
低能离子注入介导的异常汉逊酵母菌基因组重复序列的突变研究∗
2018-11-16 06:58张寒玉冯光文钱卫东蔡长龙毛培宏
新疆大学学报(自然科学版)(中英文) 2018年4期
张寒玉,王 婷,唐 朝,冯光文,钱卫东,蔡长龙,毛培宏
(1.新疆大学物理科学与技术学院,离子束生物技术中心,新疆乌鲁木齐830046;2.陕西科技大学食品与生物工程学院,陕西西安710032;3.西安工业大学离子束生物工程与生物多样性研究中心,陕西西安710032)
0 引言
低能离子注入到生物细胞内,可参与细胞的生理、生化反应,并影响生物的进化,其相互作用是一个极为复杂的物理学、化学和生物学过程[1,2].低能离子注入生命体的方法和技术已广泛应用于植物和微生物的品种改良、生命起源与进化、环境辐射与防护以及人类健康等领域[3−8].
重复序列是真核生物基因组中的重要组成部分,主要分为串联重复序列和散在重复序列两类[9].基因组中的串联重复序列包括卫星DNA[10]、小卫星DNA[11]和微卫星DNA[12].其中,微卫星DNA又称为简单重复序列(Simple Sequence Repeats,SSRs),随机分布于生物体整个基因组中,是目的基因筛选、基因诊断及其多样性分析、遗传连锁图谱构建等工作的理想的分子遗传标记.而散在重复序列,即转座子元件(Transposable Element,TE),分为RNA转座子和DNA转座子,不仅可以影响真核生物基因组的大小,还能直接或间接促成基因组重排,并影响基因表达水平、改写基因调控网络.因此TEs在基因组的进化中具有重要意义[13].
异常汉逊酵母(Hansenula anomala)是一种重要的非酿酒酵母,具有较强的产酯生香能力,且在较高温度的条件下具有较强的发酵力和酯化力,广泛用于高品质酿造食品的研发[14−18].
本研究在全基因组de novo测序的基础上,利用生物信息学方法分析低能离子注入对异常汉逊酵母菌基因组中各种重复序列的致突变分子信息,以期深入认识低能离子注入对酵母菌基因组结构特征的影响,并为异常汉逊酵母菌的分子育种及SSR分子标记的开发提供理论依据,为开展异常汉逊酵母菌遗传多样性和遗传结构的研究奠定基础.
1 材料与方法
1.1 菌株及其全基因组De novo测序
异常汉逊酵母菌来源于陕西科技大学食品与生物工程学院功能微生物研究室,常规方法传代、培养,收集遗传稳定的纯培养菌体.根据低能离子注入生命体产生的“反常辐照损伤”生物效应的基本原理[1],应用能量15 KeV、剂量10×1015ions/cm2的Ar+注入异常汉逊酵母菌菌体,参照L¨u的方法[3]进行离子注入后的菌体洗脱、分离、传代、培养,获得离子注入后的遗传稳定的纯培养菌体.
采用常规方法分别制备离子注入前后的异常汉逊酵母菌基因组DNA,并应用PacBio单分子测序技术分别对其进行全基因组De nove测序,所获得的全基因组DNA序列,作为本研究的基本数据.
1.2 基因组中重复序列的获取
应用TRF(Tandem Repeat Finder)方法(http://tandem.bu.edu/trf/trf404.linux64.download.html)来获取离子注入前后的异常汉逊酵母菌全基因组DNA序列中的串联重复序列,最大的重复单元bp数设置为2 000bp.
对TRF获取的结果进行细分,设置微卫星DNA序列重复单位为2∼6bp,小卫星DNA序列重复单位为10∼60bp.
使用RepeatMasker方法(http://www.repeatmasker.org/RMDownload.html)获取离子注入前后的异常汉逊酵母菌全基因组DNA序列中的散在重复序列.
2 结果与分析
2.1 异常汉逊酵母菌基因组中微卫星DNA序列的突变特征
利用TRF方法在低能离子注入前后的异常汉逊酵母菌基因组重复序列中各发现了175个SSR.虽然SSR数目没有变化,但离子注入后,异常汉逊酵母菌基因组中SSR的总长度从8 310bp减少到8 163 bp,SSR在基因组中的分布频率从78.63Kb减少到77.98Kb.
