
β-木糖苷酶的研究进展
2018-11-03 02:49汤勇,蔡俊*
中国酿造 2018年10期
汤 勇,蔡 俊*
(湖北工业大学 发酵工程教育部重点实验室 工业发酵湖北省协同创新中心 工业微生物湖北省重点实验室,湖北 武汉 430068)
植物细胞壁的横向结构主要由半纤维素和木质素结合而成。β-1,4-木聚糖是木质纤维素生物质中含量最多的半纤维素组分。这种复杂的多糖的基本骨架由β-1,4糖苷键相连的D-吡喃木糖组成,并且含有不同的侧链取代基,最常见的取代基为乙酰基、阿拉伯糖和葡萄糖醛酸。侧链的数量和组成取决于木聚糖来源,例如硬木木聚糖的侧链包括葡萄糖醛酸基和乙酰基,如图1所示,而软木木聚糖的侧链包括葡萄糖醛酸基和阿拉伯呋喃糖基,如图2所示。由于植物木聚糖聚合度和取代侧链的不同,完全降解需要一套木聚糖水解酶系,酶系中各种酶的作用位点见图3,主要包括β-1,4-木聚糖酶和β-木糖苷酶(β-xylosidase,β-xyl)[1]。其中内切β-1,4-木聚糖酶破坏木聚糖骨架成更短的可溶性低聚木糖;β-木糖苷酶水解可溶性低聚木糖和木二糖的非还原末端释放木糖。该催化作用还消除了内切木聚糖酶的终产物抑制作用[2]。
1 β-木糖苷酶的来源
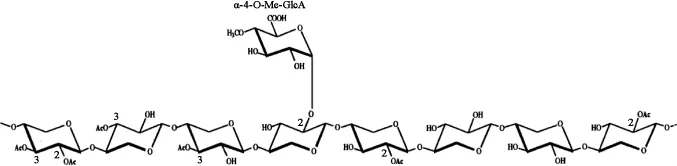
图1 硬木木聚糖的结构Fig.1 Structure of the hardwood xylan
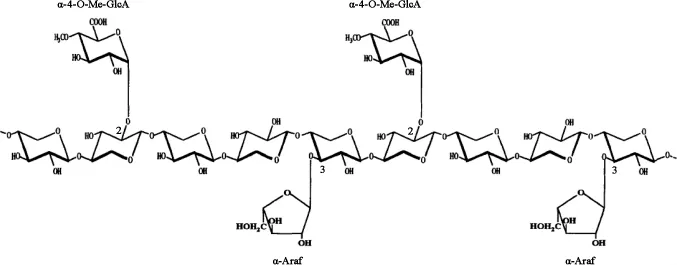
图2 软木木聚糖的结构Fig.2 Structure of the softwood xylan
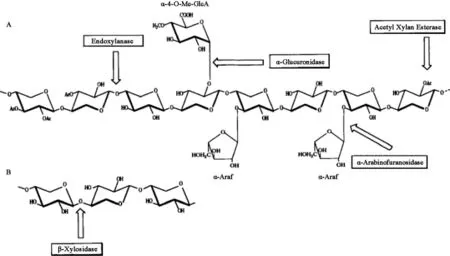
图3 木聚糖降解酶系参与水解木聚糖(A)与β-木糖苷酶水解低聚木糖(B)Fig.3 The xylanolytic enzymes involved in the degradation of xylan(A)and hydrolysis of xylooligosaccharide by β-xylosidase(B)
β-木糖苷酶在自然界中广泛存在,常见于细菌和真菌之中,真菌中又多为丝状真菌,而极少见于酵母菌中。MHETRAS N等[3]筛选得到一株产β-木糖苷酶酵母菌Pseudozyma hubeiensis NCIM 3574。虽然自然界中存在着大量产β-木糖苷酶的微生物,但其所产β-木糖苷酶的酶活和酶学特性不够理想,所以近十几年来,人们致力于通过基因工程的手段来解决上述问题。例如SHI H等[4]成功将Thermotoga thermarum DSM 5069中编码表达β-木糖苷酶的基因导入大肠杆菌(Escherichia coli)BL21中表达。该重组表达的β-木糖苷酶具有更高的活性、耐热性和木糖耐受性。
