
CpG⁃ODN通过调控细胞免疫显著增强热休克蛋白肿瘤疫苗诱导的抗肿瘤免疫效应
2018-11-07 01:12张莹莹王晓文唐劲天
转化医学电子杂志 2018年9期
张莹莹,王晓文,唐劲天
(1中南大学湘雅医院肿瘤科,湖南 长沙410078;2清华大学工程物理系医学物理与工程研究所,北京100084)
0 引言
随着医学的不断发展,肿瘤治疗的技术也在不断的进步。但是对于很多恶性肿瘤来说,常规治疗效果欠佳,死亡率居高不下,仍是临床治疗中的难点。近年来,研究者们发现,部分恶性黑素瘤患者出现肿瘤自发性的消退现象,于是逐渐关注黑色素瘤固有的致免疫性。随着研究的不断深入,人们将其认为是恶性肿瘤中具有高免疫原性的一种。因此,免疫治疗开始成为恶性黑色素瘤治疗的研究热点和发展方向,并期待取得突破性进展[1]。
已有研究[2-3]报道热疗可以增强肿瘤细胞的免疫原性,且这种增强在很大程度上与热休克蛋白(heat shock proteins,HSPs)的过表达有关。大量的实验研究[4-5]已经证实了经热诱导的癌细胞中的HSP70、HSP90等物质能够激发抗肿瘤免疫。然而,众多热休克蛋白相关肿瘤疫苗的动物或临床研究[6-8]均显示,虽然在实验对象体内可以检测到特异性免疫反应并且肿瘤生长受到不同程度地抑制,但尚难达到满意的疗效。分析原因,发现有可能与MHC⁃I抗原的表达不强有关[9]。甚至有研究[3]表明热休克处理虽然可以增加抗原肽形成,但同时抑制了抗原的呈递。B16细胞经热诱导后,会形成免疫原性的抗原。例如检测细胞中的HS,发现其P⁃抗原复合物的含量有所上升,但是MHC⁃I分子却不能有效地将抗原传递给T细胞,从而造成免疫耐受或逃逸。因而,比较理想的肿瘤疫苗应该能够有效地激活T细胞,为其提供良好的第一和第二信号,将其特异性抗肿瘤免疫效应充分发挥出来[8],同时也避免由于免疫强度不够而发生逃逸,影响患者身体健康。
要想抗原诱导机体更好地发生免疫反应,可适当地添加佐剂,适当的佐剂能够为T细胞的活化提供第二信号。所以在疫苗的研究过程中,对佐剂的研究也是其中很重要的部分。目前使用比较多的佐剂大多是人工合成的,寡聚核苷酸链(oligodeoxynucleoti⁃des,ODN)中含有非甲基化 CpG 基序(CpG⁃ODN),也是一种近年新引起广泛关注的水性佐剂。其自身无免疫原性,不会引起自身免疫疾病。更重要的是CpG⁃ODN作为Toll样受体9的配体,能够提高蛋白抗原和DNA疫苗的免疫原性,直接激活 NK细胞[10]、树突状细胞[11]、巨噬细胞[12]等,并刺激这些细胞分泌 IFN⁃γ、IL⁃2、TNF⁃α 等 Th1 样细胞因子,从而诱导 Th1型细胞免疫应答[13-14]。 此外,它还能够在一定程度上促进细胞抗原呈递,促进其细胞表面分子MHC⁃II、B7⁃1、B7⁃2 等的表达,活化树突状细胞,促进抗原呈递,协同激活 T 细胞[15-16]。 因此,可将 CpG⁃ODN作为多种抗原的免疫佐剂,以增强免疫作用[13,15,17-19],在抗肿瘤免疫治疗领域显示出巨大的潜力。
综上所述,富含热休克蛋白的肿瘤细胞瘤苗和CpG⁃ODN在抗肿瘤免疫功能上具有互补性,CpG⁃ODN有望克服单纯肿瘤细胞疫苗抗原呈递活性的不足,起到协同增效的作用。本研究的实验模型是小鼠恶性黑色素瘤,肿瘤疫苗选择经过热诱导且富含HSP70的B16细胞裂解的产物,联合新型免疫佐剂CpG⁃ODN免疫小鼠,观察疫苗对小鼠产生的免疫保护作用,并研究小鼠免疫系统的功能改变。
