的方法,关联分析也用于鉴定株高QTL的鉴定。Weng等[25]使用284个自交系材料,利用平均分布在染色体上的超过55 000个SNP标记进行基因型分析,共检测到204个与株高显著关联的SNP位点,其中1.11 bin、2.09 bin、3.02 bin、4.05 bin、5.05 bin、5.06 bin、6.03 bin、6.04 bin、6.05 bin、9.07 bin和10.03 bin是热点区域。Peiffer等[26-28]利用NAM群体,采用联合作图方法,定位到89个与株高显著关联的标记,并利用全基因组关联的方法,发现277个与株高显著关联的位点,大部分关联位点与联合作图方法得到的结果一致。其中在第九染色体长臂上有7个与已定位QTL显著关联的位点,同时在对NCRPIS(North Central Regional Plant Introduction Station)的全基因组关联分析中,鉴定到213个与株高显著关联的位点,但与NAM群体关联分析得到的结果重叠的较少。从以上研究可以看到,玉米株高受到强烈的遗传控制,并且有着高度多基因的遗传结构。
3 与株高相关基因的功能与响应途径
Emerson[29]最早报道了关于玉米矮化突变体,该突变体表现出植株矮小、叶片变短变宽颜色深绿、雄穗变粗且分支减少等特征。目前,已鉴定到超过40个由不同基因突变导致的玉米株高突变体[30]。在玉米中已克隆到了多个株高相关基因,这些基因大部分是通过突变体分析克隆得到。
目前已克隆的许多株高相关基因都涉及到赤霉素(GA)和油菜素甾醇(BR)的合成、运输和信号转导途径,此外,生长素(IAA)也被发现参与株高调控。除这些植物激素通过影响节间伸长来影响株高外,植物株高的改变还受到其他基因的调节。
3.1 赤霉素生物合成和信号传导相关基因
赤霉素(GA)是一类广泛存在的四环双帖类植物激素,在植物的生长发育过程中扮演着重要的作用,包括种子萌发、下胚轴伸长、叶片延展、开花时间、花和果实发育及成熟等,而其在茎秆伸长方面起着尤为重要的作用[31-34]。GA是最早由黑泽英一在研究水稻时发现,并在1935年由薮田贞治郎等分离出这种活性物质,并命名为赤霉素[35]。目前已在不同生物中已发现了136种结构明确的GA[36]。然而,这些GA中只有很小一部分具有生物活性,如GA1、GA3、GA4、GA7等,其余绝大部分是GA合成途径中的中间产物或活性GA分解的产物[37]。
在拟南芥和水稻中,已经对GA相关的突变体进行了大量的研究,GA生物合成和信号转导相关基因均被发现参与植株高度的调控。GA生物合成相关基因突变引起的表型改变在外源施用GA后可以回复到野生表型,为GA敏感型,这些基因通常编码GA合成途径中的一些酶,突变体通常表现为由于节间长度减少导致的株高降低、叶片变的短且宽、雄穗分支数减少、雌穗上有花药的形成等特征。而GA信号转导相关基因突变引起的表型改变在外源施用GA后不能回复到野性型表型,为GA不敏感型,突变体表现为矮化或植株细长等特征。
在玉米中,已经鉴别到5个GA敏感型突变体,即d1、d2、d3、d5和another ear 1(an1)。这些基因都是隐性突变,其中d1(ZmGA3ox2)、d3、an1已被克隆出来[38-40]。d1突变体表现为通常的矮化、GA响应表型,该基因编码GA30酶,这种酶是GA合成途径的重要酶,催化生物活性GA合成的最后一步[22];An1突变体表现为植株极端矮化、生育期延长、雌穗上形成完全花等特征,该基因同样编码一种酶,这种酶与GA合成途径中第一个四环中间产物贝壳杉烯的合成有重要作用[41]。引起d3突变的基因编码细胞色素P450蛋白,该蛋白涉及到GA合成起初阶段GA12向GA53的转化[42]。在水稻中,导致植株矮化的基因sd-1(semidwarf-1),也就是著名的水稻绿色革命基因,也是GA合成途径的关键基因。该基因编码GA合成途径中的GA20氧化酶(OsGA20ox2),该基因的突变使水稻栽培种变矮、茎秆变粗。另一GA合成途径相关基因d18编码GA羟化酶(Os-GA3ox2)[43]。