2 β-木糖苷酶的分类
由于β-木糖苷酶对不同底物的活性是不同的,所以将这些酶根据底物特异性来分类是很困难的。迄今为止,根据碳水化合物活性酶数据库(CAZy),β-木糖苷酶分为糖苷水解酶家族(glycoside hydrolases,GHs)3,30,39,43,52和54,如表1所示[5]。该家族的分类取决于酶所使用的底物、氨基酸序列相似性、三维结构和作用机制。已知丝状真菌的木糖苷酶只属于第3、第43和第54家族,如表1中第3家族中细菌木糖苷酶占89%,第43家族中细菌木糖苷酶占92%。
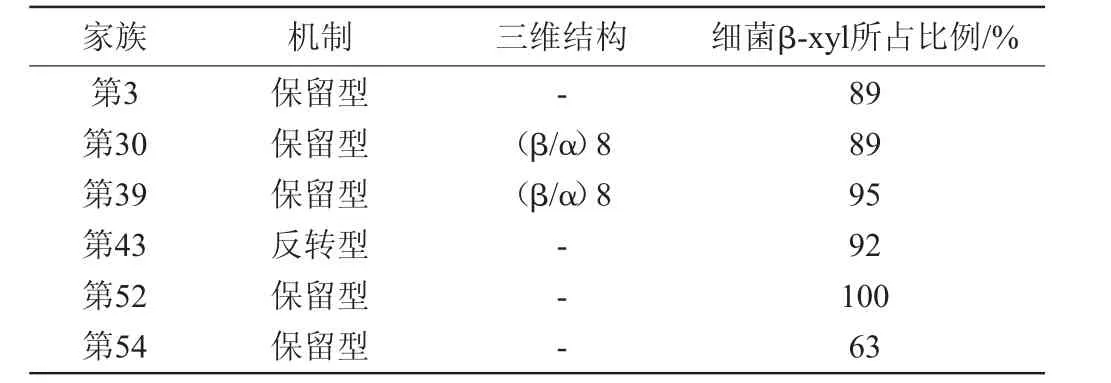
表1 CAZY数据库的β-木糖苷酶家族分类Table 1 β-xylosidase family classification according to the CAZY database
3 β-木糖苷酶的分泌部位
与胞外分泌的木聚糖酶相反,不同微生物和不同培养条件下,β-木糖苷酶可以是胞外型或膜结合型。如绝大多数细菌和酵母所产β-木糖苷酶被认为是以可溶形式分泌于细胞质中。在浅白隐球酵母(Cryptococcusalbidus)中,木二糖和木三糖通过β-木糖苷通透酶系统进入细胞,然后由β-木糖苷酶转化为木糖[6]。在这些微生物中,低聚木糖的水解必须在细胞内完成。相反,真菌所产β-木糖苷酶可分为胞内型(如Streptowycestherwoviolaceus OPC-520)、胞外型(如Aureoba sidiumpullulans)和膜结合型(如Streptomyces sp.EC 10)[7]。某些丝状真菌β-木糖苷酶早期与菌丝体结合,然后通过分泌或细胞裂解释放到胞外。大多数纯化后的真菌β-木糖苷酶分子质量>100 kDa,少数分子质量较低。从这些微生物纯化得到的β-木糖苷酶主要以单体和二聚体的形式存在。
4 β-木糖苷酶的特性
不同微生物来源的β-木糖苷酶的最适pH和最适温度存在差异见表2。
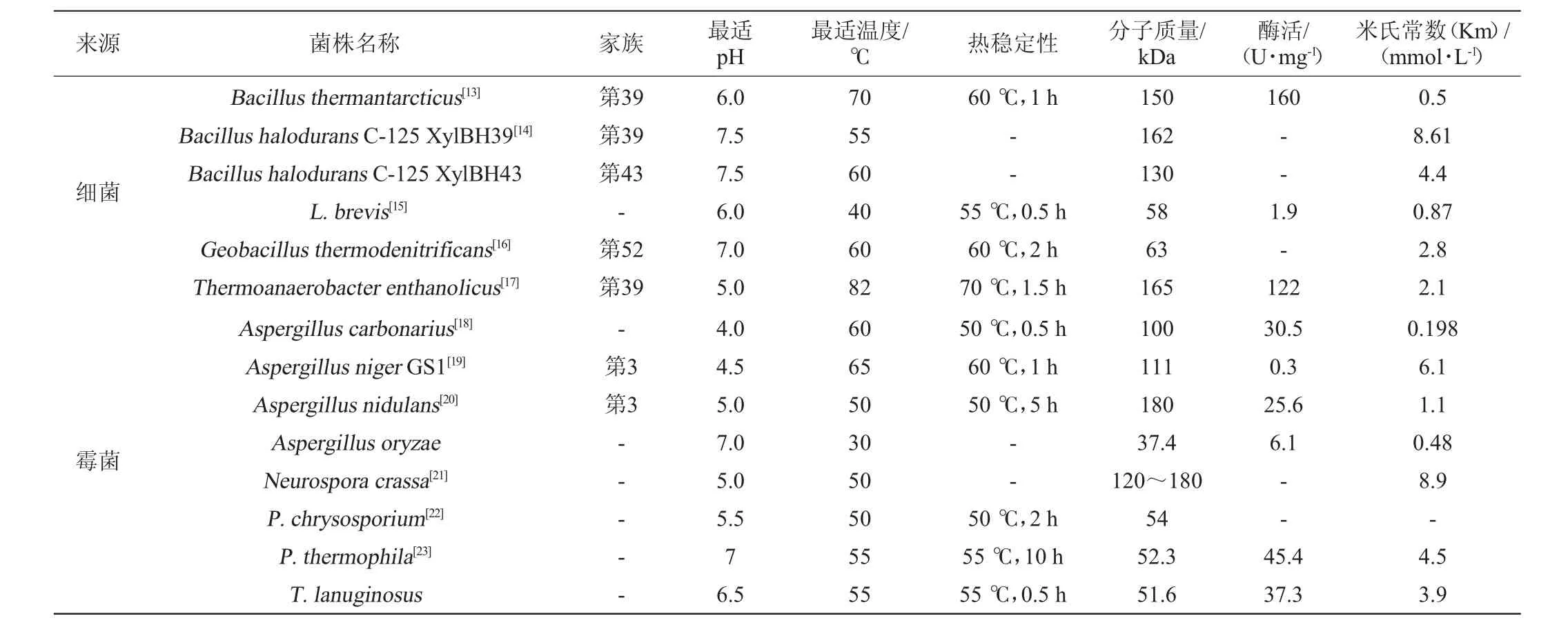
表2 不同微生物来源的β-木糖苷酶的物理化学性质Table 2 Physicochemical properties of β-xylosidase from different microorganism
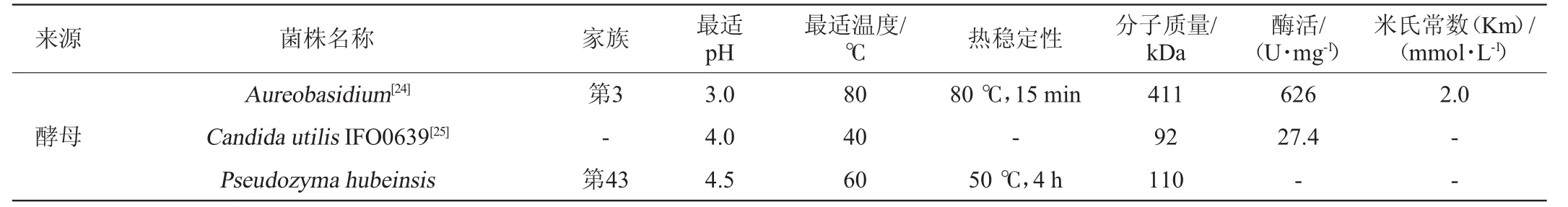
续表