在SSR的重复类型方面,离子注入后,异常汉逊酵母菌基因组SSR中的三碱基(Tri-)类型的数目从94条增加到97条,而四碱基(Tetra-)和五碱基(Penta-)类型分别减少了2条和1条.SSR在离子注入前后的异常汉逊酵母菌基因组中的分布如图1所示.
在SSR的各种类型碱基重复基序方面,由于离子注入的作用,异常汉逊酵母菌基因组中SSR三碱基重复基序的GAG和ATC分别突变为AGG和ATG.除ACC外,其它三碱基重复类型的数目均发生了变化.离子注入后,四碱基重复基序的GAAT和GTTG分别突变为GTTA和TTCA,虽然GTTA的数目只有1条,但其累积长度为128bp,占四碱基重复总长度的18.85%,其它每种类型的四碱基重复基序的累积长度范围为25bp∼27bp.而五碱基重复基序中,有9种类型重复基序发生了碱基突变.在六碱基重复基序中,突变的重复基序类型为28种,同时新增1种重复基序类型.离子注入前后,异常汉逊酵母菌基因组中SSR各碱基重复基序的统计结果见表1.
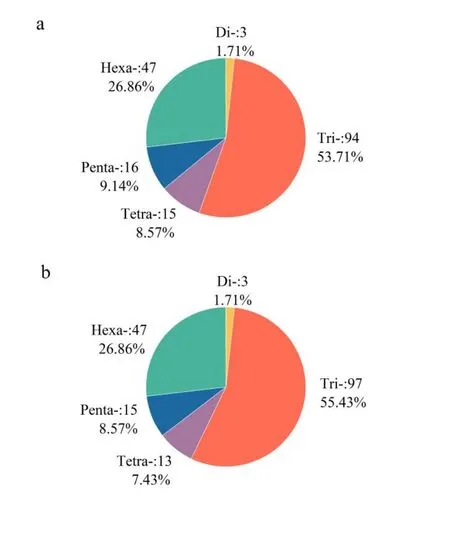
图1 离子注入前后异常汉逊酵母菌基因组中SSR的分布
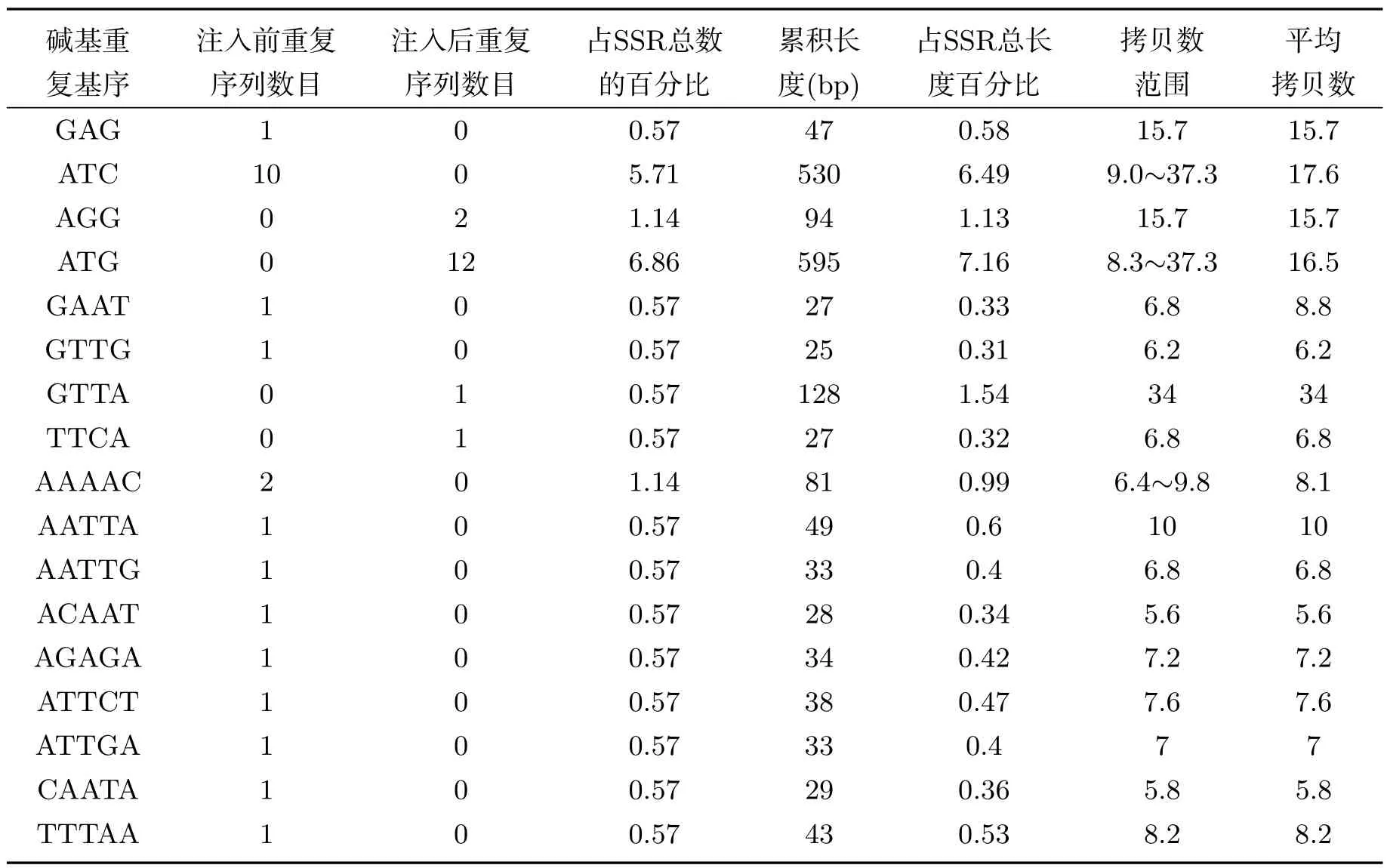
表1 离子注入前后异常汉逊酵母菌基因组微卫星DNA各类型重复序列中重复基序分布
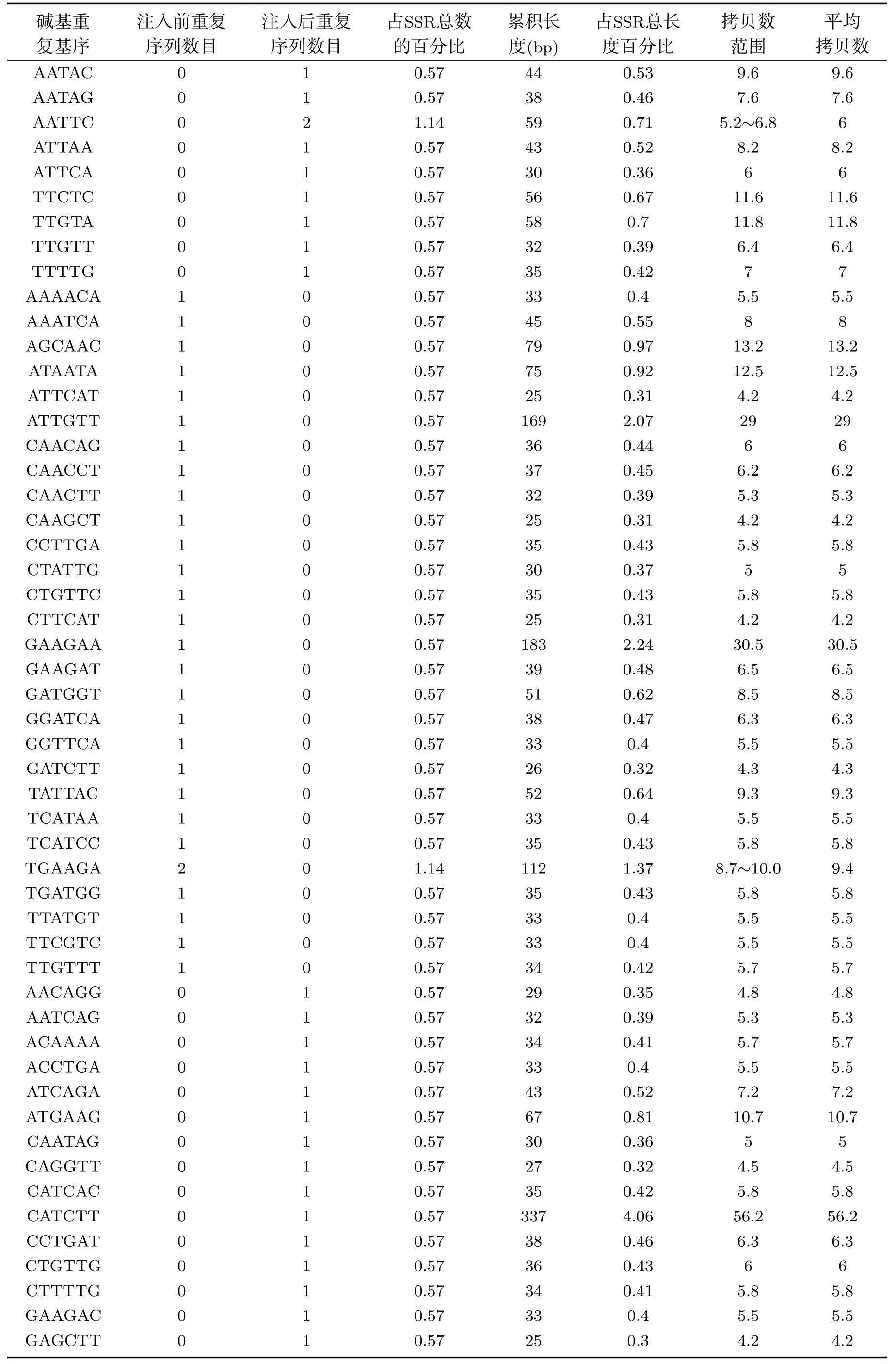
表1 续离子注入前后异常汉逊酵母菌基因组微卫星DNA各类型重复序列中重复基序分布
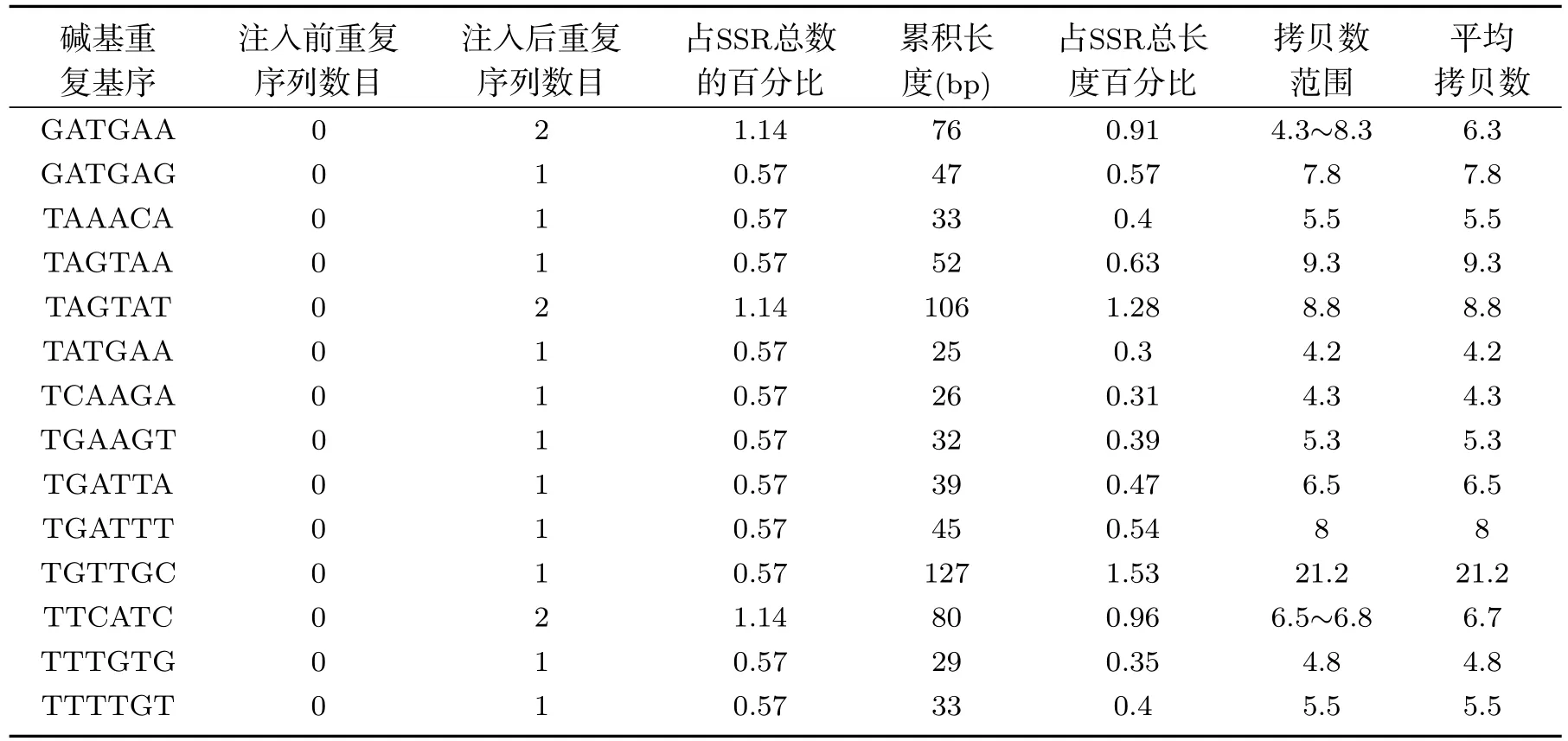
表1 续离子注入前后异常汉逊酵母菌基因组微卫星DNA各类型重复序列中重复基序分布
各种重复类型的拷贝数分析结果(表2)表明,异常汉逊酵母菌基因组微卫星DNA序列均在低拷贝区出现频率较高,离子注入前后,拷贝数低于15次的微卫星序列占比最高,分别为75.43%和73.14%;其次是拷贝数在15∼27之间的微卫星序列,占比分别为17.14%和18.29%;拷贝数在27∼39之间的占比分别6.86%和7.43%;拷贝数大于39次的微卫星序列最少,仅占0.57%和1.14%,离子注入增加了高拷贝区的微卫星序列数目.