GA信号传导途径中的基因变异同样也会导致植株高度的改变。近年来,随着拟南芥和水稻功能基因组学的发展,多个GA信号传导途径的调控因子已经被鉴定,并且其相对的基因已经被克隆,包括拟南芥中的SPY(SPINDLY)、gai(GA-insensitive)和RGA(repressor of gal-3)、玉米中的 D8(dwarf 8) 和D9(dwarf 9)、大麦的 SLN1(SLENDER1) 和水稻中的 slr1 (slender rice1)、dwarf1、OsGAI,以及小麦的 Reduced height(Rht-B1和Rht-D1)基因[44-50]。这些突变体有2种表型特征,一种是植株细长,如水稻slender突变体、大麦slender突变体和拟南芥spindly,一般为隐性突变;另一种是常见的GA不敏感矮化突变表型,一般为显性突变,如D8和D9、GID1基因、小麦Rht。对于植株瘦长突变体,植株一直处于GA响应激活状态,而矮化植株则表现出对GA信号识别和传导缺陷。黄先忠等[51]研究发现,这些克隆到的GA信号传递关键元件同源性很高(图1),都在N端部位存在一个DELLA结构,均为DELLA蛋白家族成员。DELLA是细胞核内负调节赤霉素信号蛋白,在高等植株中普遍被发现,是赤霉素信号传导的关键因素。DELLA蛋白在N末端的两个保守结构域DELLA和VHYNP是GA信号感知结构域。D8、D9这类对GA不敏感的突变体,突变基因编码的蛋白都是在DELLA结构处发生了改变,而对GA敏感的突变体大都是由于基因3'端的碱基突变形成终止密码所引起的翻译提前终止造成的。如玉米GA不敏感突变体d8的三个显性矮化等位基因D8-1、D8-MPL、D8-2023,这3个等位基因的突变都影响到所编码蛋白的N末端结构。其中D8-Mpl编码的蛋白缺少开始的105个氨基酸,其中包括DELLA domain;D8-2023缺少TVHYNP结构域的12个氨基酸;D8-1编码的蛋白中D55被1个谷氨酸替换,而且其后的4个氨基酸缺失[52]。根据DELLA蛋白结构与功能的关系,在克隆出水稻细长突变体slr后,利用这个基因构建了一个显性等位基因。这个显性基因编码的蛋白在DELLA motif有17个氨基酸的删除,被导入这个基因的水稻表现出矮化和GA不明感表型。DELLA蛋白N端的重要作用可能在于与可溶性GA受体GID1的互作有关,在水稻和拟南芥中已经证实了DELLA蛋白与GID1能互作,SLr1-d突变基因编码的DELLA蛋白与GID互作降低,由于这种互作的降低,导致GA依赖的SLr1-d蛋白降解变慢,导致突变体矮化[49]。
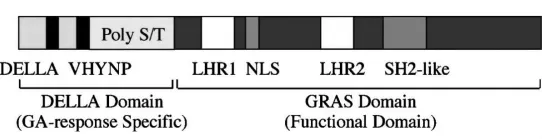
图1 DELLA蛋白家族的结构示意
3.2 油菜素甾醇(BR)生物合成和信号传导相关基因
油菜素甾醇(brassinosteroid,BR)是在1970年被发现,在1979年被分离出来,是已发现的这类物质中活性最强的分子。BR在植物各个发育阶段和多种发育过程中都有重要作用,其主要功能是促进细胞伸长、植物暗形态建成、胁迫响应等,其结构与动物中性别决定的类固醇结构相似,在BR合成或者是信号传导途径有缺陷的突变体表现出不同程度的矮化,在玉米中BR同样影响性别决定[53]。