由表2可知,大多β-木糖苷酶的最适pH在3.0~6.0。一般情况下,细菌所产β-木糖苷酶最适pH高于真菌β-木糖苷酶。当然,也有在更酸或中性pH下具有最佳活性的β-木糖苷酶。例如菌核青霉(Penicillium sclerotiorum)[8]所产β-木糖苷酶的最适pH为2.5;米曲霉(Aspergillus oryzae)所产β-木糖苷酶的最适pH为7.0[9];疏绵状嗜热丝孢菌(T.lanuginosus)[10]所产β-木糖苷酶的最适pH为6.5。
β-木糖苷酶所要求的反应温度一般比较温和,在40~60℃之间,而近年来人们越来越关注木糖苷酶的热稳定性。褐色高温单孢菌(Thermomonosporafusca)所产β-木糖苷酶在65℃的条件下保存8h以上仍有一定的活性,在70℃的条件下可保存1.5h[11]。粪堆梭菌(Clostridium stercorarium)所产β-木糖苷酶最适反应温度高达80℃,并且在50~70℃的环境下仍然具有良好的稳定性[12]。由于缺乏对木糖苷酶三维立体结构的确定,导致丝状真菌木糖苷酶热稳定性的结构因素至今还不得而知。木糖苷酶结构中存在的二硫键、疏水相互作用、电子对之间的相互作用可能是影响其热稳定性的原因。此外,在一些情况下,将硫醇和二价金属离子通过固定化或者化学修饰加到β-木糖苷酶上可以增强其热稳定性,从而防止温度的升高所导致的催化活性的降低,同时降低微生物污染的风险。耐热菌株的这一特性为酶的贮存和工业化利用提供了很大的便利。
真菌β-木糖苷酶的一个典型特征是其转木糖基化活性。转木糖基活性(transxylosylation activity)是通过形成新的糖苷键进而形成更长的低聚糖的一种能力(转糖基:将糖基供体上活化的糖基或单糖转移到糖基接受体的过程)[26]。由于其高选择性和立体定向性,这种转糖基作用也被用作酶法合成β-木聚糖酶底物和各类糖苷。SULISTYOJ等[27]报道了粉状曲霉(A.pulverulentus)所产具有转木糖基活性的β-木糖苷酶将低聚木糖的木糖残基转移到各种受体如醇和酚类化合物的过程。例如来自泡盛曲霉(A.awamori)X-100的β-木糖苷酶因其显著的转木糖基能力可以产生聚合度>7的低聚木糖[28]。
5 β-木糖苷酶的催化机制
半纤维素酶多为糖苷水解酶,一种广泛的水解两种或多种碳水化合物之间或碳水化合物和非碳水化合物部分之间的糖苷键的酶系[29]。在GHs分类中的酶,例如β-木糖苷酶,通过两种不同的机制裂解糖苷键来起作用:保留端基异位碳构型(保留型)或反转异位碳构型(反转型)[30]。糖苷水解酶第3家族和第54家族的成员采用保留型,第43家族则含有反转型糖苷水解酶。