1 材料和方法
1.1 主要试剂羊抗鼠HSP70多克隆抗体,购自美国Cellsigna公司;抗羊二抗液,被辣根过氧化物酶标记,购自北京中杉金桥生物技术有限公司,大鼠抗小鼠CD4 R⁃PE和CD8a TRI⁃COLOR荧光抗体、大鼠抗小鼠CD25⁃FITC荧光抗体均购自美国Invitrogen公司;脾淋巴细胞分离液,购自达科为生物有限公司;小鼠 IL⁃2、 IFN⁃γELISA 试剂盒,购自美国 Bioscience公司;LDH释放法检测试剂盒购自美国Promega公司(Madison, WI)。
1.2 实验动物实验小鼠,健康清洁级C57BL/6,雄性,体质量18~22 g,6~7周,购自中国医学科学院实验动物中心。小鼠饲养时温度为(23±2)℃,相对湿度(55%±2%),普食饲养。
1.3 细胞培养小鼠B16黑色素瘤细胞购自中国协和医科大学细胞库。B16细胞接种于DMEM培养基,接种之后放于37℃、5%CO2环境下培养约24 h即可进行传代,观察发现细胞的数量呈对数期增长时即可提取用于实验。
1.4 制备热处理后细胞裂解瘤苗将经45℃15 min热处理后的B16细胞(我们前期实验研究发现,相较42℃ 60 min和50℃ 5 min,45℃ 15 min热处理能诱导最高程度的HSP70蛋白表达,该western blot结果在补充材料中提供),立即更换培养基后放回37℃,5%CO2细胞培养箱复温孵育24 h后取出,收集全部细胞,用PBS将细胞洗涤两次,然后将其浓度调整至5×108个/mL。即可将细胞悬液转移至冷冻管中进行保存,然后再交替置于液氮内10 min,37℃水浴中5 min,多次操作之后待细胞完全溶解后,即可放于-80℃的环境中进行冷冻保存,以备后用。
1.5 CpG⁃ODN2395佐剂配制CpG⁃ODN2395 的序列为 5′⁃TCGTCGTTTTCGGCGCGCGCCG⁃3′,购自上海生工生物工程技术服务有限公司。根据说明书,使用PBS溶解核酸,将浓度调制为0.1 mL/OD,注意即配即用。
1.6 实验分组以及小鼠肿瘤疫苗接种及攻击实验实验小鼠随机分为下述四组。①对照组:接种PBS;②瘤苗组(heated cell lysates,HCL组):将45℃热处理肿瘤细胞裂解产物(每0.1 mL含2×107个细胞的裂解产物)+PBS等体积混合后接种;③佐剂组(CpG⁃ODN 组): CpG⁃ODN2395(浓度为 30 μg/0.1 mL) +PBS等体积混合后接种;④联合组(HCL+CpG⁃ODN组):将 45℃热处理肿瘤细胞裂解产物+CpG⁃ODN2395等体积混合后接种。
健康小鼠的疫苗接种:分别于第 0、6、12、18及24 d向小鼠注射疫苗,需注意疫苗应分四点进行注射,注射位置为小鼠双侧腹股沟和腋窝皮下,每个位置的注射量为0.05 mL,每间隔5 d进行一次注射,每只小鼠接种5次。
B16细胞攻击接种后小鼠:当B16细胞呈对数生长之后,取细胞放于0.25%胰酶中进行消化。重悬于PBS中。并将 B16的浓度调至 1×106个/mL,于第30天在小鼠为右肋腹部皮下位置进行注射,每只小鼠注射 0.1 mL。
观察并记录小鼠皮下肿瘤出现的时间、体积大小和小鼠生存天数。