在玉米中发现的突变体brd1、na1都涉及到BR合成途径,纯合的brd1突变体茎节间不伸长,在黑暗中生长时没有黄化反应,施用外源激素能在部分程度上恢复突变表型,同时突变体在叶和花等性状上也有变化,表现为叶片皱缩,雄穗雌性化,不能产生种子,该基因编码催化BR合成最后阶段的酶。na1是一个“tassel seed”(ts)突变体,该突变体株高和植株整体构型都有改变。株高的改变是由于茎节长度的变化导致,茎节数没有变化,与其他BRs合成缺陷突变体相似,植株暗形态发生改变,中胚轴伸长受损,同时突变植株雄蕊雌性化,说明BRs不仅影响株高,在促进雄蕊发育的重要作用,该基因编码BRs合成途径中的一种酶[41,54]。依据同源克隆的方法,在玉米中克隆了与拟南芥同源的BR合成相关基因Zmdwf4和ZmDWF1。拟南芥中DWF1/DIM基因编码BRs合成途径的重要的酶,ZmDWF1是玉米中与其同源的基因,该基因编码与FAD结合蛋白结构非常相似的蛋白,在ZmDWF1的RNA干扰转基因植株中,由于茎细胞在轴向方向伸长有缺陷,转基因植株在不同程度上表现矮化现象;拟南芥中的另一个基因DWF4编码BR合成途径中的一种羟基酶,这种酶催化BR合成过程中C27-C29甾醇的羟基化,突变植株表现出矮化、叶片变短且颜色变深、角果变短、雄性不育,ZmDWF4为其在玉米中的同源基因,在拟南芥dwf4的转基因植株中ZmDWF4能恢复突变体矮化表型,且能恢复雄性不育[55-56]。水稻中影响到BR生物合成和信号传导的突变基因均被鉴定到,OsDwarf2、OsDwarf11编码BR生物合成过程的相关酶,而d61在BR信号传导途径有缺陷[57-58]。
GAs和BRs是影响株高的主要的激素,在玉米中生长素(IAA)同样参与株高的调控。玉米中的brachytic2突变体表现出植株下部节间缩短紧凑,而植株上部表现正常,植株整体高度降低,研究发现该基因是通过影响茎秆居间分生组织生长素的极性运输来影响株高[30]。另一突变体vanishing tassel 2同样表现出矮化的特征,该基因编码生长素合成过程中的色氨酸转移酶,该酶催化色氨酸向3-吲哚丙酮酸的转化[59]。
除了激素以外,株高也被一些影响细胞分裂、伸长和合成酶基因所影响。玉米矮化突变体d 2003表现节间数目增加、节间长度缩短、节间细胞数目减少,研究证实突变体中的赤霉素、生长素、细胞分裂素信号传导途径表现正常,其植株矮化的原因在于Viviparous8(Vp8)基因的编码区由于单碱基的插入而导致翻译的提前终止[60]。在水稻中同样发现编码CCT domain蛋白(GHD7)和蔗糖磷酸合成酶的基因(SPS)与株高有关[61-62]。
4 展望
随着现代农业的发展和玉米机收籽粒的不断普及推广,对玉米株高和茎秆弹性提出了更高要求,不但要求茎秆直立不倒,而且要求茎秆富有弹性,以适应机械化收获的需要。玉米株高是由多基因控制的数量性状,大部分数量性状位点(QTL)效应值较小,只有少数QTL表现为主效但易受环境的影响[63-64]。虽然已经报道了许多玉米株高QTL,但由于作图群体、分析方法以及环境条件的差异,往往会造成QTL定位区间过大、区间重叠或多环境条件下不能稳定表达等现象。因此,对控制玉米株高的QTL/基因研究将成为玉米育种工作的热点,发掘、鉴定和利用新的株高基因成为玉米育种中备受重视的研究内容。随着分子技术的发展,开展玉米株高基因定位、克隆和功能研究,阐明其分子机理,对玉米育种和生产具有重要指导意义。
猜你喜欢
河北果树(2021年4期)2021-12-02
农业科技通讯(2021年1期)2021-03-06
河北果树(2020年1期)2020-02-09
农村百事通(2019年17期)2019-10-08
中国食品学报(2019年12期)2019-01-13
老友(2017年7期)2017-08-22
上海农业学报(2017年3期)2017-04-10
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