目前人们已经比较清楚的是β-木糖苷酶的水解过程为酸碱催化反应,活性位点处的两个关键催化氨基酸(如谷氨酸或天冬氨酸)一个作为一般酸(质子供体),另一个作为亲核试剂[31]。其中,保留型酶采用双取代反应机制,包括糖基化反应和转糖基化反应;反转型则采用单取代机制[32]。青霉菌(Penicillium herquei)和炭色孢腔菌(Cochliobolus carbonum)所产β-木糖苷酶是由丝状真菌所产使用保留型机制的β-木糖苷酶。谷氨酸和天冬氨酸被普遍认为是所有丝状真菌β-木糖苷的催化残基[33]。
6 β-木糖苷酶的诱导和阻遏
木聚糖降解酶系中最重要的两种酶:木聚糖酶和β-木糖苷酶均为典型的诱导酶。大多数研究发现木聚糖水解酶的表达受到木聚糖或木糖的特异性诱导和CreA介导的碳分解代谢物阻遏。即碳源调节xlnR基因的转录,CreA介导阻遏xlnR基因的转录。因此,木聚糖降解酶基因的转录受到xlnR因子诱导和CreA介导的碳分解代谢物阻遏的共同影响。
木糖可以是木聚糖降解酶基因表达的诱导剂,有时也会抑制木聚糖降解酶基因的表达[34]。木糖触发CreA介导的碳分解代谢物阻遏取决于环境中木糖的含量。例如在含有1%燕麦木聚糖和0.1%木糖的培养基中诱导康氏木霉(Trichoderma koningii)中β-木糖苷酶基因的表达。关于T.koningii的研究表明,提供少量的木糖可以促进β-木糖苷酶的诱导表达[35]。相反地,D-木糖则会阻遏T.emersonni和T.reesei中β-木糖苷酶基因的表达。对于特定的微生物,木糖苷酶的表达由不同的诱导剂诱导。例如,诱导黑曲霉(A.niger)木聚糖降解酶系的碳源是木聚糖,木二糖和D-木糖[36]。埃默森篮状菌(T.emersonii)的木糖苷酶基因(bxl1)可以由较低浓度的木聚糖——甲基-β-D-吡喃木糖苷和木糖诱导[37]。
微生物的碳分解代谢物阻遏其实是一种节能机制,一旦环境存在着大量葡萄糖或者木糖,木聚糖降解酶就不必要合成。如在葡萄糖存在下,A.niger的β-木糖苷酶基因(xlnD)的表达被抑制[38]。此外,RAJOKA M I[39]发现,除诱导外,当微生物在生长于易于代谢的底物和无机氮源时还存在另外一种β-木糖苷酶合成调节机制。
7 β-木糖甘酶的分离纯化的研究现状
原核生物所产β-木糖苷酶是胞内酶,真菌所产β-木糖苷酶可分为胞内型、胞外型和膜结合型。通过对发酵上清液进行一系列分离纯化可以得到纯的β-木糖苷酶。其分离纯化方法多为硫酸铵沉淀、超滤浓缩、离子交换层析、凝胶过滤层析等,不同微生物来源的β-木糖苷酶的常见分离纯化步骤如表3。除上述常规方法,PAN I等[40]开发出了一种使用双水相萃取对T.koningii所产β-木糖苷酶同时进行浓缩和纯化的新方法。该方法快速、高效、回收率高且目标蛋白的纯度与柱层析的纯度相当。