从五种重复类型的平均拷贝数来看,离子注入前后变化不大,如图2所示.离子注入前的五种重复类型的平均拷贝数分别为33.00、15.04、12.76、7.33、8.51,离子注入后分别为33.00、15.43、13.11、8.00、8.44.
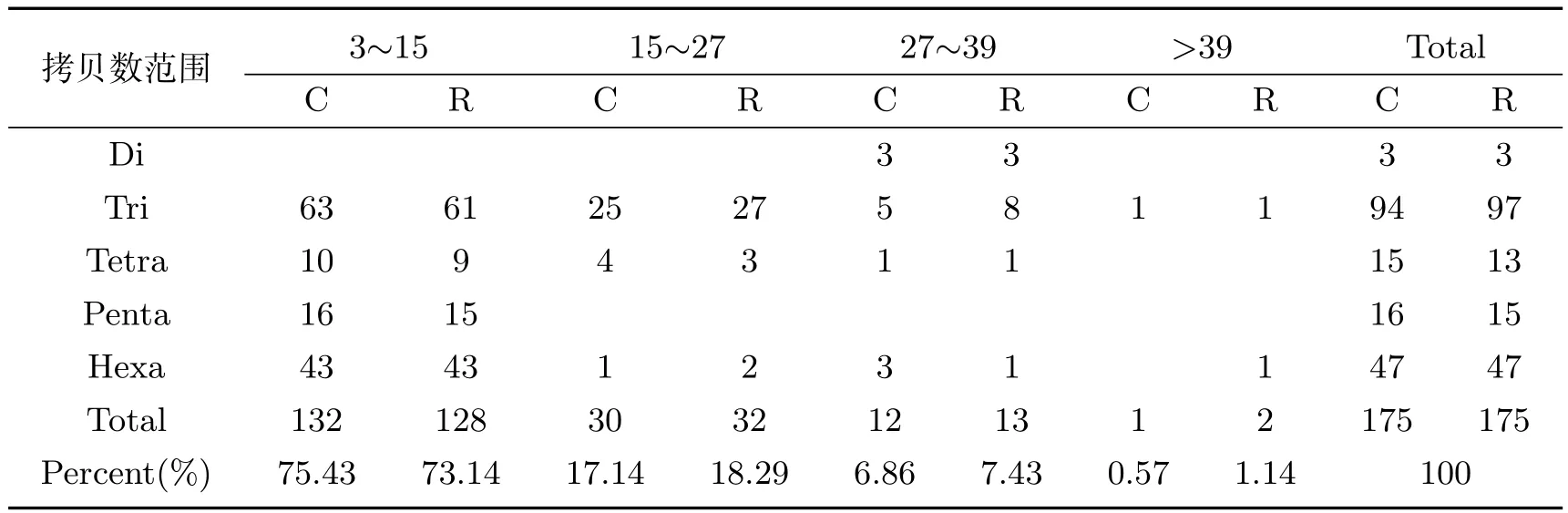
表2 离子注入前后异常汉逊酵母菌基因组中不同类型的微卫星DNA序列拷贝数分布

图2 离子注入前后异常汉逊酵母菌基因组中SSR五种重复类型的单元长度与拷贝数的变化
2.2 异常汉逊酵母菌基因组中小卫星DNA序列的突变特征
利用TRF方法在低能离子注入前后的异常汉逊酵母菌基因组的串联重复序列中各发现小卫星DNA序列1 384条和1 361条,总长分别为79 849bp和74 362bp.由此可见,离子注入导致23条小卫星DNA序列缺失,总长度达5 487bp.