1.7 外周血T细胞亚群检测免疫结束后的第六天(d30),随机从每组中挑选3只小鼠,用摘眼球取血法采集小鼠血液,并置于抗凝管中。在抗凝管中取血100 μL,置于 1.5 mL 离心管,分别向离心管中加入3 L的,在温室、避光的环境中孵育15~20 min。1000 rpm离心 5 min,弃上清。加入 2 mL PBS,1000 rpm离心5 min,将未结合抗体洗去。在溶液中加入稀释了的流式红细胞裂解液,在温室、避光的环境中孵育15 min,观察发现溶液呈透明状后取出。以1000 rpm的速度离心溶液离心5 min,弃上清。向其中加入2 mL PBS,再以1000 rpm的速度离心溶液离心5 min,重复洗涤2次。加入4%多聚甲醛,固定待测。用流式细胞仪检测外周血中CD4+T细胞、CD8+T细胞以及CD25+T细胞的数量,计算各亚群比例。
1.8 小鼠脾淋巴细胞分离和培养取血后将小鼠断颈处死,无菌取脾,置于200目尼龙网中研磨脾组织,分散的细胞经过尼龙网过滤即可进入到脾淋巴细胞分离液中。收集悬液,以1500 rpm的速度离心溶液离心30 min(密度梯度法),得到脾脏单个核细胞。用PBS液将细胞漂洗两次,于5 mL 10%FCS RPMI 1640培养液重悬(含 IL⁃2 100 U/L),以常规细胞培养方式进行培养,培养液隔天更换。
1.9 ELISA法检测小鼠脾细胞分泌Th1型细胞因子IL⁃2、INF⁃γ水平将所获各组小鼠脾淋巴细胞(单核细胞)作为效应细胞,以丝裂霉素C灭活处理后B16细胞作为刺激细胞,按10∶1的比例共培养3 d,收集培养上清,ELISA法检测其中Th1型细胞因子 IL⁃2、INF⁃γ 的水平。
1.10 LDH释放法检测免疫后小鼠脾细胞毒性T淋巴细胞(cytotoxic lymphocyte,CTL)对B16的特异性细胞毒作用将效应细胞(分离的脾单核细胞)和靶细胞(灭活的B16细胞)以效靶比为20∶1混合培养于10%FCS RPMI 1640 培养液(含 IL⁃2 100U/L)中,4 h后用LDH释放法检测试剂盒检测不同组LDH的释放水平,CTL杀伤效率计算公式:杀伤率(%)=OD(实验组⁃效应细胞自发⁃靶细胞自发)OD(靶细胞最大⁃靶细胞自发)×100%。
1.11 统计学分析所有实验均重复至少两次。实验数据采用 StatView (version 5.0) software(SAS Institute Inc.Cary,NC)统计软件处理。计量数据以x ±s表示,组间均数比较采用ANOVA分析,统计学差异显著性采用 log⁃rank检验,P<0.05为差异有统计学意义。
2 结果
2.1 同源肿瘤细胞攻击后小鼠出瘤情况、肿瘤生长曲线及生存期观察的比较总观察期为40 d,将小鼠体内肿瘤体积绘制成动态图(图1):PBS对照组肿瘤生长迅速,平均出瘤时间为(5.3±1.8) d,并在 30 d内全部死亡(小鼠的生存周期为 20.9±2.9 d)。 相比于PBS组,CpG⁃ODN组、HCL组以及联合疫苗组(HCL+CpG⁃ODN)的小鼠肿瘤均受到不同程度的抑制,其中联合疫苗组的肿瘤抑制水平最高(P<0.05)。小鼠接瘤之后,直至观察结束,约有35%未出瘤。已出瘤小鼠的平均出瘤天数为(11.7±5.0)d,超过 73%的小鼠在观察结束之后仍然存活,存活时间明显长于其他组(P<0.01)。 CpG⁃ODN 组未出瘤小鼠仅为一只,出瘤小鼠的出瘤时间为(8.6±3.2)d,小鼠 40 d 的存活率为52%(14/27)。HCL组和PBS组的出瘤时间基本相同,为(5.6±1.7) d,但是在肿瘤生长抑制作用上,两组的差别比较明显,HCL组46%(12/26)的小鼠生存期超过了40 d,较对照组延长。CpG⁃ODN组相较于HCL组来说,无论是平均出瘤时间,还是生存期都有所延长。