表3 不同微生物来源的β-木糖苷酶的常见分离纯化步骤Table 3 Common isolation and purification steps of β-xylosidase from different microorganisms
8 β-木糖苷酶的应用
β-木糖苷酶在工农业残渣加工成燃料、化学和制药、医疗用品、食品和饮料、纤维饲料、造纸工业等方面都是非常重要的[41]。β-木糖苷酶的主要应用如下:
(1)自然界中大量存在的木聚糖,使得其生物转化后的产物低聚木糖和木糖有可能成为未来的能源。这种多糖转化为单糖可以通过两种方法进行:酸水解和酶水解。然而,酸水解过程会形成污染环境的副产物且成本高昂。因此,可再生型的生物转化具有重要意义,即先通过酶水解木聚糖得到木糖,再由如酵母和休哈塔假丝酵母(Candida shehatae)等微生物通过发酵将木糖转化为乙醇[42]。
(2)木寡糖是由木糖单元组成的低聚糖,具有不同的聚合度,被广泛用作天然食品甜味剂,具有良好的降血糖和血脂的作用。木聚糖酶和木糖苷酶复合酶是生产含有2~4个残基的木寡糖所必需的。另一方面,木寡糖作为益生元,可以选择性地促进益生菌的生长,从而缓解便秘,促进消化和营养吸收等。
(3)由于传统原料成本的增加,饲料中酶的添加量在过去几年中大大增加。木聚糖降解酶能够水解小麦,玉米和其他谷物中存在的半纤维素,促进营养物质的消化率,并减少氮磷的排放。单胃动物不产生植物细胞壁降解酶,如木聚糖和纤维素降解酶,导致谷物食品的利用率降低。这些酶的添加促进了能量代谢,降低了食物黏度,提高能量和蛋白质利用率,从而增加动物体质量[43]。
(4)木聚糖水解酶复合物也用于纸浆和造纸工业中,主要用于在增白之前加工纤维素纸浆。用木聚糖酶复合物进行预处理可以降低化学品,特别是氯和氧化氯的消耗。据报道,纸浆漂白中的酶的运用可以减少30%氯化合物使用量,减少环境中15%~20%的高毒性有机氯化合物的释放[44]。
(5)常用的酿造技术中的添加剂如乳化剂,氧化剂可以被酶所替代。在啤酒酿造中,长链阿拉伯木聚糖被释放,增加了酒体黏度和啤酒浊度。木聚糖酶和β-木糖苷酶的加入可以促进阿拉伯木聚糖溶解,通过将其降解为较小的寡糖,降低其黏度,消除啤酒浊度[45]。
9 结论与展望
随着人们对木聚糖降解酶系的深入研究,近年来有关β-木糖苷酶的生物化学和分子生物学上的研究取得了重大进展,但迄今为止进行的大多数研究集中在β-木糖苷酶的筛选、纯化、表征和重组表达。
首先,关于产β-木糖苷酶微生物的筛选,还没有直接简便的筛选方法。已知产β-木糖苷酶微生物多集中在细菌和霉菌,关于产β-木糖苷酶的酵母菌和具有完整木聚糖降解酶系统的酵母的研究还很欠缺。另一方面,植物性和动物性来源的β-木糖苷酶还有待进一步开发和研究。