离子注入前,异常汉逊酵母菌基因组小卫星DNA序列中,长度为15bp的重复单位序列数目多达211条,占小卫星序列总数的15.52%,其次是12bp(175条)、18bp(160条)、21bp(116条)、13bp(62条)和24bp(59条);重复单元是15bp的序列累积长度最长,8 328bp,占小卫星序列总长的10.43%,其次是18bp(8 010bp)、12bp(7 507bp);重复单元的拷贝数范围分布最广的前5种序列是:13bp(1.9∼42.5)、12bp(2.1∼36.5)、42bp(1.9∼22.5)、21bp(1.9∼18.5)、36bp(2.0∼17.8).
离子注入后,酵母菌基因组小卫星DNA序列中,重复单位为15bp的序列数目虽然多达192条,但比离子注入前缺失了19条.除此之外,重复序列数目较多的前五位分别是12bp(175条)、18bp(165条)、21bp(111条)、13bp(60条)和20bp(60条),其中以18bp为重复单元的序列长度最长,共计8 258bp,占小卫星DNA序列总长的11.11%.其次是重复单位为15bp的序列,总长7 673bp;重复单位为12bp的序列,总长7 132bp.重复单元的拷贝数范围分布最广的前5种序列分别是:12bp(2.1∼20.7)、21bp(1.9∼19.6)、36bp(2.0∼17.4)、42bp(2.0∼16.6)、24bp(1.9∼13.0).
与离子注入前相比,离子注入后的异常汉逊酵母菌基因组小卫星DNA序列中缺失了重复单元为47bp和56bp的小卫星DNA序列,新增了重复单元为35bp、58bp和59bp的小卫星DNA序列.
异常汉逊酵母菌基因组中的小卫星DNA序列数目与其重复单位长度有一定关系,随着重复单位长度的增加呈下降趋势,这一特征在重复单元大于15bp的小卫星序列中尤为显著;与微卫星DNA序列类似,小卫星DNA序列重复单位拷贝数较低,主要分布在1∼3次;重复单元拷贝数与小卫星序列之间无显著的相关关系.
离子注入前后异常汉逊酵母菌基因组中小卫星DNA序列的重复类型数目、序列长度及拷贝数的变化如图3和图4所示.