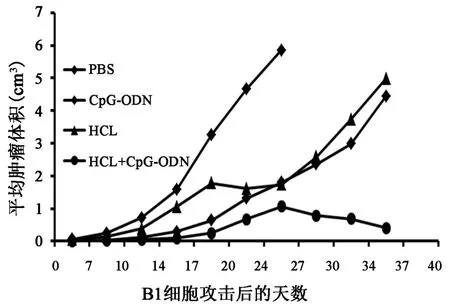
图1 小鼠肿瘤体积生长曲线图
2.2免疫后小鼠外周血T细胞亚群CD4+、CD8+的比值升高,但CD4+CD25+T细胞比例无差异研究显示机体的免疫功能状态在很大程度上与T细胞亚群的比例有关,以流式细胞术对细胞外周血进行检测,CD4+T细胞与CD8+T细胞比值低提示免疫功能低下,而 HCL 组、CpG⁃ODN 组与 HCL+CpG⁃ODN 组小鼠的外周血CD4+/CD8+的值均较PBS组升高(P<0.001)。 其中联合疫苗组 CD4+/CD8+的值最高,与其它治疗组相比,差异有统计学意义(P<0.001)。HCL 组和 CpG⁃ODN 组的 CD4+/CD8+值比较,差异有统计学意义(P=0.048,图 2)。

图2 外周血T细胞亚群分布比例
2.3 联合免疫小鼠脾细胞分泌Th1型细胞因子的活性显著增加治疗组IFN⁃γ和IL⁃2的水平与对照组相比,均有不同程度的提升(P<0.05)。 其中 HCL+CpG⁃ODN组的小鼠可诱导出最高水平的IFN⁃γ和IL⁃2,分别为(1402.5±156.5) pg/mL 和(532.9±41.8) pg/mL;CpG⁃ODN 组也可诱导出较高的 IFN⁃γ 和 IL⁃2,分别为(1021.8±136.9) pg/mL 和(395.3±20.1) pg/mL;HCL免疫组诱导出的IFN⁃γ和IL⁃2则相对较低,分别为(654.7±59.5) pg/mL 和(289.2±15.5) pg/mL,组间比较,差异有统计学意义(P<0.05,图 3)。
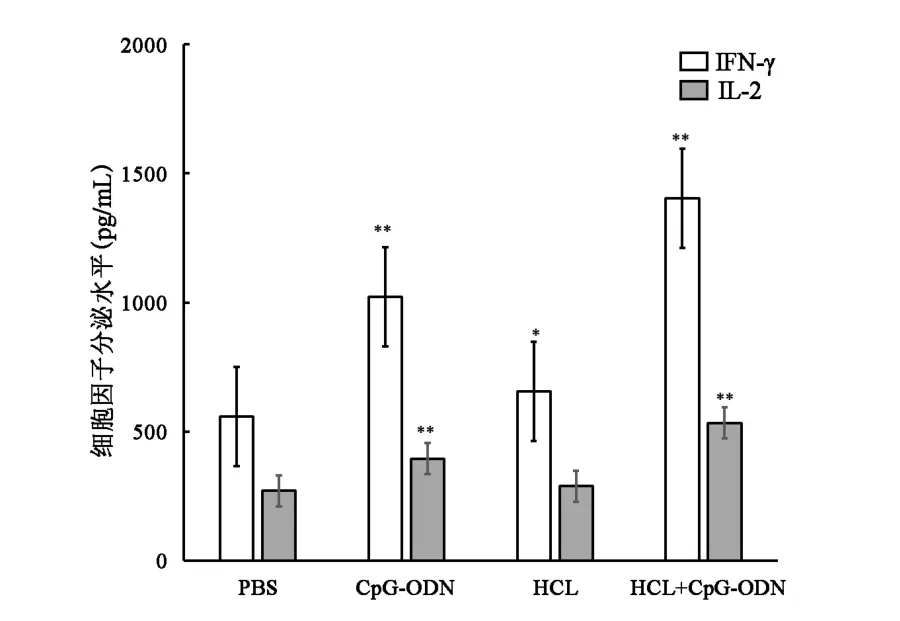
图3 各组Th1细胞因子分泌水平比较
2.4 联合免疫后小鼠脾CTL的细胞毒效率明显增加对CTL细胞进行毒效率检测,相比于PBS组,单独HCL起不到增强CTL特异性细胞毒效率的作用(P>0.05)。 CpG⁃ODN 组和 HCL+CpG⁃ODN 组均能明显增强小鼠 CTL的细胞毒杀伤效率(P<0.05),HCL+CpG⁃ODN 组与 CpG⁃ODN 组相比,差异有统计学意义(P<0.01),对细胞毒杀伤效率的增强作用最显著(表 1)。
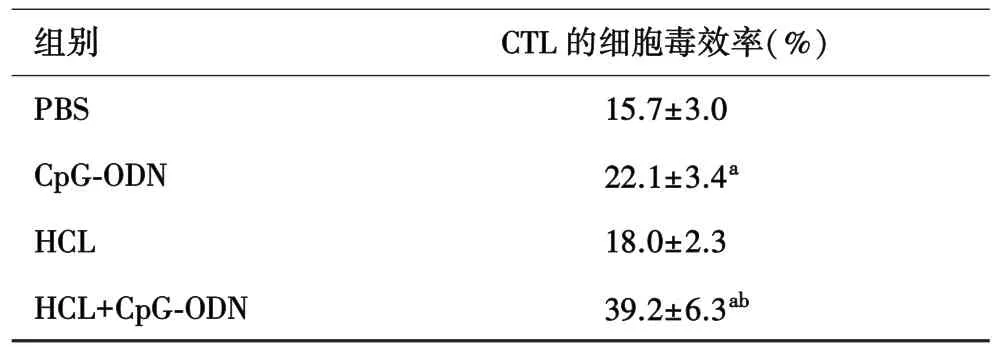
表1 各组脾CTL对B16的特异性细胞毒杀伤效率比较
3 讨论
热休克蛋白相关肿瘤疫苗及免疫活性物质CpG⁃ODN,在诱导抗肿瘤免疫反应上,能够在一定程度上起到互补的作用。本实验主要研究了小鼠的恶性黑色素瘤肿瘤细胞疫苗的疗效,并观察了不同疫苗配方对小鼠的抗肿瘤免疫效力的增强作用,通过检测,联合疫苗的效果最佳。联合疫苗能使超过35%的小鼠抵抗住了B16细胞的攻击。即使长出肿瘤的小鼠,其平均出瘤时间、生存期与其它各组相比亦有显著性延长。