其次,由于β-木糖苷酶在工业上的广泛应用使得其必须具有更高的产量、更高的活性和更好的生化特性,如热稳定性、pH稳定性等。为了提高β-木糖苷酶的产量和活性,比较常用的手段为诱变育种、基因工程育种、酶的固定化和调节酶的诱导和阻遏效应。其中诱变育种原理简单,但获得有益突变株的占比不多;相对基因工程育种,诱变育种方法的操作步骤更少。没有高通量筛选方法的条件下诱变育种的工作量较大,要处理大量突变株。然而基因工程育种目的性强,是目前用的较多、效果较好的一种手段,同时基因工程育种可以有效的改良β-木糖苷酶的生化特性,已经有过很多关于通过β-木糖苷酶基因的重组表达大大提高了其热稳定性,使其更高的应用于工业生产的报道。
由于缺乏三维立体结构测定的研究,β-木糖苷酶热稳定性和其结构之间的关系至今还不清楚。目前已知β-木糖苷酶的糖基化及其热稳定性,此外,将硫醇和二价金属离子通过固定化或者化学修饰加到β-木糖苷酶上可以提高其热稳定性。所以,β-木糖苷酶结构是未来值得研究的一个方向。
由于β-木糖苷酶是诱导酶又受到分解代谢物反馈抑制,目前比较清楚的是在一些情况下,少量木糖和乙醇可以促进β-木糖苷酶的表达,但关于低分子质量化合物诱导木聚糖水解酶系表达的分子机制还知之甚少;木聚糖和高分子量底物是β-木糖苷酶有效的诱导剂,一般情况下,β-木糖苷酶表达会受到木糖和葡萄糖引起的碳分解代谢物阻遏。所以,如何提高β-木糖苷酶木糖/葡萄糖耐受性或解除其阻遏效应或者探索β-木糖苷酶酶合成调节机制是亟待解决的问题。
最后,由于木聚糖的异质性,需要木聚糖水解酶系统:β-1,4-内切木聚糖酶、β-木糖苷酶、α-L-阿拉伯呋喃糖苷酶、α-D-葡萄糖醛酸酶、α-D-半乳糖苷酶、乙酰木聚糖酯酶、对香豆酸酯酶和阿魏酸酯酶这一系列酶的协同作用将其降解至木糖。目前研究较多的是β-1,4-内切木聚糖酶和β-木糖苷酶间的协同作用,关于其他酶之间的协同作用和实用性研究还很欠缺。然而,已有报道,某些微生物所产β-木糖苷酶还同时具有α-L-阿拉伯呋喃糖苷酶活性,对于含有两种或多种酶活性的β-木糖苷酶的来源和原理仍然值得进一步探究。
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08
农业工程学报(2022年12期)2022-09-09
红蜻蜓·低年级(2022年5期)2022-05-11
生物信息学(2022年1期)2022-04-01
红蜻蜓·低年级(2022年2期)2022-03-19
红蜻蜓·低年级(2021年12期)2021-12-19
陶瓷学报(2019年5期)2019-01-12
科技创新导报(2018年1期)2018-05-07
纺织导报(2017年4期)2017-05-05
中国塑料(2015年6期)2015-11-13