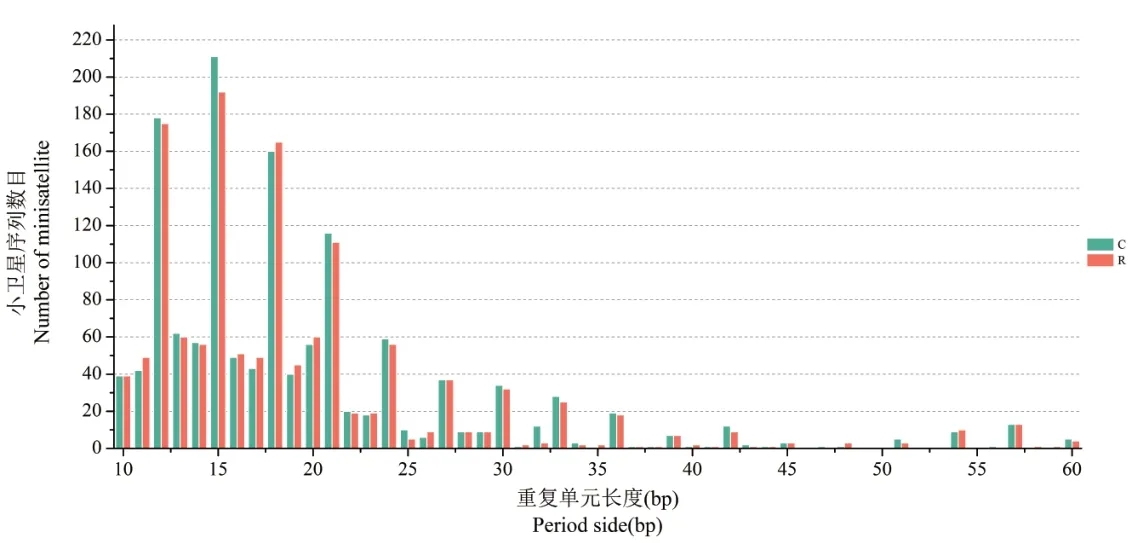
图3 离子注入前后酵母菌基因组中小卫星DNA序列的重复单元长度与其数量的变化
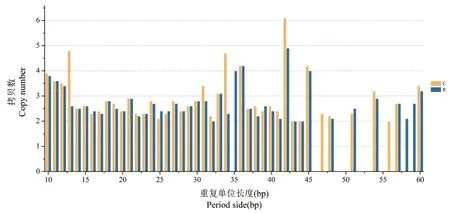
图4 离子注入前后酵母菌基因组中小卫星DNA序列的重复单元长度与其拷贝数的变化
2.3 异常汉逊酵母菌基因组中散在重复序列的突变特征
运用RepeatMasker方法,获得了离子注入前后异常汉逊酵母菌基因组中的多种散在重复序列(表3),其在基因组中占比仅为0.78%左右.离子注入前,长末端重复DNA序列(LTR)数目最多,为695条,占总数的46.30%;其次是DNA转座子,为459个;长散在重复DNA序列(LINE)共303条;而短散在重复DNA序列(SINE)只有25条;滚环(RC)14个.
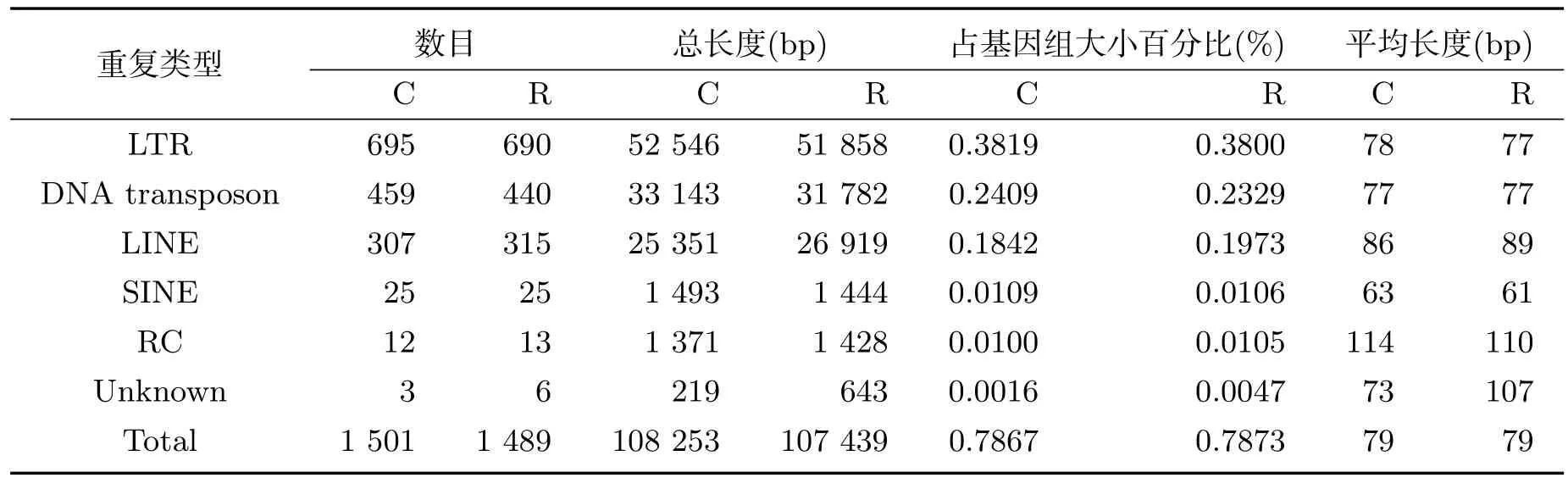
表3 离子注入前后异常汉逊酵母菌基因组中散在重复DNA序列的分布
离子注入后,异常汉逊酵母菌基因组中的LTR数目缺失了5条,DNA转座子缺失了19个,LINE新增加了12条,RC缺失了1个,而SINE没有变化.