本研究结果与Ito等[18]研究具有一致性,进行HCL瘤苗治疗,小鼠未能产生更强的免疫保护,HCL组的出瘤时间与PBS对照组相比,差异无统计学意义,但是在肿瘤的生长和小鼠的生存期上有一定的差异,说明HCL瘤苗有一定的诱导抗肿瘤免疫作用,这可能与热处理后肿瘤细胞内形成的大量HSP⁃抗原肽复合物具有一定抗肿瘤免疫活性且不依赖于MHC的表达有关[20-21]。 另外,值得一提的是 CpG⁃ODN 与HCL联合应用后部分小鼠出现了因局部变态反应导致的皮肤溃疡(该结果未在本文中列出),此现象在国内外尚未见类似报道,具体原因还有待进一步研究。此局部剧烈变态反应的出现除了证实联合疫苗可诱导极强的免疫反应外,也提醒我们需改良接种方式以减轻治疗的局部不良反应。
为对抗肿瘤机制进行更进一步的研究,本实验还对小鼠细胞免疫状态进行了检测。在小鼠体内,CD4+T细胞是细胞免疫应答的主要效应细胞,通过分泌 IL⁃2、INF⁃γ 等细胞因子来调节免疫反应;CD8+T细胞能够分泌物质,抑制T细胞因子的免疫作用,对于小鼠机体而言,CD4+/CD8+细胞比值决定着免疫是否平衡,从侧面反映出机体免疫功能状态,当CD4+/CD8+细胞比值下降,说明小鼠机体的免疫功能下降,而当免疫功能有所恢复,小鼠的CD4+/CD8+细胞比值会有所上升。本实验结果显示,当机体进行免疫治疗之后,CD4+/CD8+细胞比值均有明显的上调,其中联合疫苗组的上调最为明显。后续IL⁃2、INF⁃γ细胞因子以及CTL细胞毒杀伤效率的检测结果亦印证了该结果。
CD4+CD25+T细胞在免疫应答过程中起着负调节的作用,所以也将其称为调节性T细胞(T regular cell, Treg)。 有研究报道减少或清除 CD4+CD25+T细胞可以增强细胞疫苗的疗效[22],而CpG⁃ODN可通过TLR9损伤Treg细胞使其数目减少[23]。因此,本实验预期联合免疫组小鼠CD4+CD25+T细胞比例会低于对照组,但结果却显示各实验组无明显差异。迄今为止,Treg细胞的调控机制尚未完全明确,Treg细胞可与细胞因子、T细胞、树突状细胞以及不同肿瘤和免疫微环境等多种因素相互影响、相互调控有关[24],本研究中各组小鼠Treg细胞亚群数量无明显差异,亦反过来证实了Treg细胞调控机制的复杂性。下一步我们拟检测Treg的功能状态,探索其与CpG⁃ODN相互作用的可能信号通路,以期进一步揭示CpG⁃ODN强化抗肿瘤免疫应答的可能分子机制。
综上所述,新型免疫佐剂CpG⁃ODN可显著增强热休克蛋白肿瘤疫苗的抗肿瘤免疫效应。HCL携带多种肿瘤特异或者相关抗原的HSP⁃抗原肽复合物,在免疫佐剂CpG⁃ODN的协同作用下,免疫细胞因子分泌将会更强,改善T细胞亚群的比例,活化肿瘤免疫微环境,进而增强单核细胞对肿瘤细胞的特异性细胞毒作用,从而抑制肿瘤生长。这些实验结果为未来肿瘤疫苗的研究提供了一定的实验依据。
猜你喜欢
昆明医科大学学报(2021年12期)2021-12-30
新世纪智能(数学备考)(2021年10期)2021-12-21
昆明医科大学学报(2021年10期)2021-12-02
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年1期)2021-02-07
新世纪智能(数学备考)(2020年10期)2021-01-04
中国兽医杂志(2019年5期)2019-09-18
生物学教学(2018年10期)2018-11-29
中国交通信息化(2017年8期)2017-06-06
医学研究杂志(2015年12期)2015-06-10