3 讨论
真核生物基因组中的重复序列对生物的进化、遗传和基因的表达与调控有重要作用.基因组中重复序列的出现,表明基因组DNA在不断地进行自我复制,并进行水平交换和垂直交换,其重要作用是不断丰富生物的遗传信息[19].生物体中许多关键基因是单拷贝的,重复序列的存在能保护这些重要的基因结构不受破坏,同时重复序列也是新基因产生的物质基础,是驱动生物进化的重要因素之一[20].
低能离子注入对异常汉逊酵母菌基因组中的重复序列具有明显的致突变效应.微卫星DNA序列和小卫星DNA序列的总长度分别缺失了147bp和5 487bp;多种类型的SSR的重复基序不仅发生了单碱基和多碱基突变,其数量和拷贝数也发生了不同程度的变化,其中优势三碱基重复基序AAC增加了2条,新增的GTTA和TTCA为离子注入后的独具特征的SSR四碱基重复基序;离子注入使高拷贝区的SSR数目增加了1倍,并导致重复单元为47bp和56bp的小卫星DNA序列缺失,同时新增了重复单元为35bp、58bp和59bp的三种小卫星DNA序列.
对异常汉逊酵母菌基因组散在重复序列的分析发现,离子注入导致LTR缺失了5条、DNA转座子缺失了19个、RC缺失了1个,而LINE新增加了12条,新增总长度达1 568bp.
异常汉逊酵母菌基因组串联重复序列中两碱基重复序列数目最少,且仅有AT重复基序,在微卫星DNA和小卫星DNA中的含量均大于50%,这与全基因组DNA序列中AT含量较高有关.离子注入对AT重复基序没有任何影响,其分子机制有待进一步研究.
异常汉逊酵母菌作为汉逊酵母属中常见的一个种,具有一些酿酒酵母缺乏的酿造特性,是酿造产品香气成分的主要贡献者之一[21,22],有助于最终产品感官特性的提高[23],是高品质酿造食品研发的特色酵母菌株之一,本研究结果为其分子育种提供了理论基础,为其SSR分子标记的开发提供了丰富的DNA重复基序资源,也为后续开展其遗传多样性研究提供了基础数据.
现有的研究证明了低能离子注入是驱动微生物系统发育与进化的重要因素之一[1,2,24],本研究结果为进一步认识低能离子注入对酵母菌基因组重复序列的生物学效应及其基因表达与调控提供了素材,同时为低能离子注入介导的酵母菌基因组突变与进化提供了分子证据.
致谢
本研究由国家自然科学基金(11575149和31760016)和陕西科技大学博士科研项目(126021759)资助.北京诺禾致源生物信息科技股份有限公司微生物事业部协助进行了低能离子注入前后的异常汉逊酵母菌的全基因组de novo测序.
猜你喜欢
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
右江民族医学院学报(2022年2期)2022-05-19
南方医科大学学报(2022年3期)2022-04-13
河北医学(2021年10期)2021-10-27
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
中国临床医学影像杂志(2019年6期)2019-08-27
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
